Avian respiration
There are several differences between the avian and mammalian respiratory systems. Unlike mammals, bird lungs are inelastic and therefore do not change volume during respiration.
In birds, the relative volume of the trachea is much larger, and therefore the bird must compensate for this large dead space. Rather than having alveoli, gas exchange in birds occurs in air capillaries. Furthermore, since birds lack a diaphragm, the movement of the ribs and the sternum are important in changing the abdominal pressure in order to move air in and out of the air sacs, another structure unique to birds.Anatomy of the avian respiratory system
The avian respiratory system has a unique structure compared to other vertebrates (Fig. 14.12). The lungs do not expand; instead, air sacs act as bellows to move air in and out of the lungs. Gas exchange occurs in the lungs, but not the air sacs. Also, birds have no diaphragm, and the thoracic cavity is at atmospheric pressure instead of negative pressure as in mammals. Although birds have a larynx, sound is instead generated in the syrinx, which is composed of cartilage and vibrating soft tissue.
The avian lung is a rigid structure located in the thoracoabdominal cavity. The trachea extends from the larynx to the syrinx, and it is composed of complete tracheal cartilages. Tracheal volume in birds is about 4.5 times larger than in mammals since avian trachea are longer and wider, creating a larger dead space. Birds compensate by having a larger tidal volume and lower respiratory frequency.
Short primary bronchi extend from the syrinx to the lungs. The Intrapulmonary primary bronchus travels through the lungs to the abdominal air sac at the caudal border of the lungs. Two groups of secondary bronchi exist. The cranial group includes 4-5 medio- ventral secondary bronchi originating from medio- ventral Intrapulmonary bronchi; the caudal group includes 6-10 mediodorsal secondary bronchi.
There is also a third group, which varies among species, consisting of Iateroventral secondary bronchi. The primary and secondary bronchi do not participate in gas exchange.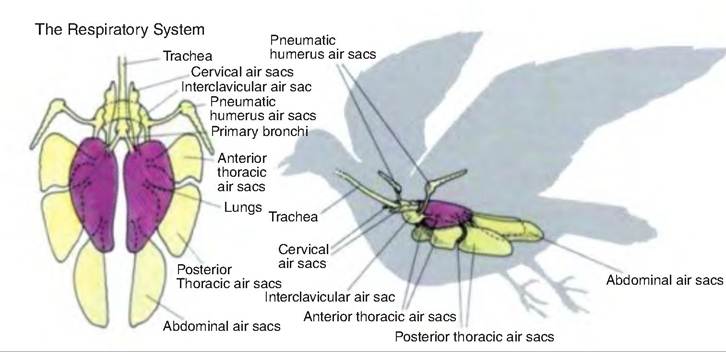
Fig. 14.12. Respiratory system in birds. (Reprinted from http://www.paulnoll.com/Oregon/Birds/Avian-Respiratory.html.)
The parabronchi, also called tertiary bronchi, originate from the secondary bronchi and are the site of gas exchange. There are two types of parabronchi. The medioventral and mediodorsal secondary bronchi, also called paleopulmonic parabronchi, appear as parallel tubes extending from the secondary bronchi. Gas flow through the paleopulmonic parabronchi is unidirectional. There is also an irregular branching system of neopulmonic parabronchi through which gas flow is bidirectional. The lumen of the parabronchi is lined with smooth muscle surrounding the entrances to the atria that branch off and lead to infundibula, which give rise to air capillaries. The air capillaries intermesh with pulmonary blood capillaries, the site of gas exchange.
Air sacs
Air sacs are poorly vascularized, thin membranous structures connected to either the primary or secondary bronchi via ostia. Comprising most of the volume of the respiratory system, they do not participate in gas exchange, but function to move air through the lungs so that gas exchange can occur in the parabronchi. Air sacs can also extend into selected bones.
Birds possess nine air sacs; four are paired and one is unpaired (Fig. 14.12). The cranial group of air sacs consists of paired cervical and cranial thoracic air sacs and the unpaired clavicular air sac. The caudal group includes the paired caudal thoracic and paired abdominal air sacs.
Avian ventilation and gas exchange
Since birds do not have a diaphragm, they have a single thoracoabdominal cavity, which functionally behaves as a single compartment. During inspiration, the sternum moves both Cranially and ventrally as the coracoids and furcula (wishbone) simultaneously rotate forward at the shoulders (Fig.
