CHAPTER 38 ASPERGILLOSIS
DESIREE S. JANSSON
Department of Animal Health and Antimicrobial Strategies, National Veterinary Institute & Department of Biomedical Sciences and Veterinary Public Health, Swedish University of Agricultural Sciences, Uppsala, Sweden
Aspergillus is a large ascomycetous genus of saprobic filamentous fungi that is of great importance in human and veterinary medicine, agriculture, biotechnology, the food industry and genetic research.
Invasive and non- invasive infections of vertebrates by Aspergillus spp. are collectively named aspergillosis. Aspergillosis is the most important fungal respiratory infection in birds and a major cause of mortality in free-living, captive and domestic birds worldwide. Sporadic cases also occur in wild and domestic mammals. Aspergillosis is also named fungal, or mycotic, pneumonia, pneumonomycosis, bronchomycosis and brooder pneumonia; and it is caused by infection with A. fumigatus, or more rarely A. flavus, A. niger, A. nidulans, A. terreus, and others.AETIOLOGY
Aspergillus spp. are fungi that recycle carbon and nitrogen from organic sources, particularly decaying vegetation in soil, compost heaps, litter and crops. They are cosmopolitan multicellular organisms consisting of elongated, septate, 3—6 μm thick, dichotomously branching cells (hyphae) that grow by apical extension to form a network (mycelium) on and within substrates. When exposed to air some hyphae differentiate into T- or L-shaped foot cells, which produce specialized structures called conidiophores (spore-bearing branches) in a perpendicular orientation to the axis of the foot cell. At the apex of the conidiophore, a vesicle is formed on which chains of mitotic (asexual) single-celled smooth- or rough-walled spores called conidia or conidiospores are formed. Conidia are released into air, and when they settle on a suitable substrate and water is available, they germinate and grow into a mycelium.
Sexual reproduction by ascospores is known in a subset of Aspergillus spp., and was only very recently identified in A. fumigatusNThe genus Aspergillus contains approximately 250 species. Fungal nomenclature is governed by the International Code on Botanical Nomenclature, based on phenotypic features, most importantly the morphological appearance of sexual and asexual reproductive structures. According to nomenclatural rules, the teleomorph (sexual) state takes precedence over the anamorph state (asexual form). Hence, aspergilli that lack a known teleomorph state are classified as fungi imperfecti, i.e. within the artificial form-phylum Deuteromycota, genus Aspergillus. However, when sexual reproduction is recognized the species are classified within the phylum Ascomycota. A single species may thus be known under two different names that are used interchangeably, e.g. Neosartorya fumi- gata and A. fumigatus, but by strict interpretation of the code, N. fumigata is the correct name. A polyphasic approach, which considers data on morphology, physiology, metabolism and multilocus sequence information, is gaining in importance. By this system, all aspergilli are considered to belong to phylum Ascomycota, genus Aspergillus, and the genus is subdivided into eight subgenera, numerous sections and species.
Infectious Diseases of Wild Mammals and Birds in Europe, First Edition. Edited by Dolores Gavier-Widen, J. Paul Duff, and Anna Meredith. © 2012 Blackwell Publishing Ltd. Published 2012 by Blackwell Publishing Ltd.
Complete genome sequences are available for several Aspergillus spp., including A. fumigatus. The genome size is between 29 and 37 Mb, and comparative genomics has shown extensive interspecies variation, but low levels of genetic diversity between strains of A. fumigatus, speculatively because of its largely clonal population structure1-2). The A. fumigatus genome is divided into eight chromosomes, each between 1.8-4.9 Mb in size, and there are 9,632 protein- coding genes, most ofwhich are of unknown function)3).
