Functional anatomy of the digestive system
The digestive system shows great variation among species (Table 17.1). These variations in structure are necessary depending on whether the animal is a carnivore (meat-eating), herbivore (plant-eating), or omnivore (meat- and plant-eating).
A rabbit is a typical nonruminant example; its stomach and small intestine are relatively small, whereas the cecum is well developed in order to allow for microbial digestion. Nonruminant herbivores typically have a well-developed cecum since this is the primary site of cellulose digestion. Ruminants have a complex stomach that accommodates microbial digestion, a proportionately long small intestine, and a large colon. Carnivores, such as dogs and cats, have a short small intestine, poorly developed cecum, and average colon. The pig, which is an omnivore, has an intermediate size colon since this is a major site of microbial digestion.Further evolutionary adaptations have occurred in the stomach and gastrointestinal tract of animals to accommodate differing methods of digesting carbohydrates. As a result, animals can be classified into four groups as follows. The first group includes animals with a simple stomach, such as humans, pigs, dogs, and cats. The second group is foregut fermenters, which includes cattle, sheep, and goats. These animals have a ruminant stomach in which they can ferment nondigestible carbohydrates. The third group consists of hindgut fermenters such as horses, rabbits, and guinea pigs. These animals rely on fermentation that occurs in the cecum. The final group consists of birds in which various adaptations have occurred to both store and grind various foodstuffs.
Mouth
The mouth is the space extending from the lips or beak to the pharynx, and it is bounded laterally by the cheeks. It is also called the oral cavity, or buccal cavity, and is where food first enters the digestive tract.
The mouth is lined with stratified squamous epithelium, which protects against friction. For further protection, the epithelium of the gums, hard palate, and dorsum of the tongue are slightly keratinized.Lips, cheeks, and gums
The lips and cheeks contain skeletal muscle covered by skin. The orbicularis oris muscle forms the lips. The lips possess long, tactile hair, and regular hair. The median cleft of the upper lip in carnivores and small ruminants is called the philtrum.
Table 17.1. Length and capacity of selected parts of the digestive tract.
| Species | Part of Digestive Tract | Relative Length of Intestines (%) | Average Length (m) | Relative Capacity (%) | Absolute Capacity (L) |
| Horse | Stomach | 8.5 | 17.96 | ||
| Small intestine | 75 | 22.44 | 30.2 | 63.82 | |
| Cecum | 4 | 1.00 | 15.9 | 33.54 | |
| Large intestine | 21 | 6.47 | 38.4 | 81.25 | |
| Pig | Stomach | 29.2 | 8.00 | ||
| Small intestine | 78 | 18.29 | 33.5 | 9.20 | |
| Cecum | 1 | 0.23 | 5.6 | 1.55 | |
| Large intestine | 21 | 4.99 | 31.7 | 8.70 | |
| Sheep and goats | Rumen | 52.9 | 23.40 | ||
| Reticulum | 4.5 | 2.00 | |||
| Omasum | 2.0 | 0.90 | |||
| Abomasum | 7.5 | 3.30 | |||
| Small intestine | 80 | 26.2 | 20.4 | 9.00 | |
| Cecum | 1 | 0.36 | 2.3 | 1.0 | |
| Large intestine | 19 | 6.17 | 10.4 | 4.6 | |
| Cat | Stomach | 69.5 | 0.34 | ||
| Small intestine | 83 | 1.72 | 14.6 | 0.11 | |
| Cecum | |||||
| Large intestine | 17 | 0.35 | 15.9 | 0.12 | |
| Chicken | Small intestine | 79 | 1.08 | ||
| Cecum | 9 | 0.13 | |||
| Large intestine | 5 | .068 | |||
| Dog | Stomach | 62.3 | 4.33 | ||
| Small intestine | 85 | 4.14 | 23.3 | 1.62 | |
| Cecum | 2 | 0.08 | 1.3 | 0.09 | |
| Large intestine | 13 | 0.60 | 13.1 | 0.91 |
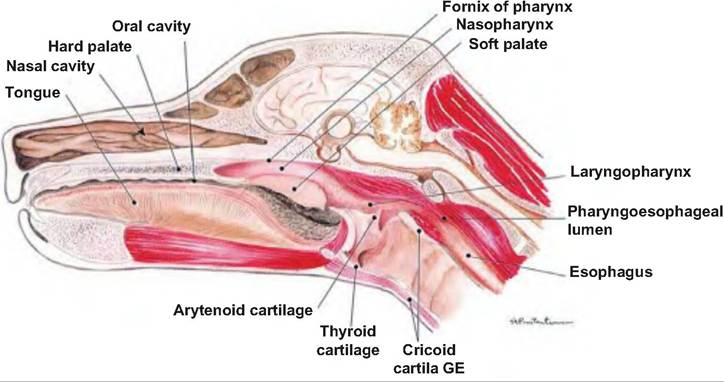
Fig.
17.5. Oral cavity. The hard and soft palate of the mouth is visible in the median section through the head of a dog. (Reprinted from Constantinescu, 2002. Used by permission of the publisher.)The cheeks form the Caudolateral wall of the oral cavity. The gums, or gingivae, enclose the necks of the teeth. The oral cavity is divided into the vestibule and oral cavity proper. The vestibule (porch) is the recess bounded internally by the gums and teeth and externally by the lips and cheeks. The oral cavity proper lies within the teeth and gums.
The palate is the roof of the oral cavity and oropharynx, and it separates the respiratory and digestive passages within the head (Fig. 17.5). It consists of a rostral bony part called the hard palate and a caudal musculomembranous portion called the soft palate. The horse is unable to voluntarily raise its soft palate, and therefore breathes through its nose. The hard palate is formed by the palatine, maxillary, and incisive bones. It forms a hard surface against which the tongue can press food.
