ACQUIRED IMMUNITY
The acquired immune system is mediated by lymphocytes; it is most developed in vertebrates and best studied in mammals. Among mammals, common lymphoid progenitors (Fig. 2.1) are produced in the bone marrow and circulate through the blood stream as undifferentiated lymphocytes.
Some migrate to the thymus gland, where they undergo a rigorous but incompletely understood “maturation” process and emerge as T (thymus-derived)-lymphocytes. Others migrate to the bursa in birds, or to bone marrow or Peyer’s patches in mammals (Tizard 2004), where they undergo a comparable maturation process and emerge as mature B (bursa/ bone marrow-derived)-lymphocytes. Both Band T-lymphocytes have distinctive sites in the lymph nodes which they inhabit.Acquired immunity has two key components: humoral immunity and cell-mediated immunity (Bartl et al. 1994). Humoral immunity is based on B-lymphocytes that can respond to foreign proteins (antigens) by producing specific serum proteins (antibodies) against the antigens. When these specific antibodies chemically combine with a specific antigen, this combination often results in the host being protected from adverse effects of that antigen or the parasite on which it occurs. Antigens often are large and complex chemicals. The specific chemical moieties of an antigen that stimulate antibody production are called epitopes; each antigen can have multiple epitopes.
Antibodies are defined as immunoglobulins. In general, mammals carry three chemical forms of globulins: a-globulins, β-globulins, and g-globulins. Immunoglobulins all are g-globulins.
Cell-mediated immunity is based on T-lymphocytes and is particularly effective in destroying parasite-infected cells and cancer cells. The T-lymphocytes can act by directly destroying target cells or by releasing cytokines which, in turn, destroy the target cells or influence other host cells to enhance immune responses.
Activities of the cell-mediated immune system overlap with those of the humoral immune system.Unique Qualities of the Acquired
Immune System
There are some important and distinctive characteristics of the acquired immune system. One is a very high level of specificity. For example, a single B- or T-lymphocyte will respond to only one specific epitope. Because there are likely millions of different types of antigens a host can encounter, there must be millions of different types of antibody-producing B-lymphocytes in mammalian hosts, and a comparable number of T-cells. Consequently there is considerable heterogeneity among both B- and T-cells. Thus, although the immune response to antigens is highly specific, the immune system also has the potential to respond to a wide diversity of different antigens and their epitopes and, consequently, to their associated parasites and toxins.
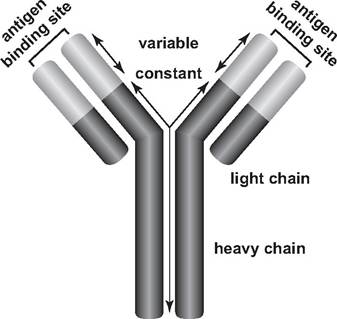
FIGURE 2.2 Diagram of an antibody (redrawn with permission of N. Anderson, University of Arizona).
A third important feature of the acquired immune system is an immunological memory. After the first exposure to an antigen, called a primary response, there is a 12- to 14-day lag before antibodies are detectable. For the hosts surviving this first exposure, a second, later exposure to that same antigen results in a shorter time span, often about 2 days, before specific antibodies to that antigen are detected. This is called a secondary response. Also, far greater numbers of those specific antibodies are produced in a short period in the secondary response. Thus, once a host has been exposed to a specific antigen, it develops an immunological memory for that antigen and responds much more quickly and aggressively to its presence. The features of specificity, heterogeneity, and memory are characteristic for both the humoral and cell-mediated systems of acquired immunity.
Humoral Immunity
In their basic form, antibodies consist of four polypeptides—two heavy chains and two light chains that are joined to form a “Y”-shaped molecule (Fig.