EPIDEMIOLOGY
Aspergillus fumigatus is the predominant agent in both mammalian and avian aspergillosis, but approximately 40 species are known opportunistic pathogens of vertebrates. Based on molecular characterization of a large number of environmental and clinical isolates of human origin, it has been suggested that, given a susceptible host, all A. fumiga- tus strains should be considered capable opportunistic agents(4). However, interstrain variation in virulence has been observed. Moreover, polyclonal infection by A. fumi- gatus has been reported from both humans and birds in recent years, which may have potentially important repercussions on diagnostics and therapy)5). Aspergillus spp. are aetiologic agents of a range of diseases in humans and in domestic animals, such as bovine mycotic abortion, sino- nasal infections in dogs, guttural pouch mycosis in horses and respiratory infections in poultry and other captive birds. Aspergillosis in poultry is a particularly important and costly disease, which is associated with reduced welfare and financial losses from mortality and carcass condemnation, especially in the turkey industry.
Aspergillosis is a rarely reported condition in free-living and captive wild mammals including cervids such as red deer ( Cervus elaphus) and roe deer ( Capreolus capreolus))6’7) cetaceans (dolphins and whales))8-10), and hare (Lepus spp.)(11). Malnutrition and immunocompromisation from anthropogenic toxins, heavy metals or viral infections have been suggested as possible predisposing factors to pulmonary aspergillosis or mycotic encephalitis caused by A. fumigatus in cetaceans. Aspergillosis is a potentially underdiagnosed infection in marine mammals and much remains to be learned about its epidemiology.
Information on prevalence of aspergillosis among wild birds is fragmentary and shows broad variations between studies and avian species. All avian species should probably be considered susceptible, but there seems to be a species predilection.
Most reported cases among free-living birds involve waterfowl (particularly Anatidae), gulls (Laridae) and birds of prey (Accipitridae and Falconidae), which may reflect true species differences, a higher risk of exposure or a higher likelihood of detection. Among captive wild birds disease has been reported in a much wider range of families and species. Aspergillosis is a non- contagious disease. The main route of entry of conidia in both mammals and birds is the airways. However, in birds aspergilli may penetrate egg shells during incubation, and growth in ovo is a well-known cause of embryonic and hatchling mortality, i.e. brooder pneumonia. Age dependency has been reported, and most cases of aspergillosis in poultry involve young birds, following massive exposure to conidia during incubation or post- hatch from aerosolized conidia from mouldy litter, straw or feed. More rarely, conidia or hyphal fragments may be introduced by puncture wounds or during surgery. Aerosolized conidia may induce keratitis. Data on the infective dose of A. fumigatus in poultry and wild birds are contradictory and may reflect differences between avian species, age, immunity, strain virulence and experimental conditions. After transcutaneous air sac inoculation in turkey poults, inflammatory lesions are apparent within 24 hours and progress rapidly(12).There are two main forms of aspergillosis occurring among free-living birds: an acute epizootic form, and sporadic disease in individual birds. Acute aspergillosis is sometimes manifested as sudden mass-mortality events in a restricted area. Such outbreaks usually involve anseriform birds, and are infrequently reported mainly in the North American literature, but could most probably occur elsewhere. The timing and location of these outbreaks are highly suggestive of a common source of conidia and exposure within a limited time frame. Outbreaks are often, but not invariably, associated with adverse weather conditions and availability of mouldy agricultural waste crops.
Otherwise healthy birds appear to be overwhelmed by massive conidial exposure and develop acute epizootic aspergillosis, but it is unclear if immunocompromisation is a necessary prerequisite. Outbreaks of acute aspergillosis are also seen in poultry and on game farms. The other form of aspergillosis in free- living birds is much more common and involves incidental and usually subacute to chronic cases. Affected birds often show signs of trauma, inanition or concomitant disease such as parasite infections1-13), mycobacteriosis(14), lead toxicity(15), carbofuran toxicity(16) or oiling(17). There is circumstantial evidence that aspergillosis may be a significant cause of mortality in certain free-living avian populations, such as in immature white storks (Ciconia ciconia) in Germany(18). Occasional reports of aspergillosis involve near-threatened, conservation-dependent or endangered species. It cannot be ruled out that this disease may have a negative impact on such free-living populations, especially if combined with other infectious diseases or toxins. Also, because aspergillosis is a particularly common sequel to captivity in zoological exhibits, during research projects, in relocation programmes or at rehabilitation centres, it is a potentially important cause of failure of conservation and translocation programmes of avian species.PATHOGENESIS, PATHOLOGY
AND IMMUNITY
The past decade has seen remarkable progress in the understanding of mammalian host-pathogen interactions and virulence determinants of Aspergillus spp. The current view is that aspergilli make use of their saprobic lifestyle, thermotolerance and competitiveness, and that these help ensure survival both in their complex environmental niche and during infection. Conidia of A. fumigatus are widely dispersed in air because of prolific asexual reproduction, conidial longevity and small size, which allows them to remain airborne.