The soft palate divides the rostral region of the pharynx into the oral and nasal portions. Projecting downward from the soft palate is the fingerlike uvula. The soft palate closes the nasopharynx as the animal swallows. Birds, unlike mammals, lack a soft palate. The oral and pharyngeal cavities are combined and referred to as the oropharynx. sion, moving food, and making sounds. The extrinsic muscles attach the tongue to bones of the skull and the soft palate. They allow the tongue to protrude, retract, and move side to side. The lingual frenulum attaches the tongue to the floor of the mouth.
The superior surface of the tongue has many papillae that are named for their shape. Filiform papillae are thorn-shaped, giving the tongue roughness and thus aiding in licking and manipulating food. They have a mechanical function. In the ox and cat, they are heavily cornified. Fungiform papillae are mushroomshaped, scattered among the more numerous filiform papillae, have taste buds, and are thus mechanical and gustatory (Fig.
17.6). Foliate papillae have a series of leaf-shaped ridges, are located on the lateral borders of the tongue, and have a gustatory function. They are absent in the ox. Vallate, or circumvallate, papillae are the largest and least numerous. They are located in a V-shaped row near the back of the tongue. They resemble the fungiform papillae, but are circled by a cleft containing taste buds. Marginal taste buds are found along the edge of the rostral portion of the tongue of newborn dogs, but they disappear when puppies switch to solid food.Tongue
The tongue is the muscular organ filling most of the oral cavity. It is composed of interlacing bundles of skeletal muscle fibers, and it is involved in gripping, repositioning food, mixing food with saliva, and forming the compact mass of food called a bolus.
The tongue has intrinsic and extrinsic muscles. The intrinsic muscles, confined to the tongue and not attached to bone, run in several directions allowing the tongue to change shape as necessary for prehenSalivary glands and saliva
Salivary glands are extramural glands (glands outside the wall of the digestive system) that are associated with the oral cavity. The secretions of the salivary glands can be serous, mucous, or mixed. Serous cells produce a watery secretion containing enzymes, ions, and a small amount of mucin, whereas mucous cells produce a viscous, stringy secretion called mucus. Minor salivary glands are located within the wall of the oral cavity and oral pharynx, and have short ducts.
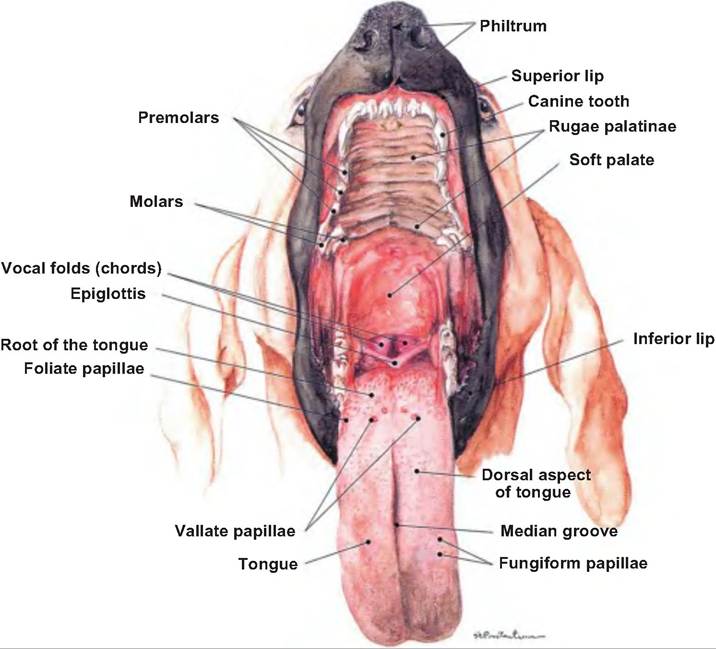
Fig. 17.6. Tongue of the dog. The vallate and fungiform papillae are shown on the tongue of the dog. (Reprinted from Constantinescu, 2002. Used by permission of the publisher.)
They are named for their location (labial, buccal, palatal). They are mixed glands, meaning they have mucous and serous secretions.
The major salivary glands are located some distance from the oral cavity, and require ducts to carry their secretions.
The parotid salivary gland is located below the ear (auricular) cartilage, between the masseter muscle and skin (Fig. 17.7). The parotid duct parallels the zygomatic arch and opens into the buccal vestibule. It produces a predominantly serous secretion. The mandibular (submandibular, submaxillary) salivary gland is located caudal to the angle of the jaw, and is a mixed gland. The mandibular duct runs ros- trally along with the sublingual duct, medial to the mandible, and opens near the sublingual caruncle. The sublingual salivary gland is under the tongue and secretes mostly mucus.Saliva consists of water (97-99.5%), and is therefore hypoosmotic. Electrolytes in the saliva include sodium, potassium, chloride, bicarbonate, and phosphate. It tends to be slightly acidic (pH 6.75-7.00). Saliva has several functions:
1. Solubilizes food. Dissolves foods so they can be tasted and digestive reactions can occur.
2. Provides alkaline buffering and fluid. Bicarbonate and phosphate in the saliva can neutralize acidic feedstuffs. As discussed further, the addition of alkaline fluid via the saliva is particularly important in ruminants.
3. Removes wastes. Metabolic waste products such as urea and uric acid are excreted in the saliva.
4. Lubricates and binds. The mucus in the saliva helps bind masticated food so that it can be formed into a bolus. In addition, saliva coats the oral cavity and esophagus, thus protecting the mucosa of the oral cavity and esophagus.
5. Initiates starch digestion. Salivary amylase begins starch digestion.
6. Assists oral hygiene. Lysozyme, found in saliva, is a bacteriostatic enzyme that lyses bacteria, thus protecting the mouth. IgA attaches to microbes, thus decreasing their ability to penetrate the epithelium. Cyanide, found in saliva, acts as a bacte- riocide, while defensins act as cytokines to attract Iymphoctyes and neutrophils that protect against microbes.
