Chapter summary
Respiratory system
1. The respiratory and circulatory systems are the means by which nutrients are delivered to each cell and wastes are removed.
2. The functions of the respiratory system include gas exchange, regulation of blood pH, olfaction, filtration of inspired air, production of sounds, and elimination of some water and heat via expired air.
3. Functionally, it can also be divided into two sections: (1) The conducting portion consists of a series of connected tubes that filters, warms, moistens, and conducts air to and from the lungs; (2) the respiratory portion is the site of gas exchange between the air and blood, and consists of the respiratory bronchioles, alveolar ducts, alveolar sacs, and alveoli.
Functional anatomy
Nose
1. The nose is the externally visible portion of the respiratory system. The external nares (nostrils) are the external openings to the respiratory tract. The philtrum is the area between the lips and nose.
2. The nasal cavity extends from the external nares to the caudal nares, and is separated from the mouth by the hard and soft palates.
3. The nasal cavity communicates with the paranasal sinuses, and posteriorly with nasopharynx through two openings called the internal nares or choanae.
4. The paranasal sinuses are air-filled cavities within some bones of the skull. The major ones are the frontal and maxillary sinuses.
The pharynx
The pharynx connects the nasal cavity and mouth to the larynx and esophagus, respectively. It directs food and air into the digestive and respiratory systems. The pharynx is divided into the oropharynx and nasopharynx, and the common caudal portion is the Iaryngopharynx.
Larynx
1. The larynx connects the Iaryngopharynx with the trachea and contains the vocal cords. The two functions of the larynx are (1) to provide a routing mechanism for air and food, and (2) to make sounds.
2. The larynx is formed by five cartilages including single epiglottic, thyroid, and cricoid cartilages, and paired arytenoid cartilages. The epiglottic cartilage closes the opening to the larynx during swallowing.
Trachea
1. The trachea extends from the larynx to the right and left primary bronchi above the base of the heart. It consists of four layers: (1) mucosa, which is the deepest layer; (2) submucosa; (3) hyaline cartilage; and (4) the adventitia, the most superficial layer composed of areolar connective tissue.
2. The trachea contains a series of dorsally incomplete, "C"-shaped hyaline cartilage rings.
Bronchi
1. The trachea divides into the right and left primary bronchi.
2. The primary bronchi contain incomplete cartilaginous rings and divide into the smaller secondary bronchi. These keep dividing into the following sequence of channels: tertiary, or segmental, bronchi → bronchioles → respiratory bronchioles → alveolar ducts → alveolar sac → alveoli.
Lungs
1. In general, the left and right lungs have two and four lobes, respectively. The horse has three right lobes.
2. The cranial portion, or apex, of each lung is located in the thoracic inlet, while the base is the caudal end of the lung resting on the diaphragm.
3. The cardiac notch is the indentation between the lobes where the heart makes contact with the lung.
Pleural membrane
1. The lungs are surrounded by a serous membrane called the pleural membrane. The narrow parietal space between these two layers contains a small amount of pleural fluid that allows the two layers to slide over one another during breathing. Inflammation of the pleural membrane is called pleurisy.
2. The pressure in the intrapleural space is negative. This negative pressure inside the intrapleural space is vital for the expansion of the lungs. If an injury to the chest wall punctures the pleural membrane, it can result in a pneumothorax.
The lung on that side can collapse.Alveoli
1. Surrounding the alveolar ducts are many alveoli and alveolar sacs. Alveoli walls contain predominantly type I alveolar cells, which are the main site of gas exchange.
2. Alveolar walls also contain type II alveolar cells, alveolar macrophages, and fibroblasts that produce reticular and elastic fibers. Type II alveolar cells secrete surfactant.
3. The respiratory membrane is where O2 and CO2 diffuse across the alveolar and capillary walls. It is a very thin membrane about 0.5 μm thick and consists of four layers.