Conidial numbers may reach particularly high levels in poultry barns and stables, at composting plants and in damp buildings. Because of their small size, conidia will readily penetrate airways, including alveoli in mammals and air sacs in birds. It is generally assumed that conidia are continuously inhaled and rendered innocuous by innate immunity mechanisms. Many conidia get expelled by mucociliary action, whereas others are internalized by respiratory epithelial cells or adhere to exposed extracellular matrix components. Alveolar macrophages recognize and kill conidia, and they trigger a polymorphonuclear immune response. Based on studies of the pathogenesis in mammalian animal models it is known that neutrophils extravasate and migrate towards the site of infection in response to chemokines that are generated during complement activation, and they use a range of pathogen recognition receptors to recognize and eliminate hyphae by reactive oxygen and nitrogen species and enzymatic granule components, and by phagocytosis. Further, dendritic cells seem to trigger an adaptive immune response1-19). In mammals, neutropenia, defective phagocyte function, and corticosteroids that impair macrophage function and epithelial integrity, are recognized risk factors for aspergillosis.Aspergilli make use of both active and passive mechanisms to circumvent innate immunity, e.g. cilio-inhibitory, cytotoxic and immunomodulatory mechanisms, and the majority of hitherto identified putative virulence determinants in aspergilli do not display classical virulence attributes. Candidate pathogenicity determinants include secondary metabolites such as catalase, proteases, elastase and the immunomodulatory substance gliotoxin, genes responsible for thermotolerant growth, a variety of cell wall components, enzymes involved in the pyrimidine biosynthesis, histidine kinases, a Ras-related protein, cAMP signalling pathways and calcineurin, nutrient sensing system components, which ensure versatility of nutrient availability, and substances that are involved in resistance to phagocytosis and scavenging of reactive oxygen species. Recent reviews expand upon virulence determinants and mammalian host-pathogen interactions1-20-22).
I mmunosuppression or transgenic models of immune deficiency combined with high infective doses are necessary prerequisites in mammalian experimental models of aspergillosis. In contrast to mammalian experimental models, immune modulation is not a necessary prerequisite in avian challenge models. Aspergillosis is readily induced by inhalation of aerosolized conidia, by intravenous and intratracheal routes, and by transcutaneous inoculation into air sacs in otherwise healthy birds. Knowledge on predisposing factors in avian aspergillosis relies mostly on anecdotal evidence. It is tempting to speculate that the apparent higher susceptibility of birds when compared with mammals may be attributed to anatomic and physiologic characteristics of the avian respiratory system, immune mechanisms and/or a higher body temperature that allows quicker fungal growth. Birds have small, nonexpanding lungs, and air sacs that act like bellows and hold most of the respiratory volume. The avian air sacs usually constitute a primary nidus of infection because the air (and conidia) reaches the caudal air sacs before it passes through those parts of the lungs in which the gas exchange takes place. The mucociliary system is unevenly distributed in the air sacs, and macrophage numbers in distal airways are low, although there is presently no scientific proof that this explains the apparent higher susceptibility of birds. In addition, avian heterophils primarily depend on nonoxidative mechanisms1-23). There is generally little information available on immunity mechanisms in avian aspergillosis.