2.2). The tips of each “Y” are highly variable and consist of 110-130 amino acids, which vary to give the antibody its specificity for binding an antigen. This variable region includes the ends of both light and heavy chains.Each B-lymphocyte has about 200,000 to 500,000 antigen receptor sites on its surface for a specific antigen (Tizard 2004). These surface receptor sites resemble antibodies. As foreign materials invade a host, some make their way through the blood to the lymph nodes, where they come in contact with many leukocytes, including monocytes, macrophages, and lymphocytes. The B-lymphocyte receptors can bind to free, soluble antigen. However, to be fully activated, the B-cell also requires co-stimulation by a specialized T-lymphocyte called a helper T-cell that has been presented with that same antigen (Tizard 2004). When an antigen (or a specific epitope) attaches to a receptor site on the surface of the specific types of B-lymphocyte with which it can bind, that clone of B-lymphocytes is stimulated to multiply; each of the progeny also produces quantities of that specific antibody. Thus, humoral immunity is a result of clonal selection, the selection for the specific B-lymphocyte type that will produce antibodies against a particular epitope (antigen) among the many clones of B-lymphocytes in the host. B-lymphocytes actively producing antibody also are called plasma cells.
Of course, parasitic organisms are not composed of a single antigen; rather, their surfaces are a collection of many different macromolecular chemical groups. Thus, a host will recognize a number of these different chemical groups (epitopes) and will produce a polyclonal antibody response to each parasite.
Once antigen recognition has occurred, B -lymphocytes proliferate to produce clones of B-cells (plasma cells) bearing identical receptor sites, each actively engaged in antibody production. Memory cells are a subset of B-lymphocytes that may remain sequestered in the host for months to years after an earlier response to an antigen and provide a quick and enhanced response upon later exposure to the same antigen (Wakelin and Apanius 1997).
Hosts also can produce a number of different immunoglobulin classes having the same antigen specificity (Bush et al. 2001). Mammalian immunoglobulins (Ig) are divided into five major classes, IgM, IgG, IgA, IgD, and IgE, based on their chemical structure and immune function.
Immunoglobulin M first evolved in cartilaginous fishes and is found in amphibians, reptiles, birds, and mammals; it is often the first antibody to respond to an antigen (Tizard 2004). In contrast to most other antibodies, its shape is a pentamere with five “Y” structures fused at the base of the “Y”; IgM stimulates macrophages to engulf those objects it coats. This antibody also damages bacterial membranes; in combination with complement molecules, IgM produces holes in bacterial cell membranes and walls. However, the plasma cells soon convert from IgM to IgG production in mammals.
Immunoglobulin G, found only in mammals, is the most abundant antibody present in the mammalian circulatory system (Tizard 2004). It is very effective against blood- disseminated microorganisms (septicemia) and is also able to cross the placental barrier in mammals. The avian equivalent of this immunoglobulin is IgY (Tizard 2004).
Immunoglobulin A is reported in birds and mammals and provides a barrier against pathogenic organisms entering the host body; IgA-producing B-lymphocytes are found on the linings of intestines, as well as in the respiratory and genitourinary tracts, saliva, tears, and perspiration (Tizard 2004). This immunoglobulin occurs in high concentrations in the colostrum of mammals; colostrum is the first milk produced immediately after birth. Thus, nursing females transmit a rich supply of protective antibodies and a quick, but temporary, immunity to offspring. Immunoglobulin A is locally synthesized in mammary tissues, although many IgA-producing cells in the mammary gland are derived from precursors originating in the intestine; these intestinal cells are a source of antibodies against intestinal pathogens (Tizard 2004).
Immunoglobulin E is known to occur only in mammals; IgE, like IgA, is made by plasma cells located beneath body surfaces (Tizard 2004). While its function is not entirely clear, IgE may help initiate the inflammatory response. Immunoglobulin E binds to the surface of mast cells and basophils, leading to histamine release when a specific antigen combines with the IgE; its presence also is associated with allergic reactions such as hay fever and asthma in humans. It may function as a defense against helminths or arthropods (Tizard 2004).
Immunoglobulin D is known to occur in humans, some other mammals, certain fish, and some amphibians and reptiles, but not in birds (Tizard 2004, Davison et al. 2008). Immunoglobulin D may be the form of antibody attached directly to B-lymphocytes as the antigen receptor; it rarely is found free in the blood (Tizard 2004).
Cell-Mediated Immunity
The cell-mediated immune system is based on T-lymphocytes and is particularly effective in destroying parasite-infected cells and cancer cells. The T-lymphocytes can act by directly destroying target cells or by releasing cytokines which, in turn, destroy the target cells or influence other host cells in ways to enhance immune responses.