Blood supply to the lungs
1. The pulmonary and bronchial arteries supply blood to the lungs. The pulmonary artery carries deoxygenated blood through the pulmonary trunk and into the right and left pulmonary arteries. Oxygenated blood returns to the left atrium via the pulmonary veins.
2. Bronchial arteries arise from the aorta and deliver oxygen to the lungs. They mostly perfuse the bronchi and bronchioles.
Pulmonary ventilation
Respiration occurs in three steps:
1. Pulmonary ventilation, or breathing, is the mechanical movement of air into (inspiration) and out (expiration) of the lungs.
2. External respiration is the exchange of gases between the lungs and the pulmonary capillaries.
3. Internal respiration is the exchange of gases between systemic capillaries and tissue.
Pressure changes during respiration
Inspiration
1. Boyle's law states that there is an inverse relationship between the volume and pressure of a gas: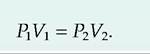
The pressure inside the lungs decreases as the volume of the lungs increases. Air moves into the lungs as a result of a decrease in air pressure within the lungs. In most animals, inspiration is an active process.
2. The main muscles responsible for quiet inhalation are the diaphragm and external intercostal muscles. As an animal increases the force of inspiration, additional muscles including the sternocleidomastoid muscles, scalene muscles, and pectoralis minor muscles are involved.
Expiration
Normal expiration is a passive process involving no active muscle contraction.
Other factors involved in pulmonary ventilation
1. Surface tension of alveolar fluid. Alveolar fluid coats the inside surface of the alveoli. In order to expand the lungs, the surface tension of the alveolar fluid must be exceeded. This surface tension accounts for approximately two-thirds of the elastic recoil of the lungs.
2. Compliance of the lungs. The distensibility of the lungs is referred to as lung compliance. High lung compliance means that the lungs will expand easily. Lung compliance is related to (1) the distensibility of lung tissue and the thoracic cage and (2) alveolar surface tension. Compliance can be decreased by several factors: (1) scar tissue formed in the lungs as a result of certain diseases, (2) pulmonary edema resulting from the accumulation of fluid in the lungs, (3) insufficiency of surfactant, and (4) a decrease in the ability of the thoracic cage to expand.
3. Airway resistance. The flow of air into the lungs is inversely related to airway resistance:
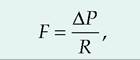
where F is gas flow, P is pressure, and R is resistance. The larger the diameter of the airway, the less the resistance. Stimulation of the sympathetic nervous system causes relaxation of these walls, which allows air to more readily enter the lungs. Diseases or injuries to the airways can increase airway resistance.
Lung volumes and capacities
Respiratory volumes
1. The respiratory capacity, or the amount of air that moves in and out of the lungs, depends on
the strength of inspirations and expirations. During normal, quiet breathing, the volume of air moving in and out of the lungs is called the tidal volume (TV). Only about 70% of the tidal volume reaches the lungs.
2. The remaining portion of the air is found in the airways including the nose, pharynx, larynx, trachea, bronchi, bronchioles, and terminal bronchioles.
These airways are collectively called the anatomical dead space.3. The minute volume (MV) is the volume of air inhaled and exhaled each minute, and is calculated as

4. The respiration rate varies by species. Because of the anatomical dead space, not all of the minute volume is available for gas exchange.
5. The alveolar ventilation rate (AVR) is that portion of the tidal volume that actually reaches the site of gas exchange:
AVR = respiration rate ? (TV-dead space). (mL∕min) (breaths/min) (mL/breath)
6. If an animal more forcefully inhales, it can increase the volume of air entering the lungs above normal tidal volume. The additional inhaled air is called the inspiratory reserve volume. Similarly, an animal can force more air out of its lungs than occurs during quiet respiration. This additional volume exhaled is called the expiratory reserve volume. Following a forced expiration, the air remaining in the lungs is the residual volume.
Exchange of oxygen and carbon dioxide
Dalton's law and Henry's law
1. The exchange of oxygen and carbon dioxide between alveolar air and pulmonary blood is a passive process, which is explained by two gas laws. Dalton's law explains how gases move by diffusion based on pressure differences, while Henry's law describes the diffusion of gas based on its solubility.
