EPIDEMIOLOGY
Lyssaviruses are zoonotic infections that invariably spill over into non-maintenance hosts (such as humans, ruminants and cats). Onward transmission within these deadend hosts does not occur, so the successful transmission of RABV in new host species is likely to represent a major adaptive challenge.
This is, in part, a reflection of the strong selective constraints that act on RABV, resulting in a high rate of deleterious mutation and hence in relatively low rates of non-synonymous substitution (nucleotide substitutions leading to amino acid changes), including at sites that might potentially enhance fitness. At a higher level, both the N and G gene phylogenies indicate that viruses sampled from other species of the family Canidae, such as foxes and raccoon dogs, as well as hosts belonging to other families within the Carnivora, e.g. the Herpestidae in southern Africa, are included in the phylogenetic diversity of dog RABV. Although no significant evidence for adaptive evolution has been found, recent studies strongly suggest that the dog has served as the main bridge for inter-species RABV liaison, generating viral lineages that then spread to other taxa(1). Determining the genetic basis of the traits that govern cross-species transmission clearly represents a major goal for future research on RABV, although it is worth noting that patterns of cross-species transmission may also be influenced by the ecological factors that shape host contact rates.Infectious Diseases of Wild Mammals and Birds in Europe, First Edition. Edited by Dolores Gavier-Widen, J. Paul Duff, and Anna Meredith. © 2012 Blackwell Publishing Ltd. Published 2012 by Blackwell Publishing Ltd.
GEOGRAPHIC DISTRIBUTION IN EUROPE
Rabies caused by RABV is widely distributed throughout the world and is present in all continents except Australia and Antarctica.
In Europe, canine rabies has persisted for centuries. By the beginning of the 20th century, strict implementation of hygienic measures (dog supervision, feral dog control, mandatory carrying of muzzle when appropriate) resulted in the virtual disappearance of dog- mediated rabies, but the disease re- emerged in a focus south of Kaliningrad during World War II. The red fox was identified as the main reservoir for the virus, and this species, in subsequent decades, was responsible for the widespread dissemination of the disease. By the mid- 1970s, fox rabies (also referred to as sylvatic or wildlife- mediated rabies), affected almost all countries within Western and Southeastern Europe. The incidence of fox rabies peaked during the early 1980s with around 16,000 to 25,000 annual cases and at this time the disease also reached its most widespread geographical distribution. Nevertheless, large regions in Europe have never experienced cases of fox rabies, or have had a long history of freedom from the disease. In addition, as a result of the oral rabies vaccination (ORV) of wildlife, since the late 1980s, the incidence of rabies in other parts of Europe has decreased dramatically. Within the last 25 years, fox rabies has been virtually eliminated in Western Europe and substantially decreased in Central European countries. At present (Figure 6.1), endemic areas are mainly restricted to the eastern and southern parts of Europe (the Baltic countries, Belarus, Russia, Ukraine and Moldavia) and the Balkan region. However, the recent emergence, or reemergence, of fox rabies clearly demonstrates that this apparent steady decline in the number of cases is not inevitable: Italy, for instance, having been rabies-free, had cases of fox- associated rabies in 2008—2010, originating from the endemic regions of the Northwest Balkans.Rabies in raccoon dogs predominantly occurs in northeastern parts of Europe, e.g. the Baltic countries, Belarus, Russia and the Ukraine, and it is linked to a high population density of this host species, and to a specific variant of the fox rabies virus.
Dog-mediated rabies (synonym ‘canine rabies’) in Europe still persists in Turkey. At present, it is unclear whether a dog rabies strain still exists in dogs in regions of Eastern Europe such as Russia (where RABV infections are notified in dogs and other domestic animals). In addition, sporadic cases of dog rabies are periodically reported in rabies- free regions as a result of importation of dogs incubating the disease from rabies-endemic countries. In some of these introductions of RABV by a single infected dog, secondary cases result, which can create short ‘chains’ of transmission that, fortunately, finish in a dead-end host outbreak. These incidents legitimately result in the loss of rabies- free status of the country in which they occur (a problem encountered in France in 2008).