7. Enables evaporative cooling. This is particularly important in dogs, which have very poorly developed sweat glands.
However, it is also used by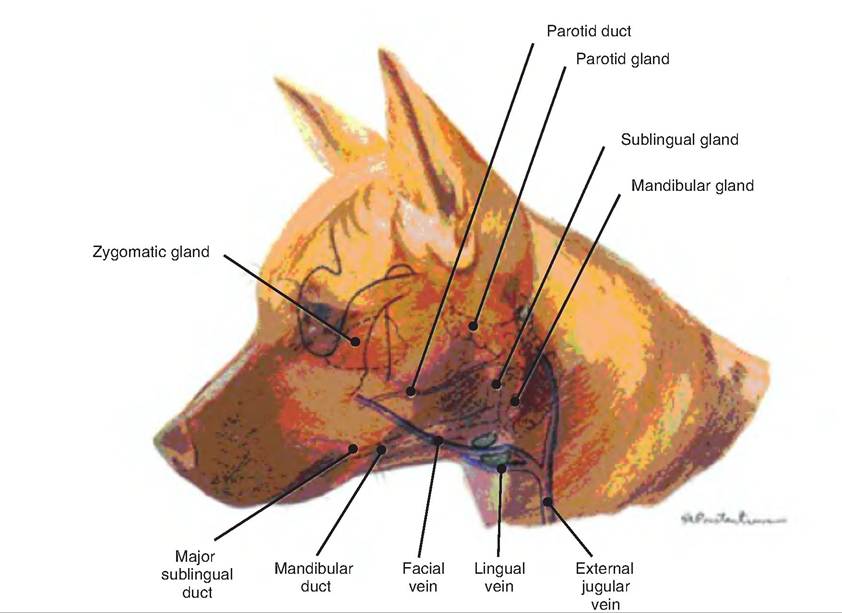
Fig. 17.7. Salivary glands. The major salivary glands are shown in the dog. (Drawings reprinted from Constantinescu, 2002, and Constantinescu and Constantinescu, 2004.)
cats that preen themselves and avian species that display gular flutter (very rapid, but shallow, respiration).
In nonruminants, as the secretion of saliva increases, the concentration of Na+, bicarbonate, and Cb increases, while K+ decreases. In ruminants, as saliva production increases, the levels of Na+ and PO∣ in the saliva decrease, while those of bicarbonate, Ch, and K+ increase.
Salivary glands continuously secrete saliva, thus keeping the oral cavity moist. However, presence of food increases salivation due to parasympathetic nervous system stimulation. Chemoreceptors and mechanoreceptors send signals to the superior and inferior salivatory nuclei in the brain stem. Parasympathetic impulses travel via the facial nerve (cranial nerve VII) and glossopharyngeal nerve (cranial nerve IX) to stimulate salivation.
The sight, smell, sound, or thought of food can also stimulate saliva production. This was evidenced when Pavlov trained dogs to salivate at the sound of a bell. Such salivation helps initiate digestion as soon as food enters the oral cavity.
The saliva in ruminants is isotonic, containing high concentrations of bicarbonate and phosphate, and a high pH. This saliva acts to buffer the acids produced during fermentation in the rumen. An adult cow can produce as much as 100-200 L of saliva daily.
Teeth
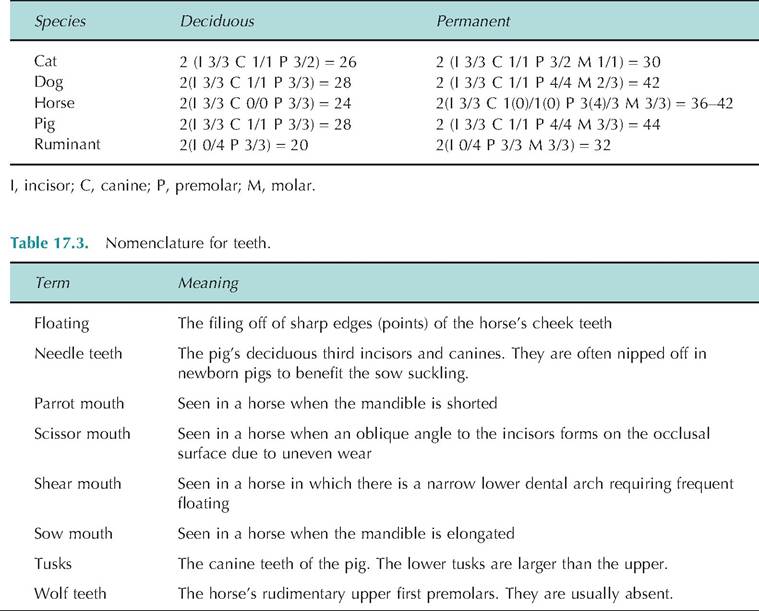
The teeth, or dentes, are accessory digestive organs. They are located in the sockets of the alveolar processes of the mandible and maxillae. Domestic animals have two types of teeth: low-crowned (brachydont) and high-crowned (hypsodont). All domestic species have two sets of teeth, deciduous and permanent. Deciduous teeth are smaller and fewer in number.
Low-crowned teeth are simple teeth, as found in man, carnivores, pigs, ruminant incisors, and horse deciduous incisors. They consist of a crown, neck, and root. The crown is the part projecting above the gum line and is covered with enamel. The neck is the constriction between the crown and root, and it is located at the gum line. The root is the part below the gum line. High-crowned teeth, which have no distinct neck, are found in all permanent horse teeth, ruminant cheek teeth (i.e., premolars and molars), and the tusks of pigs.