2. Dalton's law states that each gas within a mixture exerts its own pressure independent of the other gases present. The pressure of an individual gas is called its partial pressure, designated Px. The total pressure of a gas mixture is calculated by summing all its partial pressures.
3. Henry's law says that the quantity of a gas that will dissolve in a liquid is proportional to its partial pressure and its solubility coefficient. Therefore, gases will dissolve in body fluids more readily if they have a greater partial pressure and solubility coefficient.
The solubility coefficient of CO2 is 24 times higher than that of O2. Therefore, CO2 dissolves in blood more readily than O2. In contrast, the solubility of nitrogen is very low, so even though atmospheric air has 79% N2, it has very little effect on body functions.External and internal respiration
1. External respiration, also called pulmonary gas exchange, is the diffusion of O2 and CO2 from the alveoli to pulmonary blood. Pulmonary blood is deoxygenated blood arriving from the right ventricle. Blood circulating through the body picks up CO2 and delivers O2. As this blood travels through the pulmonary capillaries, CO2 diffuses into the alveoli while O2 diffuses from the alveoli to pulmonary blood. The exchange of these gases occurs independently, and passively.
2. Pulmonary gas exchange is facilitated by a very thin respiratory membrane.
3. In addition, there is a close association between the amount of gas reaching the alveoli, that is, ventilation, and the blood flow through the pulmonary capillaries, that is, perfusion. When ventilation becomes inadequate within alveoli, the Po, will decrease. This causes an autoregulatory response in which pulmonary arterioles constrict. Conversely, if Po, increases, then the pulmonary arterioles dilate allowing more blood to flow to those areas that can maximize gas exchange. Note that this is the opposite of what happens in systemic circulation where a decrease in Po, results in vasodilation.
4. Internal respiration, or systemic gas exchange, occurs at the tissue level where there is an exchange of O2 and CO2 between systemic capillaries and tissue. O2 diffuses from the capillaries into the cells while CO2 diffuses from the cells into the systemic capillaries.
Transport of oxygen and carbon dioxide
Oxygen transport
1. Oxygen has a low solubility coefficient, so it does not readily dissolve in blood. Instead, over
98% of O2 is bound to hemoglobin. The heme portion of hemoglobin contains four atoms of iron, each able to bind to one molecule of O2. Oxygen bound to hemoglobin forms oxyhemoglobin. Hemoglobin that has released O2 is called reduced hemoglobin, or deoxyhemoglobin. Because it picks up a H+ ion after releasing O2, reduced hemoglobin is abbreviated as HHb:
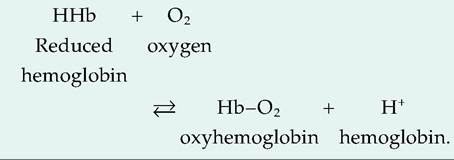
2. Pch is the most important factor controlling how much O2 is bound to hemoglobin. When reduced hemoglobin is all converted to oxyhemoglobin, it is fully saturated.
3. When an animal is at rest, hemoglobin only releases approximately 25% of its oxygen. It maintains a reserve that is available when needed. If the animal begins vigorous exercise, hemoglobin is able to respond by releasing a greater amount of O2.
Other factors affecting the oxygen-hemoglobin dissociation curve
1. Other factors influence oxygen-hemoglobin dissociation.
a. pH of the Blood. Increasing the acidity of blood, that is, lowering the pH, lowers the affinity of hemoglobin for O2. Therefore, as the metabolism of tissue increases, resulting in increased lactic and carbonic acid at the same l∖h, hemoglobin releases more O2 at that site. This is termed the Bohr effect.
b. Temperature. An increase in temperature, like increased acidity, is a by-product of increased metabolism. Increased metabolism requires additional O2, so shifting the dissociation curve to the right provides necessary O2.
c. BPG. An increase in the production of BPG, formerly called diphosphoglycerate, liberates more O2. Increased production of BPG is also associated with increased metabolism.
