Physiology of Insulin Signaling and Adipose Tissue
Insulin acts on glucose and lipid metabolism, favoring the storage of energy after meals. At the clinical level, insulin resistance is defined as the inability or decreased ability of insulin to control glycemia, which means that a high level of insulin is required to maintain normoglycemia or that, when glycemia is increased, the levels of insulinemia are disproportionately elevated.
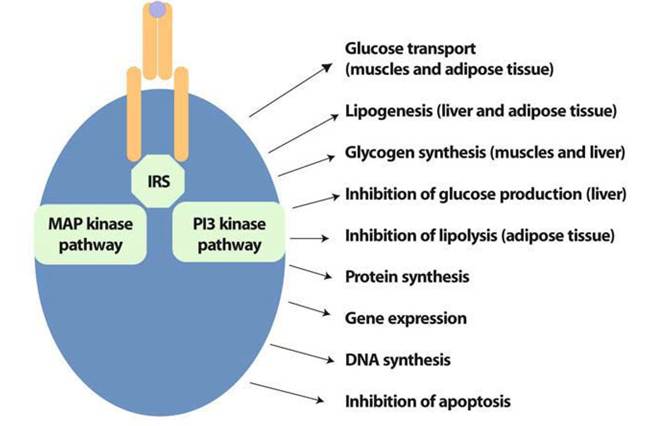
To evaluate insulin resistance, the most used index is HOMA (homeostasis model assessment), which is calculated from fasting glycemia levels (mmol/L) multiplied by fasting insulin levels (mU/ml) divided by 22.5. This index, or its derivatives (FIRI, QUICKI), is correlated with the evaluation of insulin sensitivity given by the gold standard test, i.e., the euglycemic hyperinsu- linemic clamp but, at the individual level, this correlation is rather low [8]. The measurement of fasting glycemia and insuline- mia mainly relies on the effect of insulin on the liver, where insulin inhibits hepatic glucose production by inhibiting glycogenolysis and gluconeogenesis. During the clamp test, or when a post-prandial measurement is performed, the presence of elevated levels of glycemia and insulin is mainly due to the defective entry and utilization of glucose at the muscle level.At the cellular level, the mechanisms whereby insulin transduces its signal have been studied for a long time, as have the mechanisms responsible for insulin resistance. In addition to the liver and muscles, adipose tissue is an important target tissue of insulin (Fig. 1). Insulin signals inside the cell by activating its receptor, which possesses a tyrosine kinase activity, and afterwards, insulin binding phosphorylates its intracellular domain, therefore allowing cytosol substrate proteins to associate with the receptor. The insulin receptor substrate (IRS) class of proteins transduces the signal towards different intracellular pathways: briefly, the metabolic signals are mainly transduced through the phosphatidylinositol 3 kinase (PI3K) pathway, while the mitogenic signals are mainly transmitted through the mitogen-activated protein (MAP) kinase pathway, which can use the IRS and SHC classes of proteins as receptor substrates.
At the level of the liver, insulin acts on glycogen synthesis and increases lipogenesis, in particular from glucose. It inhibits hepatic glucose production by impairing glycogenolysis and gluconeogenesis. In muscle cells, insulin recruits the glucoseINSULIN
The three main insulin target tissues are the liver, muscles and adipose tissue
Fig. 1 Insulin signal leads to the activation of major anabolic pathways. MAP kinase, mitogen activated kinase; PI3 kinase, phosphatidylinositol 3 kinase
transporters GLUT4 to the plasma membrane and activates glucose entry together with its storage as glycogen and its oxidation. In adipocytes, insulin plays a strong metabolic role by increasing TG storage through the activation of lipoprotein lipase and glucose entry and by decreasing TG hydrolysis through the inhibition of the hormone-sensitive lipase involved in lipolysis [9], a pathway stimulated by catecholamines in humans.
