INTRODUCTION
Urokinase-type plasminogen activator (uPA) and uPA receptor (uPAR) are central components of the plasminogen activation system. Plasminogen is converted to plasmin by uPA and the conversion is enhanced when uPA is bound to its cell surface receptor, uPAR.
Most physiological with tissue remodelling processes depend on activation of plasminogen to the proteolytically active enzyme plasmin, i.e., fibrinolysis, cell migration, wound healing, ovulation, trophoblast invasion, postlactational mammary gland involution, angiogenesis, activation of growth factors, cytokines and other proteolytic enzymes. In addition, tumour growth, invasion and metastasis also depend on the plasminogen activator (PA)-system [1-6]. However, uPAR not only functions as a protease receptor but also modulates cell adhesion, migration, chemotaxis, proliferation and differentiation through intracellular signalling [7-10]. Thus, through proteolytic and non- proteolytic functions, the PA-system, and uPA and uPAR in particular, are also involved in the inflammatory response and in innate as well as adaptive immunity.In 2000 it was demonstrated that high blood levels of soluble uPAR (suPAR) independently predicted mortality in antiretroviral-untreated HIV-1 infected patients [11]. Later studies confirmed the negative predictive value of high circulating suPAR levels in HIV-1 [12-15] and HIV-2 infection [13]. In addition to HIV-1 infection, the blood level of suPAR is increased in several diseases categorized by systemic inflammation and immune activation: Sepsis [16-19], Streptococcus pneumoniae bacteraemia [20,21], endotoxemia [17,22,23], systemic inflammatory response syndrome (SIRS) [24], certain CNS infections [25], pulmonary [13] and extra-pulmonary [26] tuberculosis, hepatitis B virus (HBV) infection [27], malaria [2830], uremia [31], rheumatoid arthritis and primary Sjogrens syndrome [32], multiple sclerosis and stroke [25] and several malignant diseases [33].
In many of these diseases, a high blood level of suPAR has negative prognostic value and may serve as a marker to monitor disease progression and treatment response: Streptococcus pneumoniae bacteraemia [20], SIRS [24], pulmonary tuberculosis [13], malaria [28,29], rheumatoid arthritis [32] and certain malignant diseases [34].In 2001, it was demonstrated that uPA exerts antiretroviral activity by inhibiting late steps in HIV replication cycle in various cell lines [35], a finding later confirmed by independent groups [36-38].
Given the strong prognostic value of suPAR and the molecular association between uPA and HIV-1 infection, the uPA/uPAR system and suPAR have been intensively investigated within the past 10 years.
In this chapter, the actors in the plasminogen activation system will briefly be introduced - with focus on uPA, uPAR and suPAR and their link to inflammation. The association between HIV-1 infection and uPA, uPAR and suPAR will be discussed in detail with reference to HIV-1 pathogenesis and highly active antiretroviral therapy (HAART). Finally, future perspectives will be discussed.
Biology of uPA, uPAR and suPAR uPA is secreted as an inactive single-chain proenzyme, pro-uPA [2]. Secreted pro-uPA binds to uPAR and is converted, faster than soluble pro-uPA, to active uPA through proteolytic cleavage by plasmin [4,5]. A positive feedback mechanism (referred to as reciprocal proenzyme activation) exists between pro-uPA and plasmin generation (and hence cell-associated proteolysis) since plasmin cleaves uPAR-bound pro-uPA which then converts neighbouring membrane-associated plasminogen to plasmin, tethering the proteolytic activity of plasmin to the membrane and hereby in close proximity to uPAR-bound pro-uPA [4,5]. Other serine proteases can also activate pro-uPA although their physiological significance remains unknown [1,2,4,5,39] (Fig. 1).
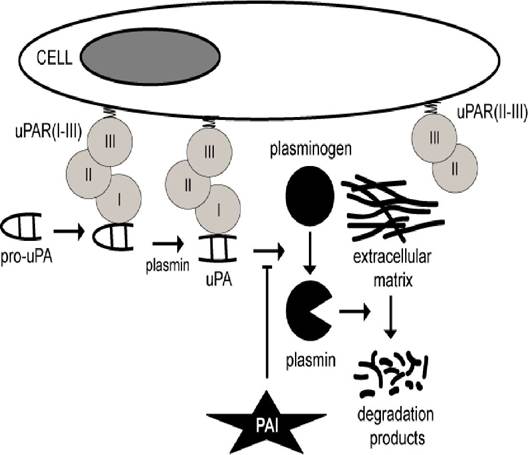
Figure 1: The urokinase PA-system.
The cascade is initiated by conversion of pro-uPA to active uPA by plasmin. uPA then converts plasminogen to plasmin, which degrades ECM components. The reaction is enhanced by binding of uPA to uPAR and by binding of plasminogen to its cellular receptor(s). The reaction is inhibited by plasminogen activator inhibitor (PAI)-1 and 2. The truncated receptor uPAR(II-III) can neither bind uPA nor pro-uPAuPA comprises two polypeptide chains that are held together by disulfide bridges. The carboxy-terminal B-chain contains the serine protease domain. The amino-terminal A-chain (ATF) contains the EGF-like domain and the kringle domain [2]. The serine protease domain is responsible for the catalytic activity whereas the EGF-like and kringle domains are responsible for the binding of pro-uPA/uPA to uPAR and for interaction with cell surfaces, respectively [3-5]. The enzymatic activity of uPA is about 250-fold higher than that of pro-uPA [5].Although uPA has restricted substrate specificity, it can activate latent forms of hepatocyte growth factor, macrophage-stimulating protein, pro-TGF-β, pro-IL-1, pro-IL-6 and MMPs [3,5,8]. Furthermore, uPA can convert the precursor chemokine HCC-1 (amino-acid residues 1-74) into a potent truncated form (amino-acid residues 9-74) which is a strong CCR5 agonist [40]. Finally, uPA catalyzes cleavage of PAI-1 and intradomain cleavage of uPAR with subsequent release of uPAR(I).