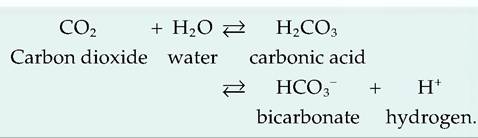
d. Pco2- A decrease in pH acts similarly to an increase in Pco2- As shown next, CO2 can react with water to form carbonic acid, which then dissociates to form bicarbonate and H+. Thus, increased lfo. is associated with decreased pH:
Hemoglobin-nitric oxide
1. Nitric oxide (NO), a gas, plays an important role in vasomotor tone. It is a potent vasodilator. Produced in lung and endothelial cells, NO can be carried by hemoglobin.
2. The binding of O2 to hemoglobin causes a change in the conformation of hemoglobin allowing NO to bind to the cysteine of hemoglobin. This protects NO from being broken down by the Fe in hemoglobin. As oxyhemoglobin releases its O2, it simultaneously releases NO. NO dilates local blood vessels, thus further aiding in supplying O2 to areas in need.
Carbon dioxide transport
Carbon dioxide is a waste product of metabolism. It is transported in the blood to the lungs in three forms:
1. Dissolved CO2. Accounting for the smallest amount of transported CO2, 7-10% is carried dissolved in the plasma.
2. Carbamino compounds. Approximately 20% of CO2 is transported in the red blood cells attached to the amino acids of globin forming Carbaminohemoglobin:

3. Bicarbonate ions. Most carbon dioxide, about 70%, is transported in the blood as bicarbonate ions (HCO3 ).
Control of respiration: neural mechanisms
1. The medullary rhythmicity area is located in the medulla oblongata, and it controls the basic respiration rhythm. It consists of two areas, the inspiratory and expiratory areas, also called the dorsal respiratory group and ventral respiratory group, respectively.
2. The inspiratory area sends signals to the diaphragm via the phrenic nerves and to the external intercostal muscles via the intercostal nerves. These signals cause muscle contraction
resulting in inspirations. Transection between the spinal cord and the medulla oblongata stops breathing.
3. While not active during quiet breathing, forceful expiration requires signals from the expiratory area that cause contraction of the internal inter- Costals and abdominal muscles. Contraction of these muscles further decreases the volume of the thoracic cavity, thus increasing exhalation.
Pneumotaxic and apneustic areas
1. Located in the upper pons, the pneumotaxic area, also called the pontine respiratory group, sends inhibitory signals to the inspiratory area. These signals primarily function to prevent overfilling of the lungs.
2. Conversely, the apneustic area located in the lower pons sends stimulatory signals to the inspiratory area that prolongs inspiration. The pneumotaxic area can override the apneustic area.
Chemoreceptors
1. The respiratory system functions to bring in O2 and eliminate CO2 from the body. This function is assisted by specialized receptors called chemoreceptors that monitor the levels of CO2, O2, and H+, and then send such information to the respiratory center.
2. Respiratory chemoreceptors are located in several locations. Central chemoreceptors found in the medulla oblongata respond to changes in cerebrospinal fluid H+ and Pco-,- Peripheral chemoreceptors include the aortic bodies and carotid bodies whose removal eliminates a respiratory response to hypoxia.
3. The levels of CO2 and H+ are highly correlated. Throughout the body, CO2 is quickly converted to carbonic acid catalyzed by the enzyme carbonic anhydrase. Carbonic acid dissociates into HCO3- and H+. Therefore, increases in CO2 lead to increases in H+, while decreases in CO2 lead to decreases in H+. As a result, Pco„ has a large effect on respiration, whereas Po-, only affects respiration if its levels change substantially.
4. Increases in arterial blood CO2, called hypercapnia, cause an increase in H+. This has a large effect on central chemoreceptors which causes increased respiration rate, possibly causing hyperventilation. Low arterial blood CO2, called hypocapnia, inhibits respiration. Large drops in arterial Pq2 increase ventilation by stimulating peripheral chemoreceptors.