14.13). The sternal ribs also move cranially, thus laterally expanding the sternal ribs and thoracoabdominal cavity. Both inspiration and expiration are active processes involving various muscles (Table 14.2). Therefore, in birds, the volume of the respiratory system is half of end- inspiratory and end-expiratory volumes when relaxed. This is in contrast to mammals, where during relaxation only the residual volume remains.During inspiration, approximately half the tidal volume first enters the caudal air sacs and half enters the cranial air sacs. As the thoracic cavity expands during inspiration, the volume, and therefore pressure, inside the air sacs decreases. This causes air to move caudally through the Intrapulmonary bronchus. About half of this air goes through the neopulmonic lung continuing directly to the caudal thoracic and abdominal air sacs. The other half goes into the mediodorsal secondary bronchi and into the paleopulmonic parabronchi, and then into the cranial group of air sacs (Fig. 14.14). During expiration, contraction of the expiratory muscles decreases the volume of the thoracoabdominal cavity causing air to flow out of the caudal thoracic and abdominal air sacs (Fig. 14.14). This gas
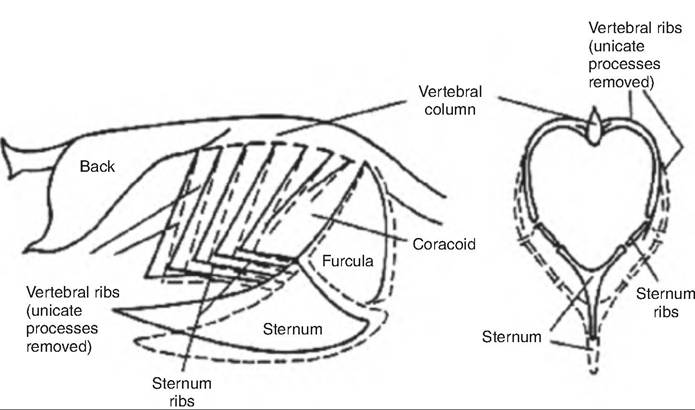
Fig. 14.13. Changes in the thoracic skeleton during respiration in birds. Solid lines show thoracic position at the end of expiration; dashed lines show position at the end of inspiration (Reprinted from Whittow, 2000. Used by permission of the publisher.)
Table 14.2. Muscles involved in respiration in birds.
| Inspiration | Expiration |
| M. scalenus | Mm intercostals externi of fifth and sixth spaces |
| Mm intercostals externi | Mm intercostals interni of |
| (except in fifth and sixth spaces) | third to sixth spaces |
| M. intercostalis interni in second space | M. Costasernalis pars minor |
| M. Costostrernalis pars major | M. obliquus externus abdominis |
| Mm. Ievatores costarum | M. obliquus internus abdominis |
| M. serratus profundus | M. transversus abdominis M. rectus abdominis M. serratus superficialis, pars cranialis and caudalis M. Costoseptalis |
passes through the neopulmonic lungs to the paleo- pulmonic lungs. Simultaneously, air leaves the cranial air sacs through the medioventral secondary bronchi flowing into the primary bronchus and the trachea to exit the body. Therefore, during quiet breathing, all air moves through the paleopulmonic parabronchi, and air passes through the paleopulmonic parabronchi in a caudal-to-cranial direction while traveling through the neopulmonary parabronchi bidirectionally.
Gas exchange occurs between the air capillaries and blood capillaries. As shown in Figure 14.15, the flow of gas within the air capillaries is in the opposite direction as the flow of blood in the blood capillaries. Thus, this is a countercurrent system. As air moves through the parabronchus, CO2 is added to the blood while O2 is removed from the blood. The interaction between the respiratory system and the cardiovascular system is shown in Figure 14.16.