There are several types of T-lymphocytes, distinguished by the type of antigen receptors they carry, their accessory molecules, and their functions (Tizard 2004). These include two types of helper T-cells, regulatory T-cells, cytotoxic T-cells, and natural killer (NK) cells. Helper T-lymphocytes respond to antigens by secreting a variety of cytokines that influence the responses and activities of all other leukocyte types (Clough and Roth 1998). One type of helper T-cell collaborates with B-lymphocytes to enhance antibody formation; another induces a cell-mediated immune response and assists the direct cell-to-cell contacts associated with cell- mediated immunity (Clough and Roth 1998).
Regulatory T-cells can suppress some immune responses.
For example, they can suppress the proliferation of helper T-cells in response to antigens and prevent inappropriate T-cell activation in the absence of antigens (Tizard 2004).Cytotoxic T-lymphocytes respond to foreign antigens presented on cell surfaces by killing the cells that produced the antigens (Clough and Roth 1998). Cytotoxic T-lymphocytes differ
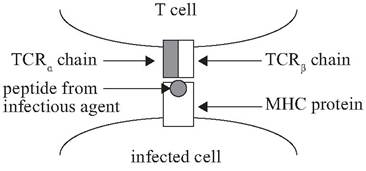
FIGURE 2.3 Model of an infected antigen-processing cell with a Major Histocompatibility Complex (MHC) protein presenting a foreign peptide to a T cell receptor (TCR) (From Bartl et al. Molecular evolution of the vertebrate immune system. Proceedings of the National Academy of Sciences 91: 10769-10770. Copyright 1994, National Academy of Sciences, USA).
from the natural killer (NK) cells discussed in the innate immune system in that the cytotoxic T-cells respond to specific antigens, whereas NK cells do not require previous exposure to a specific antigen to kill virus-infected or cancerous cells.
Cancerous host cells or host cells infected with intracellular microorganisms typically will have foreign peptides on their surface that attract cytotoxic T-lymphocytes (Clough and Roth 1998). Cytotoxic T-lymphocytes use specialized T-cell receptors on their surfaces to attach to peptides from infectious agents that have been bound to proteins on the surface of infected cells or tumor cells (Fig. 2.3) (Bartl et al. 1994). The T-cells can kill these infected or tumor cells by binding molecules on the surface of these cells; this binding often initiates dissolution of the nucleus or disrupts the fluid equilibrium of the cell (Clough and Roth 1998). In the process, the T-lymphocytes also may cooperate with phagocytes or B-cells in destroying these target cells. Thus, T-lymphocytes, B-lymphocytes, and the innate immune system all can work in a cooperative fashion.
Some T-lymphocyte cytokines may regulate the activity of other T-lymphocytes, as well as B-lymphocytes. Depending on the specific cytokines released, T-lymphocytes can inhibit the response of helper T-lymphocytes involved in the antibody response and inhibit proliferation of the cytotoxic T-lymphocytes (Bush et al. 2001). They also may induce and influence the inflammatory responses by regulating the production, migration, and functional activation of granulocytes and monocytes (Wakelin and Apanius 1997). Different parasites selectively stimulate different helper T-lymphocytes subsets; each T-lymphocyte subset secretes a characteristic set of cytokines, referred to as a T-lymphocyte cytokine profile (Bush et al. 2001).
During chemical breakdown and processing of antigens, the resulting peptides become bound to major histocompatibility complex (MHC) receptors on the surface of the antigenprocessing cells (Tizard 2004). The MHC proteins occur on the membranes of host cells and are highly pleomorphic between individuals in a species; they are required in the presentation of processed peptide antigens to T-lymphocytes (Clough and Roth 1998). T-lymphocytes use their T-cell receptors to respond to antigen in the form of processed peptides bound to the cell surface proteins encoded in the MHC. The importance of MHC molecules in immune responses against pathogens led to the proposal that the high diversity of MHC molecules evolved as a result of parasite interactions (Snell 1968, Doherty and Zinkernagel 1975). Further, there is evidence that parasites with a long history of co-evolution with their hosts have had the greatest impact on this MCH polymorphism (Klein and O'Huigin 1997).