Pulmonary and airway receptors
Three types of sensory receptors have been identified in the lungs including slowly adapting stretch receptors, irritant receptors, and unmyelinated C fibers. The stretch receptors increase their firing rate as the lungs and larger airways inflate.
Avian respiration
Anatomy of the avian respiratory system
1. The lungs do not expand, but instead, air sacs act as bellows to move air in and out of the lungs.
2. Gas exchange occurs in the lungs, but not in the air sacs.
3. Birds have no diaphragm, and the thoracic cavity is at atmospheric pressure.
4. Sound is generated in the syrinx, which is composed of cartilage and vibrating soft tissue.
5. The avian lung is a rigid structure located in the thoracoabdominal cavity.
6. Tracheal volume in birds is about 4.5 times larger than in mammals, creating a larger dead space. Birds compensate by having a larger tidal volume and lower respiratory frequency.
7. The primary and secondary bronchi do not participate in gas exchange. The parabronchi, also called tertiary bronchi, originate from the secondary bronchi, and are the site of gas exchange.
8. There are two types of parabronchi. The paleo- pulmonic parabronchi appear as parallel tubes extending from the secondary bronchi, and gas flow is unidirectional.
9. There is also an irregular branching system of neopulmonic parabronchi through which gas flow is bidirectional.
Air sacs
1. Air sacs are poorly vascularized, thin membranous structures connected to either the primary or secondary bronchi. They do not participate in gas exchange, but function to move air through the lungs so that gas exchange can occur in the parabronchi.
2. Air sacs can also extend into selected bones.
3. Birds possess nine air sacs; four are paired and one is unpaired.
Avian ventilation and gas exchange
1. During inspiration, approximately half of the tidal volume first enters the caudal air sacs and half enters the cranial air sacs. About half of this air goes through the neopulmonic lung continuing directly to the caudal thoracic and abdominal air sacs. The other half goes into the mediodorsal secondary bronchi and into the paleopulmonic parabronchi, and then into the cranial group of air sacs.
2. During expiration, contraction of the expiratory 3. muscles causes air to flow out of the caudal tho
racic and abdominal air sacs. This gas passes through the neopulmonic lungs to the paleopulmonic lungs. Simultaneously, air leaves the cranial air sacs through the medioventral secondary bronchi flowing into the primary bronchus and the trachea to exit the body. Therefore, all air moves through the paleopulmonic parabronchi, and air passes through the paleopulmonic parabronchi in a caudal-to-cranial direction while traveling through the neopulmo- nary parabronchi bidirectionally.
Gas exchange occurs between the air capillaries and blood capillaries.
^∙s>2⅛> Review questions and answers are available a online.
References
Constantinescu, G.M. 2001. Guide to Regional Ruminant Anatomy Based on the Dissection of the Goat. Iowa State Press, Ames, Iowa.
Constantinescu, G.M. 2002. Clinical Anatomy for Small Animal Practitioners. Iowa State Press, Ames, Iowa.
Constantinescu, G.M. and LA. Constantinescu. 2004. Clinical Dissection Guide for Large Animals, Horse and Large Ruminants, 2nd edition. Iowa State Press, Ames, Iowa.
Ikegami, M., A.H. Jobe, J. Newnham, D.H. Polk, K.E. Willet, and P. Sly. 1997. Repetitive prenatal glucocorticoids improve lung function and decrease growth in preterm lambs. Am. J. Respir. Crit. Care Med. 156: 178- 184.
Sturkie, RD. 1986. Avian Physiology, 4th edition. Springer- Verlag, New York.
Swenson, JJ. and W.O. Reece. 1993. Duke's Physiology of Domestic Animals, Ilth edition. Cornell University Press, Cornell University, Ithaca, New York.
Whittow, G.C. 2000. Sturkie's Avian Physiology, 5th edition. Academic Press, San Diego, California.