Teeth are composed of three layers: cementum, enamel, and dentin (Fig. 17.8). Cementum, a thin, bonelike covering, is found on the entire tooth of
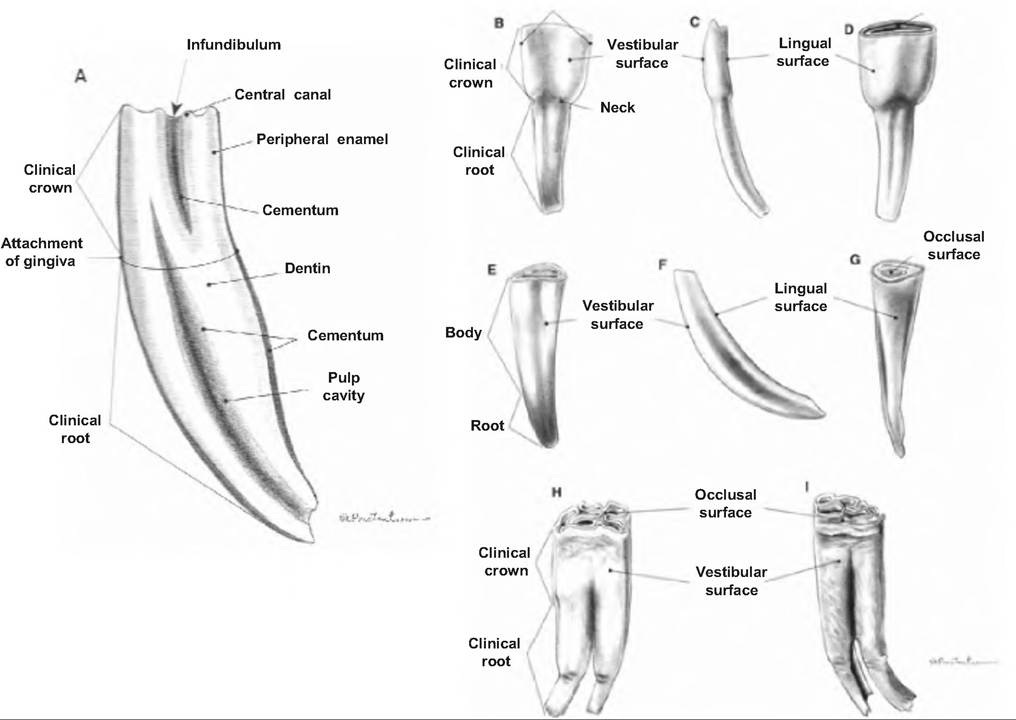
Fig. 17.8. Teeth of the horse. Vertical section of a permanent incisor (A). Vestibular surface (B), left side (C), and lingual surface (D) of the first right-lower deciduous incisor. Vestibular surface (E), left side (F), and lingual surface (G) of first right-lower permanent incisor. Third right-lower permanent premolar (H) and second left-lower permanent molar (I). (Reprinted from Constantinescu and Constantinescu, 2004. Used by permission of the publisher.)
high-crowned teeth, but only on the root of low- crowned teeth. It attaches the root to the periodontal ligament. Enamel, the hardest substance in the body (consisting of 95% calcium salts by dry weight), covers the crown of low-crowned teeth and the body (portion of tooth below crown in high-crowned teeth) and crown of high-crowned teeth. The enamel protects the dentin from acids. Dentin, which makes up the bulk of the tooth, is similar to bone only harder because it has a higher content of calcium salts.
The dentin surrounds a cavity. Within the crown, this cavity is the pulp cavity, and it is filled with pulp, a connective tissue containing blood vessels, nerves, and lymphatic vessels. Narrow extensions of the pulp cavity project into the roots and are called the root canals.
Teeth are divided into groups according to their location and function. Incisors are located in the rostral portion of the mouth (Fig. 17.9). The upper incisors are embedded in the incisive bone and the lower incisors in the incisive part of the mandible. The canine is the large tooth between the incisors and cheek teeth. Cheek teeth are those teeth caudal to the canine and incisors in the maxillary. They include the premolars located in the rostral cheek area and molars located caudal to the premolars.
Cheek teeth function in grinding, while incisors are merely for shearing and biting.
Pigs have 44 teeth; other domestic species have fewer due to a decreased number of cheek teeth (Table 17.2). Ruminants lack upper incisors and canines, which are replaced by a dental pad. They also lack the first upper and lower premolar, giving them 32 permanent teeth. Dogs are missing the upper third molars and therefore have 42 teeth. Brachiocephalic breeds (those dogs that have shortened noses and very prominent eyes due to shallow orbit) may be missing additional teeth. Horses are usually missing the first upper premolar and are always missing the lower first premolar. Mares often have smaller canines that may not
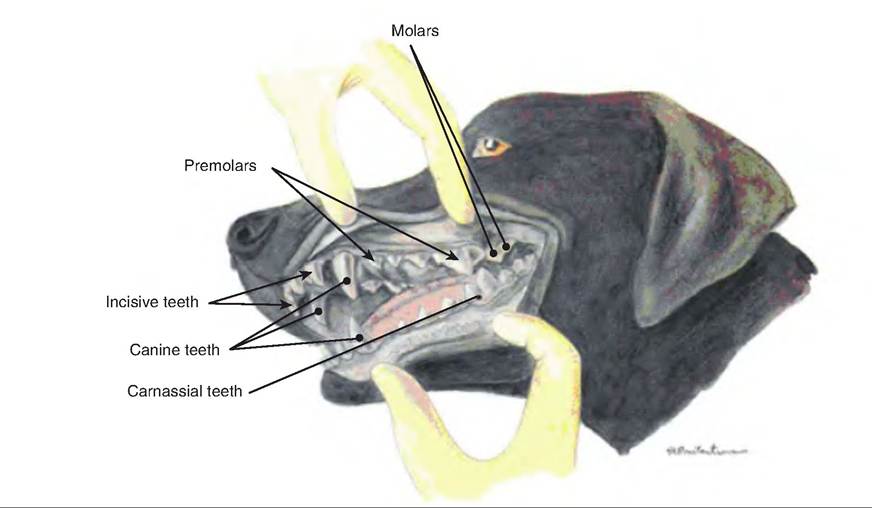
Fig. 17.9. Teeth of the dog. (Reprinted from Constantinescu, 2002. Used by permission of the publisher.)