Acutely affected birds are often in normal body condition, but as the disease progresses, birds lose condition and may become emaciated. Lesions may be confined to air sacs and lungs, but trachea, syrinx and bronchi may also be primary targets. Virtually any organ and tissue can become secondarily involved from direct extension of contiguous respiratory lesions, and/or by haematogenous dissemination of fungal elements contained within macrophages. Birds with acute aspergillosis present with pulmonary congestion, oedema, haemorrhage, consolidation and miliary 1—10 mm diameter, coalescing, round to irregular, caseous, parenchymatous and pleural nodules (Figure 38.1), sometimes accompanied by white luminal domeshaped nodules or plaques in air sacs. The more common subacute to chronic presentation is characterized by diffusely thickened, opaque, and prominently vascularized air sac membranes with scattered flat, white to tan, adhering caseonecrotic plaques of varying size, sometimes with umbilicated centres, in single or multiple air sacs (Figure
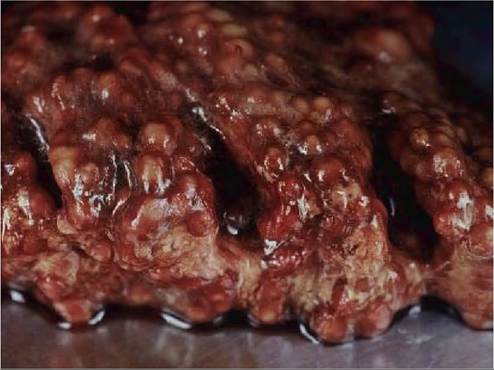
FIGURE 38.1 Acute aspergillosis caused by Aspergillus fumigatus in an immature bean goose (Anser fabalis). The lung contains large numbers of coalescing pale nodules. Photo: Bengt Ekberg, SVA.
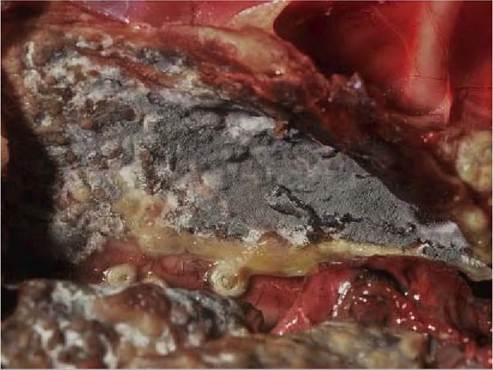
FIGURE 38.2 Severe chronic aspergillosis (Aspergillusfumigatus'') in a young white-tailed eagle (Haliaeetus albicilla). The air sac is lined by confluent, grey-green sporulating fungal growth. Photo: Bengt Ekberg, SVA.
38.2). In advanced cases, there is extensive, confluent mycotic growth with severely thickened air sacs, accumulation of fibrinopurulent exudates within air sacs, and extension oflesions into surrounding tissues. Fungal reproduction within air sacs is a common feature, upon which plaques become velvety and change colour to shades of green, olive, brown or black depending on the fungal species involved, which leads to further dispersal of conidia within the airways. Gross lesions of varying age and severity may present simultaneously in the same bird. Asphyxiation from focal invasive growth in proximal airways may occur.
Histopathologically, lung lesions in the acute forms of aspergillosis consist of pleuritis, interstitial pneumonia with infiltrates consisting of predominantly heterophils, admixed with lymphocytes, macrophages and fibrin, and coalescing necrotic to pyogranulomatous lesions, which efface the normal pulmonary architecture and compress surrounding tissue. Pyogranulomas have an eosinophilic centre consisting of degenerating heterophils and/or central necrosis, surrounded by heterophils admixed with lymphocytes, and palisading epithelioid and multinuclear giant cells at the periphery. The amount of fibrous tissue at the periphery of pyogranulomas increases as lesions age. Severe suppurative and/or necrotizing pneumonia sometimes occurs in birds. Air sac membranes display infiltration with heterophils, macrophages and to a lesser degree lymphocytes and plasma cells, prominent vasculature,
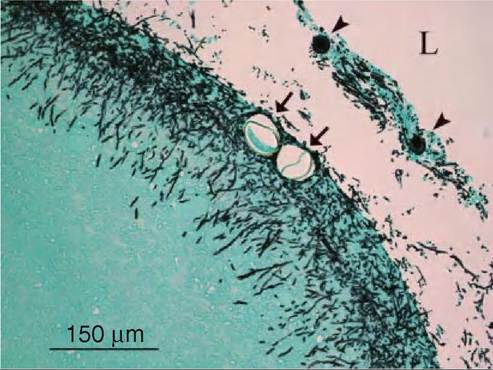
FIGURE 38.3 Hyphae (Aspergillus fumigatus) growing in a severely thickened air-sac membrane of a white-tailed eagle (the same bird as in Figures 38.2 and 38.4). Note the presence of nematode eggs (arrows) and conidiophores (arrowheads). L: air sac lumen. Grocotts methen- amine silver stain. Photo: Desiree Jansson.