In addition to two specific lyssavirus genotypes (EBLV1 and EBLV2, mapped without distinction on Figure 6.2), a West Caucasian bat lyssavirus (WCBV), has been found in the European part of the Caucasus. The vast majority of bat rabies cases detected in Western Europe were identified as EBLV1. Further molecular characterization revealed the existence of phylogenetic sublineages 1a and 1b. While EBLV1a seems to occur in the lowlands and coastal plains along the North or Baltic Seas, and in Central Europe, sublineage 1b is restricted to the Iberian Peninsula, France and the Netherlands. Field surveys in Spain suggested that this virus is endemic in bat colonies. By contrast, EBLV2 has only been found sporadically in the Netherlands, UK, Switzerland, Germany and Finland(2). Sero-surveys of bat populations performed in the UK suggest that EBLV2 is
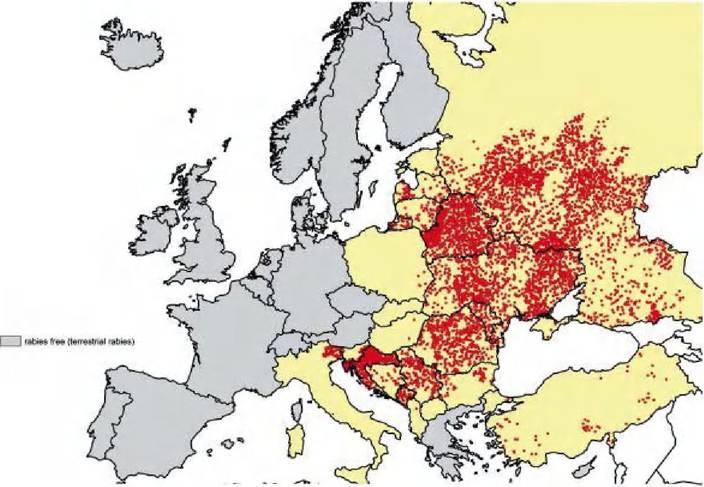
FIGURE 6.1
Rabies in terrestrial mammals; the situation in Europe in 2009.
FIGURE 6.2
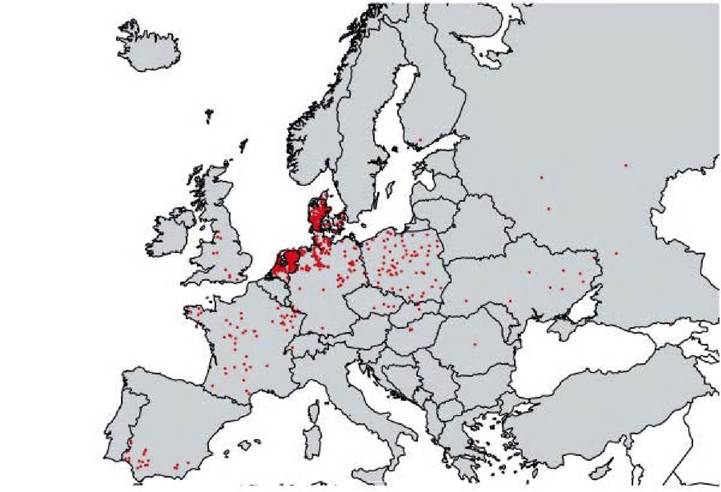
Bat rabies cases in Europe from 1977 to 2009, as reported to the World Health Organization.
also endemic with a low prevalence.
Considering the large distribution area of the suggested main reservoir species in Eurasia, ranging from Western Europe to Central Asia, there is reason to believe that infections of bats with those bat lyssaviruses could be far more widespread across Europe than observed by current surveillance.SPECIES-RELATED SUSCEPTIBILITY
Only mammals are currently considered susceptible under natural conditions to rabies virus. However, species susceptibility is variable and, to a lesser extent, relates to individual factors such as immunological status. For rabies, the identification of susceptibility and resistance factors is difficult because of confusion with factors such as virus variants and ecological characteristics of the host that affect exposure risk and clinical outcome. Nevertheless, species affected by rabies can be classified into high (order Carnivora — canid, mephitid, procyonid, viverrid — or Chiroptera), moderate (felid, mustelid, ungulate (Artio- dactyla) or primate) and low-receptivity/susceptibility (insectivore, lagomorph, monotreme, marsupial or rodent) species(3). The susceptibility to RABV infection of marine mammals (cetaceans, pinnipeds) is unknown.
From an epidemiological point of view, each species either belongs to a reservoir or spillover host category, at least for a period of time. As a general rule, highly susceptible species are usually reservoir species; moderate- and low-receptive species are usually considered to be spillover hosts.
