uPA, uPAR AND suPAR IN HIV INFECTION
uPAR and HIV-1
Several studies have demonstrated that HIV influences uPAR-expression in immune cells. The first indication of a link between uPAR and HIV came in 1994 when Nykaer et al.
showed that CD3+ lymphocytes fromhealthy donors exhibited no significant uPAR expression whereas antiretroviral-untreated HIV-1 infected patients showed distinct uPAR expression in up to 80% of all T cells - and especially on activated HLA- DR+CD25+T cells [55].
This finding was later confirmed by in vitro studies reporting increased uPAR-expression (mRNA, protein) in T cells, monocytes and PBMC infected by HIV-1 in vitro [107,108]. In addition, it was shown that HIV-1- induced upregulation of uPAR was attributed to enhanced transcription rate of uPAR rather than stabilization of uPAR mRNA [107]. Also, autopsies from HIV infected patients with AIDS dementia complex or opportunistic CNS infections have revealed high uPAR-expression in HIV infected microglia cells, multinucleated giant cells and macrophages [109,110]. In contrast, granulocytes from HIV infected patients have reduced cell surface expression of uPAR that correlates positively with CD4-count [111]. Despite low uPAR-expression, granulocytes from HIV-1-infected patients displayed normal upregulation of uPAR in response to chemoattractants [111]. Also, children with AIDS have reduced levels of uPAR/uPA-expressing monocytes (but normal levels of uPAR/uPA-expressing lymphocytes in contrast to [55]) coinciding with increased uPA levels that increase with disease progression [112].
uPA and HIV-1
In addition to interaction between HIV-1 and uPAR, several studies have demonstrated interactions between HIV-1 and uPA.
In 1996, Handley and co-workers showed that uPA can bind to HIV-1 particles and cleave recombinant gp120 at the V3 loop of HIV-1, resulting in increased viral replication in vitro through enhanced HIV infectivity of macrophages and production of HIV-1 particles [113].
In contrast, several studies performed by two independent groups have demonstrated inhibitory effects of uPA on late steps of HIV replication [35-38].
In 2001, Wada and co-workers were trying to identify HIV antiviral factors in a CD8+ cell line supernatant (referred to as CAF), and found that one of the most potent antiviral protein among other candidates was ATF, the receptor-binding moiety of uPA [3 5]. The difference in uPA modulation of HIV infection is most likely due to the different cell models applied. While Handley used HIV virions and macrophages, Wada’s model system was based on the cocultivation of promonocytic U937 cells (bearing CD4 and CXCR4 on their cell surface) with a well-known chronically infected cell line (named U1) derived from acutely infected U937 cells. In the later system, HIV is generated from two integrated HIV DNA strands following activation with phorbol 12, myristate-13, acetate (PMA) or other extracellular stimuli including proinflammatory cytokines [114].
Alfano and co-workers [36] also used the U1 cell system and showed that uPA inhibits PMA induced HIV-1 expression through the trapping of virions in endosomes and not at the transcription level (Fig. 3).
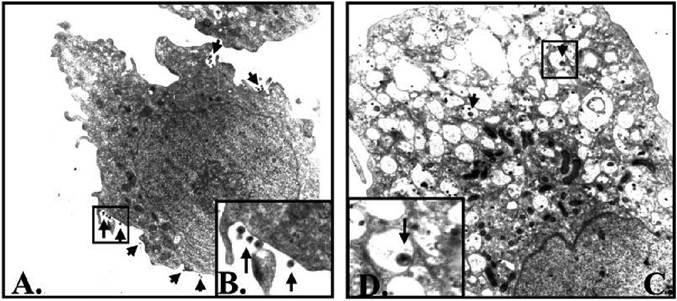
Figure 3. Accumulation of HIV-1 virions in intracytoplasmatic vacuoles in PMA-stimulated U1 cells in the presence of pro-uPA. U1 cells were stimulated for 48 h with (A) PMA (magnification ?11,000) or (B) PMA plus pro- uPA (10 nM) (magnification ?15,000). Pictures kindly provided by M. Alfano [36].
In 2007, Eila and coworker [37] found that the uPA mediated intracellular trapping of HIV virions in U1 cells was dependent of vitronectin, which binds the uPA/uPA receptor complex, resulting in cell adhesion to the culture disk surface. The authors suggest that macrophage cell adhesion per se curtails HIV replication.
In addition to binding uPA and vitronectin, uPAR interacts with Mac-1 (CD11b/CD18), a cell surface interaction that can be blocked using M25, a peptide homologous to a portion of CD 11b
Recently, Alfano and co-workers [38] showed that siRNA against uPAR as well as antibodies against Mac-1 abolished the anti-HIV effects of uPA in the U1 cell line. Also, addition of M25 inhibited HIV virion release in PMA-stimulated U1 cells. Altogether, either uPA/uPAR interaction, Mac-1 activation or prevention of its association with uPAR triggers a signalling pathway leading to the inefficient release of HIV-1 from monocytic cells.
In addition to a direct effect on HIV-1 replication, uPA can activate the precursor chemokine HCC-1 into a potent truncated form [40] that can block entry of HIV-1 strains that use CCR5 as a co-receptor [115].