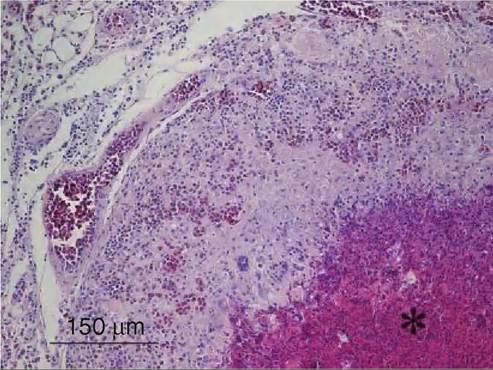
FIGURE 38.4 Microscopic appearance of lung tissue from a whitetailed eagle (the same bird as in Figures 38.2 and 38.3). A necrotic centre (*) of a pyogranuloma is seen in the lower right corner, and the lesion is surrounded by multinucleated giant cells and epithelioid macrophages. Note hyperaemia and the mixed interstitial inflammation. Haematoxylin and eosin. Photo: Desiree Jansson.
perivasculitis and oedema. As the disease progresses, air sac membranes become thickened with fibroplasia, massive leucocyte infiltration, formation of pyogranulomas and the presence of caseous exudates. Visualization of conidia, germinating conidia and hyphae is improved by various special stains (Figure 38.3). Fungal elements are predominantly located in pyogranulomas, in necrotic areas and within epithelioid and multinucleate giant cells. Hyphae appear as dichotomously branching, septate structures, having parallel cell walls. Signs of angioinvasion, i.e. hyphal invasion of blood vessels, subendothelial and mural mononuclear to polymorphonuclear invasion (Figure 38.4 ), thrombosis and coagulative necrosis may be observed in some birds. Upon sporulation in aerated tissues, conidiophores are present microscopically in exudates. Brain lesions are rarely reported from free-living birds, but when they occur they consist of solitary abscesses with necrotic centres and heterophil and macrophage infiltration at the periphery.
CLINICAL SIGNS AND TREATMENT
Affected free-living wild birds are usually found moribund or dead. When clinical signs are observed they are nonspecific and vary with the severity of the disease, but are often subtle. Severely affected birds may be lethargic, dehydrated, are unable to fly and withdraw socially. Respiratory signs include nasal discharge, bill opening, gasping, dyspnoea, changes in vocalization, accelerated breathing and cyanosis. Neurological signs such as ataxia, torticollis and opisthotonos are occasionally reported from wild birds.
Treatment of aspergillosis in birds generally meets with a low success rate because of the often advanced disease stage when the diagnosis is confirmed, the lack of pharmacokinetic data of antifungal drugs in birds, the failure of drugs to reach target tissues (especially encapsulated granulomatous lesions), the fact that birds possess elaborate airways and pneumatized bones to which the infection may spread, and the presence of concurrent diseases and immunosuppression. Moreover, drug resistance to antifungal drugs has been reported in A. fumigatus isolates of avian origin(24). Antifungal agents that may be used in birds include amphotericin B, terbinafine HCL, 5-fluorocytosine, itraconazole, fluconazole, ketoconazole, miconazole, clotrimazole, enilconazole and voriconazole. Drugs can be administered topically by nebulization, flushing of proximal airways and/or air sacs at surgery, or systemically by intravenous or oral routes, either alone or in different combinations. Surgical debridement of caseous material and granulomas may be attempted if the bird can tolerate anaesthesia.