Towards the end of the 20th century, in Europe, the wildlife species most frequently affected with rabies has been the red fox, accounting for 75—80% of all recorded cases (90—94% of all wildlife cases). In addition, rabies cases were also reported in highly susceptible species, e.g. in raccoon dogs, jackals (Canis latrans) or wolves (Canis lupus), all other canids, and in the invasive raccoons (Procyon lotor, a procyonid translocated from North America). Infection in a wide range of other wildlife and domestic species has been reported, usually in a spatial and temporal association with fox rabies, e.g.
stone martens (Martes foina) and pine martens (Martes martes), badgers (Meles meles) and other mustelids, and the roe deer ( Capre- olus capreolus), as well as in domesticated animals (dogs, cats and cattle, accounting for 89—95% of rabies cases in domesticated animals during recent decades); these are all considered to be spillover infections from foxes because all other forms of rabies disappeared in those areas where fox- mediated rabies was successfully eradicated. In recent decades, in the Baltic countries the epidemiological situation has changed, with the raccoon dog accounting for nearly half of the reported cases.In Turkey, the domestic dog is the main reservoir species, and cases in other species — wildlife, domestic animals or humans — are the result of spillover events from dogs.
Bat rabies cases in Europe involve infectious cycles that are independent from sylvatic and canine rabies, and the epidemiology is significantly different. Out of 45 known indigenous bat species present in Europe, rabies infection has been detected in only 10 species. EBLV1 has been detected most frequently in serotine bats (Eptesicus serotinus, Eptesicus isabellinus), which are therefore considered to play a key role in the maintenance of this virus. Cases of EBLV1 have been found in bat species by using the standard fluorescent antibody test (FAT) for rabies diagnosis. Moreover, there is evidence of infection of other bat species (Nyctalus noctula, Myotis myotis, Pipistrellus nath- usi), Pipistrelluspipistrellus and Plecotus auritus) by EBLV1, including some infected live individuals. Therefore, the role of the different bat species (as reservoirs, or perhaps victims of the active circulation of EBLV1 in other bat species) remains to be clarified.
By contrast, EBLV2 has been found only sporadically in Myotis bats (an abundant genus of bat species in Europe), e.g. M. daubentonii and M. dascyneme. The recently described WCBV has only been isolated once from a Schreiber’s bat (Miniopterus schreibersi) in the European part of the Caucasus.
I nfection of mammals other than bats with EBLV is rare. To date, occurrences of EBLV1 have been reported in a limited number of terrestrial mammals, including five domestic sheep, a stone marten and two domestic cats. Cases of EBLV transmission to humans were recently reported in Europe: since 1985, three and two human deaths from EBLV1 and EBLV2, respectively, have been suspected or confirmed. The suboptimal level of passive surveillance in wildlife species in many countries of Europe on the one hand and the limitation of investigation in species protected by law on the other hand, hinder the compilation of an accurate picture of affected bat species and associated host factors. This is in stark contrast to the situation in the Americas, where it was shown that almost all bat species are infected with bat-associated rabies viral strains.
ENVIRONMENTAL FACTORS
Sylvatic rabies in Europe is intrinsically linked to the behavioural ecology of its reservoir species, the red fox, and the interactions of parameters of two major biological associations, the virus-host and host-environment rela- tionships(4). The population density of reservoir species is mainly affected by the carrying capacity of an area, which in turn depends on its food resources, shelter availability and social regulation. A high population density of susceptible hosts increases the incidence but not the spread of rabies. In epizootic situations, the speed of colonization of new areas by RABV infection is narrowly dependent on the size of the home ranges of the host mammal species(5).
In the European Arctic tundra, red-fox-mediated rabies is absent. Here another reservoir species, e.g. the arctic fox (Alopex lagopus) fills the ecological niche, in particular in the tundra regions bordering Siberia. In the tundra, the rabies virus variant infecting the arctic fox is slightly different from that of the red fox and is closer to the dog virus from which it derived, probably in central Asia.
As the red fox is considered an opportunistic species, it can adapt and benefit from human activities. Anthropogenic changes, e.g. in the agricultural use of land, fragmentation of landscape, or urbanization, provide additional food resources and result in increased fox densities and colonization of new territories. Urban fox populations for instance, long considered a British phenomenon, have been increasing in Western and Central Europe for the past two decades. They posed a particular challenge for the final phase of rabies elimination in Germany when residual foci of the disease persisted in areas of high fox density in the southwest of the country1-6). The voluntary, or unintentional, moving of animals by people, sometimes when the animals are in a latent phase of infection, can influence the spread of rabies, and although in Europe translocation of incubating foxes was a rare occurrence, translocation of the raccoon dog from Asia to Europe, or, to a lesser extent, the raccoon from North America, effectively provided new susceptible host ‘niches’ for rabies virus transmission.
Natural barriers such as rivers or mountain ranges only temporarily block the spread of sylvatic rabies. In high mountainous areas such as the Alps, the spread of sylvatic rabies has been shown to follow the valleys. If the infection pressure is high enough, even large rivers or high mountains are crossed at their ‘weakest points’, e.g. at bridges, along lower mountain passes, or between end-to-end valleys. Nevertheless, natural barriers should be taken into account when implementing ORV campaigns in wildlife. Although regulated in Europe, translocation of wildlife may substantially contribute to the spread of the disease over large geographic distances, as was shown for the Americas.