In 1963, Randle [10] described the glucose-fatty acid cycle and its role in physiology and pathology. He showed that free fatty acids (FFA) are used in preference to glucose by the heart and diaphragm muscles, leading to decreased glucose utilization. This concept has been revisited by Persegh- in et al. [11]. By performing both human and animal studies in vivo, they revealed that FFA entering the muscle cell are transformed into acyl-CoA derivatives and they activate the PKCθ isoform and IKKβ, which in turn activate IRS phosphorylation on serine residues. This serine phosphorylation has been shown by numerous groups to stop insulin signaling and to impede the activation of the downstream steps, including the recruitment of GLUT4 transporters from their intracellular location to the plasma membrane. In that situation, muscle and liver cells use FFA to produce the energy required for their metabolism.
In addition to its metabolic function, adipose tissue now appears to play an important endocrine role: it can release a number of hormones, proteins, and cytokines acting through endocrine or autocrine/paracrine mechanisms and collectively called adipokines (Fig.
2) [12-14].Leptin
Leptin is a 16-kDa protein mainly produced by subcutaneous adipose tissue, and it acts as an endocrine factor at the hypothalamic
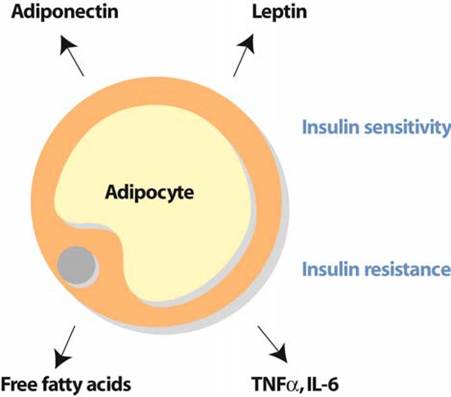
Fig. 2 Adipocytes secrete a range of adipocytokines which control insulin sensitivity
level to reduce food intake and to regulate energy production and utilization. In addition, leptin exerts pleiotropic actions and plays a role in peripheral energy and bone metabolism, reproduction, and immunity. Leptin receptors located on muscle cells have been shown to activate AMP kinase (AMPK) and thereby to favor FFA oxidation (see the following section).
Adiponectin
Adiponectin or adipoQ or ACRP30 is secreted at high levels by adipose tissue, and circulating levels are between 5 and 10 μg^l in human plasma. They are negatively correlated with BMI and with the content in visceral fat [15] and are decreased in diabetic and obese patients. It has been reported in numerous studies that the level of circulating adiponectin is inversely related to insulin resistance. Adiponectin is also decreased in patients with ischemic heart disease.
The mechanisms whereby adiponectin acts are now starting to be understood. Two receptors have been cloned, but the transduction pathway is still poorly understood. The activation of the two adiponectin receptors leads to the activation of the transcription factor PPARα and AMPK in the liver and muscles [16]. There has been a major interest in the role of this enzyme in cell metabolism. AMPK is activated by AMP, the level of which is increased when ATP has been hydrolyzed. AMPK now appears as an energy level-detecting enzyme, which plays a key role in redirecting the energy-synthesizing pathways of glucose and lipid metabolism. First, activated AMPK is able to indirectly activate the entry of acyl-CoA into the mitochondria to be degraded by the β-oxida- tion pathway in the liver and muscles and to be converted into energy.
In liver cells, AMPK is also able to inhibit the expression of the gluconeogenic enzymes required for hepatic glucose production. In muscle cells, AMPK can recruit glucose transporters GLUT4 to the membrane, even in the absence of insulin, allowing glucose entry and utilization [12]. In addition to its insulin-sensitizing effect, adiponectin can act at the level of the arterial wall. Atherosclerotic cellular changes include monocyte adhesion to endothelial cells due to the expression of adhesion molecules, uptake of oxidized LDL by macrophages through scavenger receptors, and proliferation of vascular smooth muscle cells in response to PDGF. Adiponectin has been found to inhibit tumor necrosis factor TNF-α production and TNF-α- induced adverse effects on the vascular wall, therefore inhibiting all these atherogenic processes [15, 17]. Thus, it probably has a potent anti-atherogenic role and could protect injured vessels against the development of atherogenic lesions. The expression of adiponectin has been found to be inhibited in vitro by TNF-α and interleukin (IL)-6.Tumor Necrosis Factor-α
TNF-α is synthesized as a 26-kDa plasma membrane-bound monomer. A secreted trimer is formed by proteolysis of the precursor, giving rise to a 17-kDa cytokine, which binds to two receptors, type I and II, and activates the classic inflammatory NFκB pathway [18]. In physiological conditions, adipose tissue production is very low.