DIAGNOSIS
Ante mortem diagnosis of aspergillosis is difficult and relies on a combination of clinical signs, culture and cytology on tracheal, bronchoalveolar or air sac lavage, haematology, serology, protein electrophoresis, endoscopic findings and radiology. A positive culture must be interpreted with caution because aspergilli are frequent airway contaminants. Heterophilic leucocytosis and increased β and γ globulins are indicative of aspergillosis. Enzyme-linked immunosorbent assays and galactomannan analysis (fungus-specific polysaccharide) have been applied to avian samples.
The post mortem diagnosis relies on gross findings and microscopic features of the characteristic hyphal growth pattern in wet mounts in 20% KOH and ink dye, or by histology. Confirmation is achieved by isolation on myco- logic media such as Sabouraud dextrose agar, Czapek Dox agar or potato dextrose agar. Diagnostic key characters should be evaluated using standardized culture conditions, and include characteristics such as the colour of the aerial mycelium and conidia, appearance of the colony margin and surface texture, growth rate in vitro, thermotolerance and, in particular, the size and morphology of conidiophores. Most aspergilli are fast-growing and display a powdery to cotton-like texture. The colour of colonies is highly variable between species. Hyphae and conidiophores may be collected from culture plates by cellophane tape, which is placed in a drop of lactophenol cotton blue and examined microscopically. Immunohistochemistry on formalin-fixed tissues can be used retrospectively. Molecular methods, especially polymerase chain reaction on DNA extracted from cultures, are useful tools for species identification.
MANAGEMENT, CONTROL AND REGULATIONS
Prevention is by far the best way to control high-risk situations such as captivity of wild birds in zoological exhibits, in relocation programmes or at rehabilitation centres. Animal facilities, transport crates, incubators and hatchers should be cleaned and disinfected with antifungal agents before use, and potential sources of conidia such as mouldy litter materials and feed should never be introduced. Care should be taken to minimize any stress. Incubators and areas around feeders and drinkers should be kept clean and dry to prevent fungal growth. To prevent outbreaks in wildlife, mouldy agricultural waste should be covered, ploughed under or cut and removed. Bird feeders should be kept clean and dry.
PUBLIC HEALTH CONCERN AND SIGNIFICANCE AND IMPLICATIONS FOR ANIMAL HEALTH
Aspergilli are rarely pathogenic to healthy people and domestic animals, with the exception of poultry in heavily contaminated environments. However, care should be taken to avoid heavy environmental exposure of humans and domestic animals to conidia. Protective face masks should preferably be used when removing mouldy feed and litter from poultry barns and stables, when visiting densely populated seabird colonies, and when performing necropsies on affected animals. Sporulating cultures should be handled with care at the laboratory.
REFERENCES
1. O’Gorman, C.M., Fuller, H.T. & Dyer, P.S. Discovery of a sexual cycle in the opportunistic fungal pathogen Aspergillus fumigatus. Nature. 2009;457:471-4.
2. Pringle, A., Baker, D.M., Platt, J.L., Wares, J.P., Latge, J.P. & Taylor, J.W Cryptic speciation in the cosmopolitan and clonal human pathogenic fungus Aspergillus fumigatus. Evolution. 2005;59:1886-99.
3. Nierman, WC., Pain, A., Anderson, M.J., et al. Genomic sequence of the pathogenic and allergenic filamentous fungus Aspergillus fumigatus. Nature. 2005;438:1151-6.
4. Debeaupuis, J., Sarfati, J., Chazalet, V. & Latge, J. Genetic diversity among clinical and environmental isolates of Aspergillus fumigatus. Infection and Immunity. 1997;65:3080-5.
5. Alvarez-Perez, S., Mateos, A., Dominguez, L., Martinez-Nevado, E., Blanco, J.L. & Garcia, M.E. Polyclonal Aspergillus fumigatus infection in captive penguins. Veterinary Microbiology. 2010;144:444-9.