Similar environmental pressures apply to dog-mediated rabies. However, canine rabies is more human-associated, as the dog is the principle reservoir and has a close relationship with humans. As a consequence of cultural, religious and social conditions, the incidence and spread of canine rabies in endemic areas is influenced by the social structure of the dog populations in those areas - that is, the owned, ownerless (community) or feral dog status.
European bat rabies is different from ‘terrestrial’ rabies, and the epidemiological traits relate to the abundance of different bat species in Europe, their different species behaviours and migration habits. As some bat species are migratory, the possibility of virus introduction and wider dispersal is possible, and consequently rapid spread might be expected. Furthermore, bat roosts constantly change location, depending on the seasons, and this may affect and enhance virus transmission opportunities between bats. Therefore, natural barriers do not appear at present to play a role in the spread of bat rabies.
Epidemiological Role of the Affected Species
Reservoir host species play a key role in the epidemiology of sylvatic, canine and bat rabies. The continuation of the infection in endemic areas in Europe relies on independent infectious cycles, in which the associated viral variants are constantly and periodically circulating among conspecifics (see the section on molecular epidemiology, below).
Regarding sylvatic rabies in Europe, there are several arguments indicating that the red fox is the main reser- voir(4’5). However, other potential reservoir hosts, such as the raccoon dog as well as the raccoon, need careful evaluation. Both species were introduced into European Russia and Germany, respectively, during the last century and progressively spread across Europe. They are important reservoir species for rabies in their natural habitats in Asia and America, respectively. Both species when infected pose a high zoonotic potential and must be considered potential reservoir species for classical rabies viruses in Europe too. They are prime examples of a human- made ecological problem resulting in establishment of additional reservoirs of high zoonotic potential with almost no observed interspecies competition with the red fox. Although it is not yet proven, there is evidence that the raccoon dog could act as another rabies reservoir, as it is the second most reported species to be infected in Central and Baltic Europe, and it is mainly infected by one of the fox RABV variants, although its distribution is much larger in Europe than the distribution of this peculiar RABV variant. There are some areas with high populations of raccoons in Europe; however, fox rabies variants have not managed to adapt to this new host yet, and only sporadic spillover cases are reported.
Although spillover hosts, e.g. dogs and cats in the case of fox rabies, play an important role in the transmission of rabies to humans, this seems not to apply for bat- associated lyssavirus variants.
MOLECULAR EPIDEMIOLOGY
Both the N and G gene phylogenies indicate that viruses sampled from other species of the family Canidae (such as foxes and raccoon dogs), as well as hosts belonging to other Carnivora families, e.g. the Herpestidae in southern Africa, are included in the phylogenetic diversity of dog RABV. Although no significant evidence for adaptive evolution has been found, recent studies strongly suggest that the dog has served as the main link for inter- species RABV transmission, generating viral lineages that then spread to other taxa(1).
Dog-associated RABV form a distinct phylogenetic group among lyssaviruses of genotype 1, comprising six major clusters, identified as the Africa 2, Africa 3, Arctic- related, Asian, Cosmopolitan and Indian subcontinent clades(1). The Cosmopolitan clade included dog, wolf and fox isolates from Europe, the Middle East, Iran and Kazakhstan and further east to the Republic of Tuva in Russia. It also included a number of dog RABV isolates from the Americas and from North, Central and South Africa, suggesting that they probably represent secondary migrations from Eurasia.
Further, the lack of mixing between clades supports the idea that, over longer timescales, the persistence of RABV in its enzootic stage does not depend upon regular immigration of infected individuals1-7). Rather, it is more likely that the dispersal of RABV reflects the gradual spatial spread of virus between animals that themselves move relatively small distances, as previously demonstrated in Europe with red foxes and raccoon dogs and in North America with raccoons.
In Europe, phylogenetic analysis of the gene sequence data revealed a number of distinct groups, each associated with a particular geographical area. This resulting pattern suggests that the rabies virus has spread westwards and southwards in Europe during the 20th century, but that physical barriers such as the Vistula River in Poland have enabled localized independent evolution1-8). During this dispersal process, two species jumps took place: one into red foxes and the second into raccoon dogs, although it is unclear whether virus strains are preferentially adapted to particular animal species or whether ecological factors explain the occurrence of the phylogenetic groups. In Europe, there are only three terrestrial non-flying mammal species that are capable of sustaining an epidemiological cycle on their own. These are the European red fox, the invasive raccoon dog, originating from Asia, and the domestic dog, which are infected by different specific variants of RABV.