However, its production has been found to be increased in pathological conditions, even if the reason for this increased production remains unclear. Animal studies have revealed that in cases of obesity and insulin resistance, the secretion of TNF-α was increased and could be involved in insulin resistance. In particular, TNF-α has been found to act at the local level on adipocytes and to inhibit insulin signal transduction through the inhibitory phosphorylation of the IRS1 protein together with a decreased expression of GLUT4 [12].
In human studies, the deleterious role of TNF-α has been questioned. Several studies reported that the circulating levels of TNF-α were related with obesity and insulin resistance. However, systemic inhibition of TNF-α in type 2 diabetic patients was not able to decrease insulin resistance. It could be proposed that TNF-α acts mainly at the local and not the systemic level and that it induces adipose tissue resistance, which results in increased lipolysis and FFA fluxes (Fig. 3). In addition, TNF-α acts on adipocytes by decreasing adiponectin and increasing IL-6 expression. These latter cytokines, which act at a distance from adipose tissue, probably play a major role in insulin sensitivity and indirectly trigger the insulin-resistant effect of TNF-α.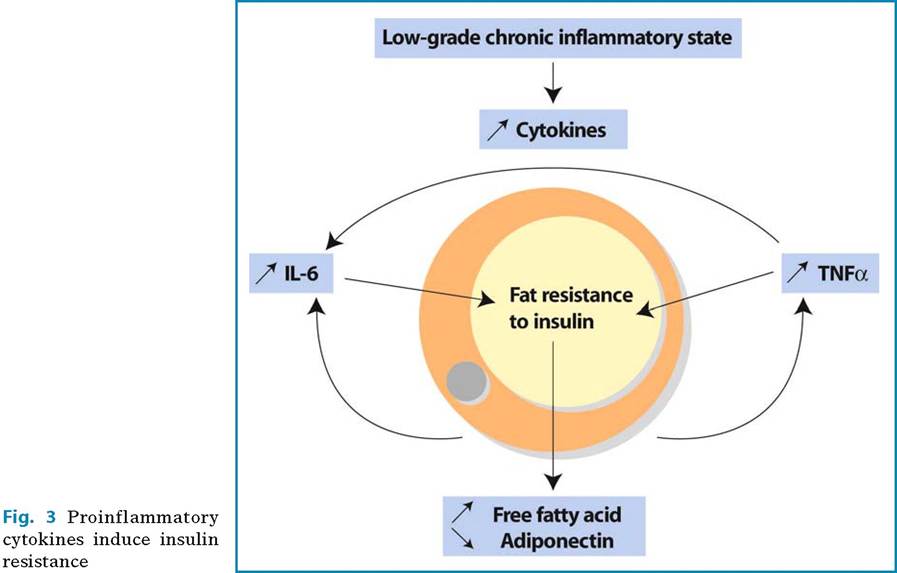
In addition, in patients with HIV-related metabolic syndrome, several studies consistently revealed increased levels of TNF-α, pointing to a major role for this cytokine in this condition (see the following section).
Interleukin-6
Interleukin-6 is a pro-inflammatory cytokine produced by several tissues and cells and, in particular, by adipose tissue: 10-30% of circulating Interleukin-6 could be produced by adipose tissue. Interleukin-6 may exert its effects at the central level on the hypothalamus. At the liver level, Interleukin-6 induces the production of acute-phase proteins and in particular of C-reactive protein (CRP). The levels of circulating Interleukin-6 were shown to be increased in obesity and were related to insulin resistance. Interleukin-6 also acts by paracrine/autocrine mechanisms on adipocytes and induces insulin resistance and altered differentiation in particular by inducing the expression of SOCS3 [19].
Plasminogen Activator Inhibitor-1
Plasminogen activator inhibitor-1 (PAI-1) is a protein involved in thrombosis, which compromises the clearance of fibrin. Its level is increased in obesity, type-2 diabetes, and metabolic syndrome. In addition, it is strongly associated with vascular disease including myocardial infarction and venous thrombosis [13].