For T-lymphocytes, two types of receptor molecules are required for recognition of antigen. First, the MHC molecule on the antigen-processing cell (e.g., dendritic cell) binds the processed antigenic peptide; second, the T-cell receptor binds the MHC molecule-antigenic peptide complex (Tizard 2004). The MHC receptor molecules appear on the surface of dendritic or other antigen-presenting cells (Fig. 2.3). It is only when antigenic fragments are presented by an MHC receptor in this way that they can be recognized by specific T-lymphocyte receptors or initiate specific B- or T-lymphocyte immune responses (Doherty and Zinkernagel 1975, Wakelin and Apanius 1997). This dependence on correct MHC-mediated presentation is called MHC restriction and constitutes a potential bottleneck for recognition of parasite antigens (Zinkernagel and Doherty 1979, 1997). While processing of complex antigens such as those derived from parasites may produce many epitopes, most subsequent immune response is directed against a relatively small number of “immunodominant epitopes” (Wakelin and Apanius 1997).
Both B - and T-cells have similar types of specificity, heterogeneity, and memory. Thus, each T-lymphocyte also responds to a single epitope and, following combination with a specific antigen, there is a clonal proliferation of these cells. Cellular immunity involves the presentation of antigenic epitopes in combination with specific macrophage surface molecules (the major histocompatibility proteins) to the T-lymphocytes.
Basis of lymphocyte heterogeneity
A mammalian acquired immune system requires antigen receptors (immunoglobulins and T-cell receptors), antigen-presenting molecules (MHC), and gene-rearranging proteins (Fig. 2.3) (Bartl et al. 1994). It is estimated that mammals can produce up to 1015 different antigen receptors to be expressed on B- and T-lymphocytes, but that in order to produce this enormous diversity they use fewer than 500 genes (Tizard 2004). Initially, stem-cell lymphocyte precursors all are alike in genetic composition within an individual host. In mammals these precursor cells undergo a maturation process in the bone marrow, Peyer's patches, or thymus whereby their genetic compositions are altered so that they encode unique receptor proteins and each becomes able to recognize and respond to a unique epitope.
The key to generating receptor diversity of gene-rearranging proteins lies in the fact that multiple genes are required to code for each receptor peptide chain. Several genes code for each variable region of the B -cell or T-cell receptors, whereas one gene codes for the constant region. As a result, a single constant-region gene can be combined with any one of a large number of different variable-region genes to make a complete receptor peptide chain. Thus, instead of having to store information about all possible receptor chains, it is only necessary to store the information (genes) for all the variable domains and to match these, when required, with the appropriate constant-region gene to produce a complete range of receptors. Following this, light and heavy chains may be paired in different combinations, a process called combinatorial association (Tizard 2004).
The development of lymphocyte heterogeneity can be seen as analogous to shuffling a deck of several thousand cards and then randomly distributing a limited number of cards (genes) among many players (lymphocytes). There are a huge number of possible combinations of genes, with little likelihood of identical combinations. And in the case of immunity, there are three separate gene segments from which one is selecting and distributing genes (Bartl et al. 1994).
Basis of immunological memory
Immunological memory is based on differential responses by B- or T-lymphocytes of a clone encountering a specific antigen. Most B- or T-lymphocytes that respond to a specific antigen multiply, produce antibodies (B-cells), or attack target cells (T-cells), and die. However, a few cells in each of these clones remain quiescent; these long-lived “memory cells” remain ready to respond to a subsequent exposure to the same antigen.
Length of immunological memory is greatly influenced by the nature of the antigen. Although some antigens, such as mumps and smallpox in humans, may elicit a lifelong immunity, many others (e.g., rabies) have an effective immunity of a few years. In contrast, botulism toxin results in a short memory or none whatsoever, based on recovered birds that were banded and released, and succumbed to the disease the following year. It is possible that a sublethal dose of toxin may have too little antigenic mass to stimulate an antibody response.
More on the topic ACQUIRED IMMUNITY:
- IMMUNITY AMONG DIFFERENT ANIMAL GROUPS
- Acquired Inhibitors of Coagulation Factors
- Community-Acquired Pneumonia
- INNATE IMMUNITY
- PATHOGENESIS, PATHOLOGY AND IMMUNITY
- ACQUIRED AMPUTATIONS
- Commonly Acquired Lower-Limb Amputations in Children
- Acquired Coagulation Disorders
- Noninfantile Acquired Botulism
- TWO Introduction to Immunity