Table 17.2. Number of teeth in various species.
erupt. As a result, horses may possess 36-42 permanent teeth. Various terms used when discussing teeth are shown in Table 17.3.
The eruption and wear of the lower incisors can be used to estimate a horse's age (Fig. 17.10). If no permanent incisors are present, the horse is probably under 2-1/2 years old. Deciduous teeth are characterized by a distinct neck and are smaller and usually lack longitudinal ridges seen in permanent teeth. The incisors erupt at the following times: II, 2-1/2 years;
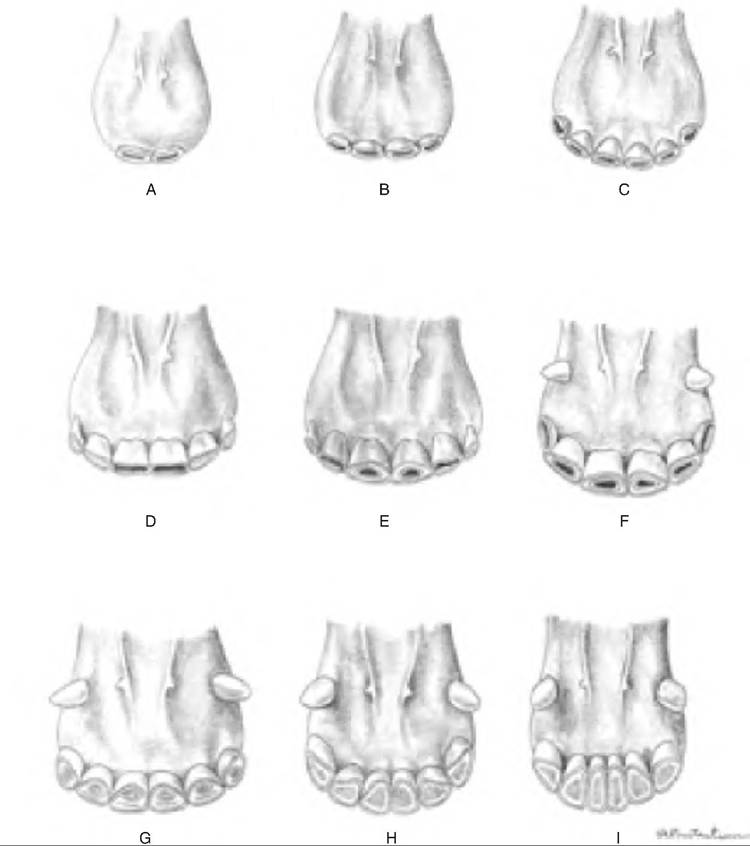
Fig. 17.10. Aging horses using the lower incisors. (A) 4-7 days old. (B) 3-5 weeks old. (C) 7 months old. (D) 3 years old. (E) 4 years old. (F) 5 years old. (G) 10 years old. (H) 15 years old. (I) 20 years old. (Reprinted from Constantinescu and Constantinescu, 2004. Used by permission of the publisher.)
12, 3-1 / 2 years; and 13, 4-1 / 2 years. If all incisors have erupted, and 13 is worn such that a little dentin is seen, the horse is approximately 5 years old. Disappearance of the cup from the respective lower incisors can indicate age as follows: II, 6 years old; 12, 7 years old; and 13, 8 years old. Disappearance of the cup from the upper incisors can indicate age as follows: II, 9 years old; 12, 10 years old; and 13, 11 years old.
Pharynx
The pharynx is the common passageway for food and air. As food first passes from the mouth, it enters the pharynx. It extends from the internal nares to the esophagus. It connects the nasal and oral cavities with the trachea and esophagus, respectively. The soft palate divides the rostral portion of the pharynx into the oropharynx and nasopharynx, and the caudal- most portion of the pharynx is called the Iaryngophar- ynx (Fig. 17.11). The nasopharynx is located dorsal to the soft palate extending from the caudal nares to the Iaryngopharynx with the caudal edge of the soft palate and palatopharyngeal arches separating it from the Iaryngopharynx. The oropharynx lies ventral to the soft palate. The Iaryngopharynx is where air crosses to the larynx and food and water crosses to the esophagus. It is located between the base of the epiglottis and esophageal entrance.
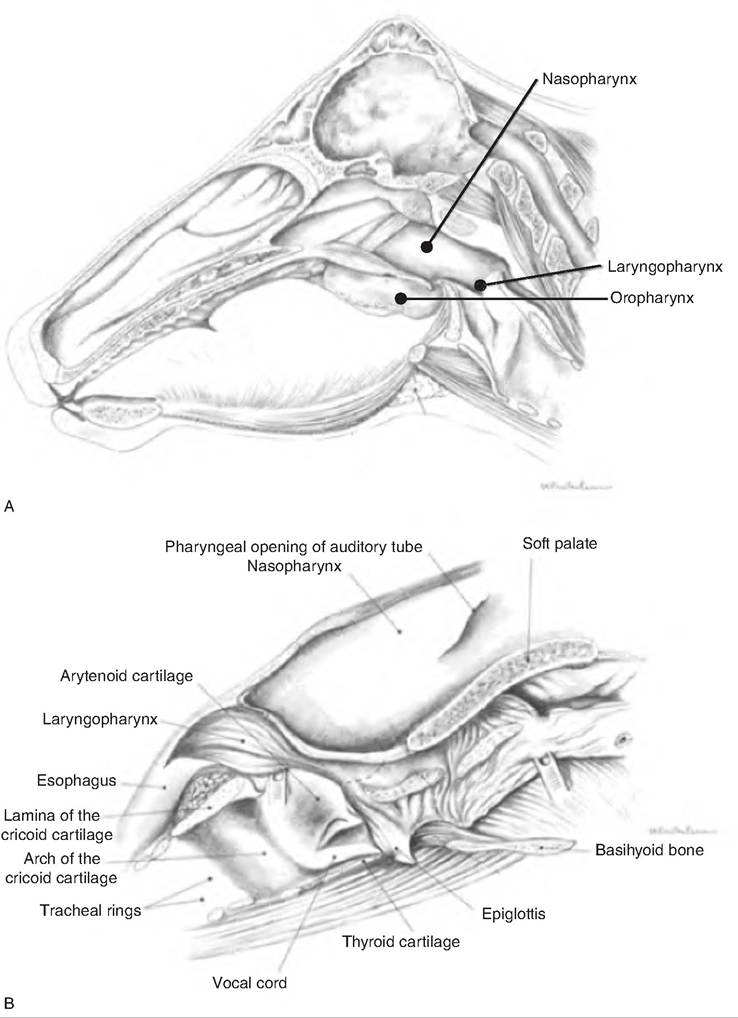
Fig. 17.11. Upper digestive tract of horse. (A) A median section of the head of the horse showing the nasopharynx, oropharynx, and
Iaryngopharynx. (B) An expanded drawing of the pharynx and larynx. (Reprinted from Constantinescu et al., 2004. Used by permission of the publisher.)