Due to the excessive background level of protease inhibitors, the most abundant circulating form of uPA is pro-uPA and complexes between uPA and PAI-1, as pro-uPA activation rapidly leads to complex formation between uPA and PAI-1 [41]. The activity of uPA is regulated by uPAR, plasmin, PAIs and by endocytosis.
uPAR is the cellular receptor for uPA [42]. It is a 55-60 kDa single-chain highly glycosylated protein consisting of three homologous domains (uP AR(I), uPAR(II) and uP AR(III)) [43,44]. uPAR contains no intracellular domain as the carboxy-terminal (uPAR(III)) is attached to the cell membrane by a GPI-anchor [45].
The amino-terminal uPAR(I) contains the binding site for uPA [44], but high affinity binding requires interaction with uPAR(II) and uPAR(III) as the intact receptor (and uPAR(I-II)) binds uPA with 102 to 103fold higher affinity than uPAR(I) alone [46]. uPAR(I) can also bind vitronectin, and again, the intact molecule is required for high affinity binding [47]. uPA and vitronectin can bind simultaneously to uP AR(I-III), and uPAR(I-III) binds more readily to vitronectin when it is pre-bound to either pro-uPA, uPA, ATF or the uPA/PAI-1 complex [47-51].
uPAR is expressed by most APCs (monocytes, macrophages, DCs) [52,53], neutrophils, eosinophils [52], activated T cells and NK cells [52,54-56], megakaryocytes [57], ECs [58-61], tubular epithelial cells [17], smooth muscle cells [60], keratinocytes, fibroblasts and many malignant cells [33,52]. It is absent on the cell surface of erythrocytes, platelets (though observed on platelets in [62]) and B cells [52,57] and only expressed in low levels by naive/resting T cells [54-56]. In the hematopoietic system, uPAR-expression follows a differentiation-dependent pattern in the myelomonocytic and megakaryocytic cell lineages but an activationdependent pattern in monocytes, neutrophils and T cells [52,63].
uPA, plasmin, chymotrypsin and other proteases can cleave uPAR(I-III) in the linker region between uPAR(I) and uP AR(II), with subsequent release of uP AR(I) [8,41,42,44,64]. uPA-catalyzed cleavage of membranebound uPAR(I-III) is favoured compared to cleavage of suPAR(I-III) whereas plasmin-catalyzed cleavage of membrane-bound and soluble uPAR(I-III) is comparable [65]. The intra-domain cleavage of uPAR(I-III) leaves the two-domain receptor uPAR(II-III) on the cell surface without ligand binding capacity [42,47,64]. Thus, uPAR is present on the surface of cells as three-domain uPAR(I-III) or as a cleaved receptor, uPAR(II- III) [64] (Fig. 2).
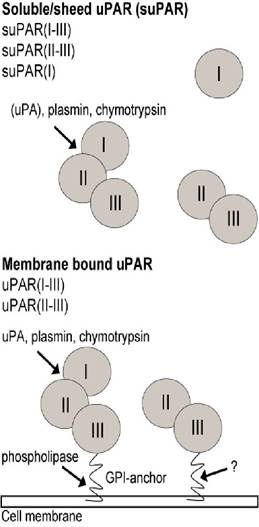
Figure 2: The urokinase receptor.
Domains and cleavage sites are shown. uPAR can be shed from the cell membrane by phospholipase C and D-catalyzed cleavage. uPA, plasmin, chymotrypsin and other proteases can cleave uPAR in the linker region between uPAR(I) and uPAR(II) with subsequent release of uPAR(I). uP AR(I-III) is shed from the cell membrane by neutrophil derived Cathepsin G, phospholipase C and D- catalyzed cleavage of the GPI-anchor [45,66] and by yet unknown mechanisms resulting in shedding of GPI- uPAR (suPAR(I-III) with an intact GPI-anchor) [42,66]. However, in contrast to previous findings, a recent study has found uPAR cleavage to be PLD independent [67], which might highlight the importance of neutrophil derived cathepsin D cleavage or yet undefined proteases responsible for uPAR cleavage. In addition, a soluble form of uPAR(I-III) resulting from alternative splicing of mRNA has also been reported [68]. It is not known whether and/or how uPAR(II-III) is shed from the cell surface, but suPAR is present in various human body fluids (blood, pleural-, pericardial-, peritoneal- and cystic-fluids, urine) and in cellculture supernatant as suPAR(I-III), suPAR(II-III) and suPAR(I) [16,69-72]. In addition, complexes between suPAR(I-III) and uPA, PAI, vitronectin, uPA/PAI, uPA/vitronectin and uPA/PAI/vitronectin are also present in body fluids [4,5,42].PAI-1 can inhibit both soluble uPA and uPAR-bound uPA through assembly of a PAI-1/uPAR/uPA complex. The complex is internalized through interactions between uPAR and LRP [4,42], which is followed by degradation of uPA and PAI-1 and recycling of uPAR [73].
Cleavage of cell-bound uPAR(I-III) also regulates the function of uPAR as uPAR(II-III) is unable to bind pro- uPA [65], uPA/PAI-1 complexes and vitronectin [47,65]. Consequently, uPAR(II-III) cannot promote plasminogen activation or be internalized and uPAR(II-III) is probably unable to mediate cell adhesion through integrin binding [74-76]. However, cleavage of cell-bound uPAR(I-III) exposes a potent chemotactic epitope that may function as a cell-bound or soluble chemokine (if shed from cell surface).