6. Burgisser, H. Mycoses nasales chez le chevreuil. Schweizer Archiv fur Tierheilkunde. 1955;97:434-8.
7. Munro, R., Hunter, A.R., Bonniwell, M., & Corrigall, W Systemic mycosis in Scottish red deer (Cervus elaphus}. Journal of Comparative Pathology. 1985;95:281-9.
8. Domingo, M., Visa, J., Pumarola, M. et al. Pathologic and immunocytochemical studies of morbillivirus infection in striped dolphins (Stenella coeruleoalba). Veterinary Pathology. 1992;29:1-10.
9. Dagleish, M.P., Foster, G., Howie, F.E., Reid, R.J. & Barley, J. Fatal mycotic encephalitis caused by Aspergillus fumigatus in a northern bottlenose whale (Hyperoodon ampullatus). The Veterinary Record. 2008;163:602-4.
10. Prahl, S., Jepson, PD., Sanchez-Hanke, M., Deaville, R. & Siebert, U. Aspergillosis in the middle ear of a harbour porpoise (Phocoena phoc- oena). Mycoses. 2010;54:e260 4.
11. Hulphers, G. & Lilleengen, K. Mogelsvampsinfektion, aspergillos, hos hare och vilt levande fagel. Svensk VeterinUrtidskrift. 1947;52:235-9.
12. Kunkle, R.A. & Rimler, R.B. Pathology of acute aspergillosis in turkeys. Avian Diseases. 1996;40:875-86.
13. Young, E.A., Cornix, T.E. & Little, S.E. Concomitant mycotic and verminous pneumonia in a blue jay from Georgia. Journal of Wildlife Diseases. 1998;34:625-8.
14. Kaliner, G. & Cooper, J.E. Dual infection of an African fish eagle with acid-fast bacilli and an Aspergillus sp. Journal of Wildlife Diseases. 1973;9:51-5.
15. Souza, M.J. & Degernes, L.A. Mortality due to aspergillosis in wild swans in northeast Washington State, 2000-2002. Journal of Avian Medicine and Surgery 2005;19:98-106.
16. Jung, K., Kim, Y., Lee, H. & Kim, J. Aspergillus fumigatus infection in two wild Eurasian black vultures (Aegypius monachus Linnaeus) with carbofuran insecticide poisoning: a case report. Veterinary Journal. 2009;179:307-12.
17. Carrasco, L., Lima, J.S., Halfen, D.C., Salguero, FJ., Sanchez-Cordon, P. & Becker, G. Systemic aspergillosis in an oiled Magellanic penguin (Spheniscus magellanicus). Journal of Veterinary Medicine Series B. 2001;48:551-4.
18. Olias, P, Gruber, A.D., Bohmer, W, Hafez, H.M. & Lierz, M. Fungal pneumonia as a major cause of mortality in white stork (Ciconia ciconid). Avian Diseases. 2010;54:94-8.
19. Bozza, S., Gaziano, R., Spreca, A. et al. Dendritic cells transport conidia and hyphae of Aspergillus fumigatus from the airways to the draining lymph nodes and initiate disparate Th responses to the fungus. Journal of Immunology. 2002;168:1362-71.
20. Askew, D.S. Aspergillus fumigatus: virulence genes in a street-smart mold. Current Opinion in Microbiology. 2008;11:331-7.
21. Balloy, V. & Chignard, M. The innate immune response to Aspergillus fumigatus. Microbes and Infection. 2009;11:919-27.
22. Brakhage, A.A., Bruns, S., Thywissen, A., Zipfel, P.F. & Behnsen, J. Interaction of phagocytes with filamentous fungi. Current Opinion in Microbiology. 2010;13:1-7.
23. Harmon, B.G. Avian heterophils in inflammation and disease resistance. Poultry Science. 1996;77:972-7.
24. Beernaert, L.A., Pasmans, F., Van Waeyenberghe, L. et al. Avian Aspergillus fumigatus strains resistant to both itroconazole and voriconazole. Antimicrobial Agents and Chemotherapy. 2009;53:2199-201.