The tonsils are an aggregation of lymphatic tissue in the mucosa of the pharynx. They are named for their location (i.e., palatine, pharyngeal, or tubal, which are found around the auditory tube). They help protect the pharyngeal opening against microorganisms and toxic substances. In birds, there is no sharp distinction between the pharynx and mouth.
Swallowing
The act of swallowing, or deglutition, moves food from the mouth, through the pharynx to the esophagus, so that it can be transported to the stomach. Saliva and mucus facilitate this movement. Swallowing involves three stages:
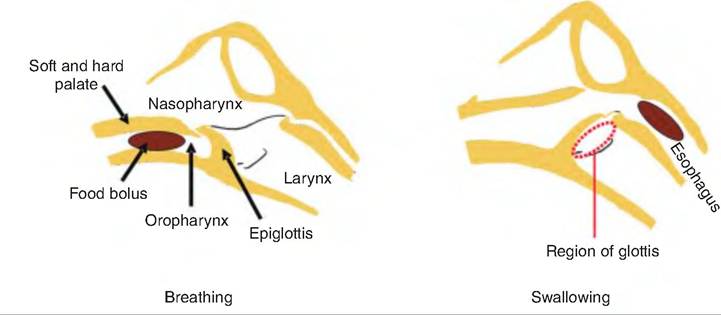
Fig. 17.12. Swallowing. During swallowing, the tongue forces the bolus into the oropharynx, thus raising the soft palate and closing off the nasopharynx. Then, the larynx rises, allowing the epiglottis to cover the glottis, directing the bolus to the esophagus while preventing its entry into the larynx. Once in the esophagus, the bolus is moved to the stomach by peristaltic waves. (Figure modified from Pasquini et al., 1995.)
1. Voluntary stage. Bolus moved into the oropharynx.
2. Pharyngeal stage. Bolus moves involuntarily through the pharynx to the esophagus.
3. Esophageal stage. Bolus moves involuntarily through the esophagus to the stomach.
The tongue, after forming a bolus, propels it from the oral cavity to the oropharynx (Fig. 17.12). This is carried out by skeletal muscle fibers, and is thus the voluntary stage. The presence of a bolus in the oropharynx initiates the pharyngeal stage in which impulses are carried to the deglutition center in the medullar oblongata and lower pons of the brain stem. Motor signals from these centers close off the nasopharynx, and cause the larynx to move forward and upward, allowing the epiglottis to move backward and downward sealing off the rima glottide, the opening in the larynx. After the bolus travels from the Iaryngopharynx to the esophagus, the respiratory passageways reopen.
Esophagus
The esophagus is a collapsible muscular tube lying behind the trachea. It extends from the Iaryngophar- ynx, passes through the mediastinum, pierces the diaphragm at the esophageal hiatus, and ends at the superior portion of the stomach. Ahiatal hernia occurs when a part of the stomach protrudes through the diaphragm at the esophageal hiatus.
In many species of birds, the upper portion of the esophagus is expanded to form the crop. The crop stores food and, in some species (i.e., pigeons), produces a secretion called crop milk that is used to feed the young.
Histology of the esophagus
The esophagus has four layers, as described previously in this chapter. The outermost layer is the adventitia rather than the serosa since the areolar connective tissue is not covered by mesothelium, and the connective tissue merges with structures in the mediastinum, thus attaching the esophagus to surrounding structures. The muscularis externa layer varies in the proportion of skeletal and smooth muscle, depending on the species. The esophagus of birds consists entirely of smooth muscle; that of cats, dogs, pigs, and ruminants consists mostly of smooth muscle, with a small portion of skeletal muscle just as the esophagus nears the stomach.
Stomach
The stomach is located at the inferior end of the esophagus and cranial portion of the abdominal cavity. It is found left of the median plane. It has four functions: (1) storage of ingested food, (2) mechanical breakdown of ingested food, (3) disruption of chemical bonds of food through the action of acids and enzymes, and (4) production of the intrinsic factor required for vitamin B12 absorption from the small intestine. Monogastric animals have a single, simple stomach, and ruminants have a complex stomach consisting of four chambers. The true stomach is the area that produces hydrochloric acid.
Anatomy of the monogastric stomach
In monogastric animals, the stomach appears as a J-shaped structure. Its concave lateral surface is the greater curvature, and the smaller concave medial
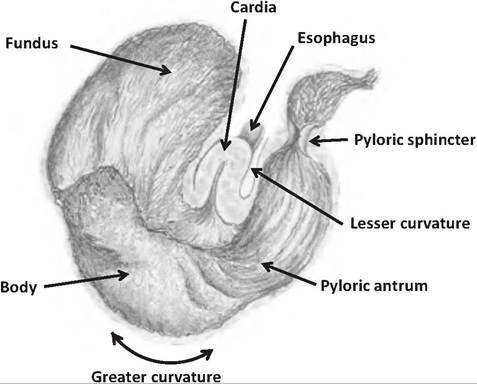
Fig. 17.13. Stomach of the horse. (Adapted from Constantinescu,
G.M. and I.A. Constantinescu, 2004. Clinical Dissection Guidefor Large Animals. Iowa State Press, Ames, Iowa.)
surface is the lesser curvature. The greater and lesser omenta attach to the greater and lesser curvature, respectively.