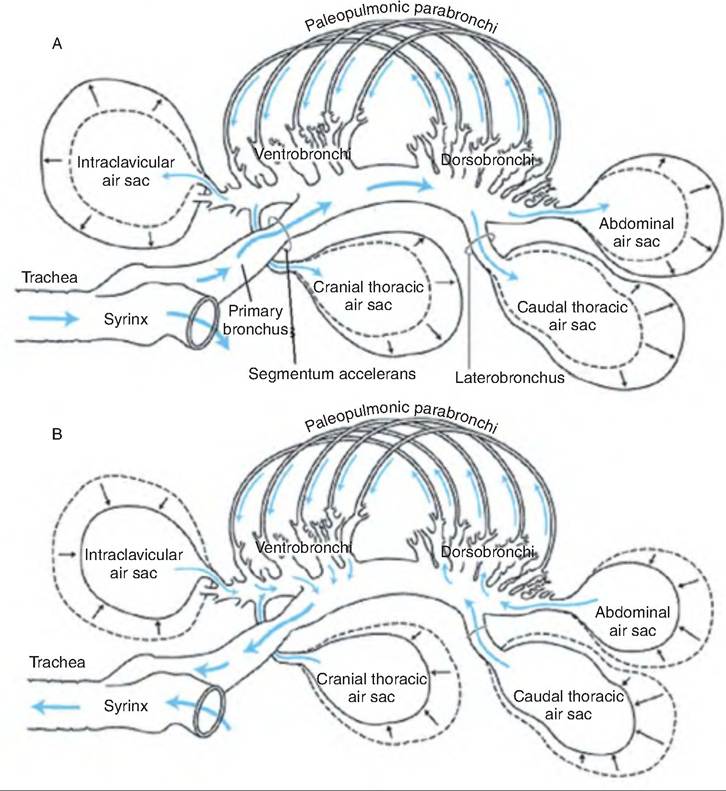
Fig. 14.14. Pathway of gas flow through the avian pulmonary system during inspiration and expiration. (A) Inspiration through the right paleopulmonic parabronchi; the Iteopulmonic parabronchi have been omitted for clarity. (B) Expiration. (Used with permission from John W. Ludders and Michael Simmons, Cornell University, http://people.eku.edu/ritchisong/birdrespiration.html).
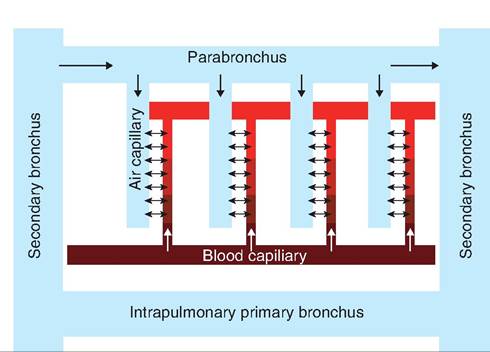
Fig. 14.15. Cross-current gas exchange in the avian lung. The parabronchi lie parallel to the intrapulmonary primary bronchus.
Air capillaries branch perpendicular from the parabronchi. Gas exchange occurs between the air capillaries and blood capillaries, each of which flows in an opposite direction. (Modified from Sturkie, 1986.)
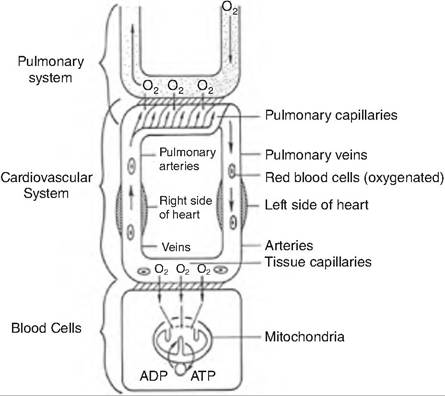
Fig. 14.16. Cooperation between respiratory and circulatory systems. Oxygen diffuses from the air capillaries to the pulmonary capillaries. It is then carried through the cardiovascular system and delivered to the tissues. (Reprinted from Sturkie, 1986. Used by permission of the publisher.)
Prehatching respiration in birds
Bird eggs contain all the nutrients necessary for embryonic development except oxygen. Oxygen passively diffuses into the egg through the pores found in the eggshell while carbon dioxide diffuses out of the same pores. A normal chicken egg contains approximately 10,000 pores. The number of these pores is important because the shell must allow adequate entry of oxygen to meet the embryo's needs while simultaneously preventing too much moisture loss, which would result in dehydration, or too little carbon dioxide loss, thus disrupting acid-base balance.
During the first 18 days of incubation, the prenatal period, the capillaries located in the chorioallantoic membrane, analogous to the mammalian placenta, function in gas exchange across the shell and its associated shell membranes. Such gas exchange is ample to meet the embryonic needs through 18 days of incubation. After this time, the embryo prepares for hatching. Simple diffusion across the shell is insufficient to meet the oxygen needs of the embryo.
On day 19 of incubation, the chick's beak penetrates the internal shell membrane, thus poking into the air cell located at the blunt end of the egg. This process, called internal pipping, allows the chick to begin pulmonary respiration using air located in the air cell. This time during which the embryo is breathing from the air cell is called the paranatal period. About 6 hours after internal pipping, the chick's beak penetrates the eggshell, which is called external pipping. The chick is now able to receive sufficient oxygen to complete the hatching process.