In addition to the circular and longitudinal smooth muscle layers found along the remainder of the digestive tract, the muscularis externa of the stomach has an additional inner oblique or transverse layer. This extra layer of muscle helps strengthen the stomach wall and assist with mixing the chyme, the partially digested food, with enzymes and acid. As food is ingested, the muscles of the stomach relax to accommodate the increased volume of food. While relaxed, prominent folds called rugae are visible on the stomach mucosa. As the stomach expands, the rugae spread or flatten out.
The stomach is typically divided into four regions (Fig. 17.13):
1. Cardia. The cardia is the smallest region, and is found at the junction between the stomach and esophagus. It is located near the heart, and thus is the "cardia" region. This region contains numerous mucous glands that help protect the esophagus from the acids and enzymes of the stomach.
2. Fundus. The fundus lies superior to the junction between the cardia region, acting as a blind-ended sac.
3. Body. The body, the largest region, is located between the fundus and the pylorus. The body functions as a mixing tank for the stomach, and it is the site where most acid and enzyme secretion occurs.
4. Pyloric region. The pyloric region is the caudal- most portion of the stomach. It consists of the pyloric antrum (antrum = cave) connected to the body. The pyloric antrum narrows to the pyloric canal, which connects to the pylorus. The pylorus is separated from the duodenum by the pyloric sphincter. The pyloric sphincter consists of modified smooth muscle that acts as a valve controlling the flow of chyme exiting the stomach.
The stomach is innervated by fibers from the autonomic nervous system. Sympathetic fibers originate from the thoracic splanchnic nerves and carry signals to the celiac plexus. Parasympathetic fibers are supplied by the vagus nerve (cranial nerve X). The arterial supply comes from branches of the gastric and splenic arteries; the veins are part of the hepatic portal system.
Histology of the monogastric stomach
The stomach is lined with simple columnar epithelium with goblet cells. The many goblet cells produce a layer of mucus protecting the mucosal surface of the stomach. Gastric pits, or shallow depressions, are visible on the mucosal surface. The walls of the gastric pits are formed mainly from goblet cells. Gastric pits open into gastric glands (Fig. 17.14). The gastric glands are composed of different cells depending on the stomach region. The cardia and pyloric regions contain primarily mucus-secreting cells, and the glands in the pyloric antrum produce mucus and hormones including gastrin. In the fundus, the glands produce the majority of the stomach secretions, including acid. There are four cell types in gastric glands.
1. Mucous neck cells. Found in the upper, or neck, region of a gastric gland, they produce a more acidic mucus than goblet cells.
2. Parietal cells. Found in the middle region, they secrete hydrochloric acid (HCl) and intrinsic factor. Intrinsic factor is a glycoprotein necessary for absorption of vitamin B12 in the small intestine. These cells are shaped like pitchforks with three prongs, each covered extensively with microvilli, thus increasing their surface area for secretion. The HCl decreases the pH of the stomach to 1.5- 3.5. This low pH has several functions: (1) It is necessary for the function of pepsin; (2) it provides a harsh environment for bacteria ingested with food; (3) it denatures proteins and inactivates enzymes in food; and (4) it breaks down cell walls of plant material and connective tissue in meat.
3. Chief cells. These cells produce pepsinogen, the inactive form of pepsin, and an enzyme that digests proteins. When pepsinogen is first released, it interacts with HCl and converts it to its active form, pepsin. Once activated, pepsin can convert other molecules of pepsinogen to pepsin. Chief cells also secrete minor amounts of lipases.
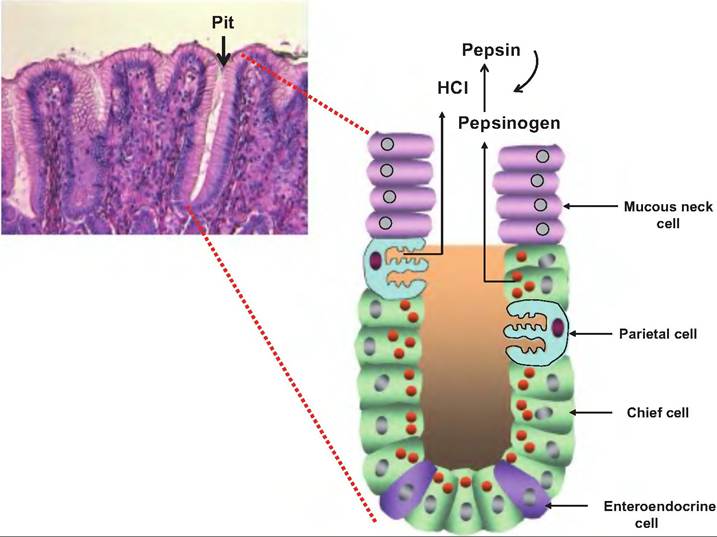
Fig. 17.14. Gastric glands. The histological section of a gastric gland shows the gastric pit, the entrance to the gastric gland. Within the gland are neck cells that secrete mucus, parietal cells that secrete HCI, chief cells that secrete pepsinogen, and enteroendocrine cells that secrete hormones. G cells, which secrete gastrin, are an example of enteroendocrine cells. Once released into the stomach lumen, pepsinogen is converted to pepsin by the action of HCI and pepsin.
4. Enteroendocrine cells. These cells produce a variety of hormones or hormonelike products that are released into the lamina propria. Products include gastrin, histamine, endorphins, serotonin, cholecystokinin (CCK), and somatostatin.
The mucosa of the stomach must provide a substantial barrier against the harsh environment found in the stomach. The concentration of H+ is 10,000 times that found in blood. Pepsin, if not contained, can digest the lining of the stomach. The mucosal barrier of the stomach contains a thick, bicarbonate-rich mucus lining. In addition, the epithelial cells are connected by tight junctions to prevent the leakage of luminal contents to deeper gastric layers. Finally, the epithelial cells are replaced every 3-6 days by division of undifferentiated stem cells found in the gastric pits.
Gastric secretions
Pepsin is not produced within the chief cells because this would cause the self-digestion of the cells. Instead, chief cells produce the zymogen (i.e., the precursor form of an enzyme) pepsinogen, which is activated when it enters the stomach lumen and comes in contact with HCl (Fig. 17.14). Once activated, pepsin can activate other pepsinogen molecules.
Protein digestion is initiated in the stomach via the action of pepsin, the only enzyme found in the stomach of adult animals. Dietary proteins are denatured by HCl secreted by the parietal cells. However, HCl is not produced within the parietal cells because it would destroy the cell. Both H+ and CF are independently transported from the parietal cells into the stomach lumen (Fig. 17.15). Hydrogen ions are generated from the dissociation of carbonic acid that is produced by the enzyme carbonic anhydrase acting upon CO2 and H2O. Hydrogen ions are then transported into the stomach lumen in exchange for K+. Chloride ions enter the parietal cell in exchange for bicarbonate ions. The chloride ions then travel down their concentration gradient and enter the stomach lumen. Once in the lumen, hydrogen and chloride ions combine, producing HCl. Since the pH of the lumen can be as low as 1.5-2.0, this can represent nearly a millionfold (6 log units) increase in hydrogen ion concentration. When parietal cells are producing considerable HCl, a significant amount of bicarbonate enters the blood, thus increasing the pH in what is called the alkaline tide.
Parietal cells respond to many signals. Located on their surface are receptors for histamine, acetylcholine (ACh), and gastrin (Fig. 17.16). Histamine comes from mast cells located in the lamina propria, ACh from postganglionic parasympathetic fibers, and gastrin from G cells. Histamine acts at H2 receptors, whereas ACh acts at muscarinic receptors. Stimulation of these receptors results in stimulation of protein kinase,
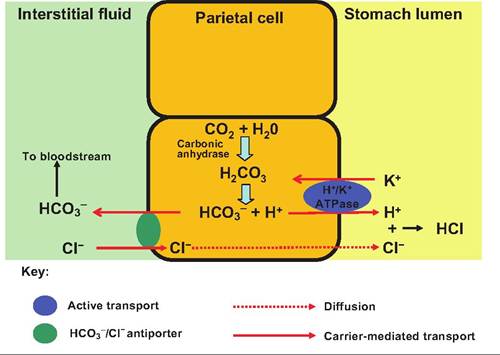
Fig. 17.15. Secretion of HCI by the stomach. Carbonic acid is produced within the parietal cells by the action of carbonic anhydrase. After dissociation, the bicarbonate ions are transported into the intestinal fluid, whereas the hydrogen ion is actively transported into the stomach. Chloride ions enter the cell in exchange for HCO3-, and then move down their concentration gradient and into the stomach lumen where they combine with hydrogen ions forming HCI.
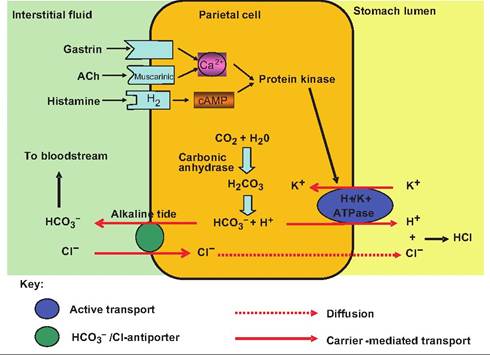
Fig. 17.16. Regulation of HCI secretion from parietal cells. Parietal cells have receptor sites for histamine, gastrin, and acetylcholine (ACh). Histamine binds to H2 receptors, while ACh acts at muscarinic receptors. Stimulation of these receptors by the appropriate ligand initiates a second-messenger system that results in increased HCI secretion into the stomach lumen.
which then stimulates the H+∕K+ ATPase, thus increasing HCl secretion by parietal cells.
Endocrine cells located in pyloric glands produce at least seven hormones. The major hormone, gastrin, is secreted by G cells found most abundantly in the gastric pits of the pyloric antrum. Gastrin stimulates secretion of both parietal and chief cells, and causes contractions of the gastric wall, thus mixing luminal contents. Pyloric glands also contain D cells that secrete somatostatin. Somatostatin, which is released into the interstitial fluid bathing the G cells, inhibits gastrin release. This inhibition can be overridden by other neural and hormonal stimuli such as ACh and histamine.
Gastric motility
With the arrival of food, the stomach can stretch to accommodate this increased volume without an increase in luminal pressure. This reflexive relaxation is mediated by the vagus nerve. In addition, the stomach can actively dilate in a process called adaptive relaxation, which appears mediated by the release of nitric oxide freed by local neurons. In addition to the propulsion of food into the duodenum, the stomach churns and mixes food within its lumen.
Peristalsis in the stomach begins near the cardiac sphincter with gentle ripple-like movements toward the pyloric sphincter. The peristaltic waves strengthen as they move toward the pylorus. The pyloric sphincter, acting sort of like a dam, allows only liquids and small particles to pass over its opening. Heavier particles settle below the level of the sphincter and thus do not pass through. As the peristaltic wave nears the pyloric sphincter, a small amount of chyme is squirted through the sphincter before the peristaltic wave closes the sphincter, causing the remainder of the material to be propelled backward into the pylorus and further churned. Such an action further breaks down the particle size of the ingesta.
The peristaltic rhythm is controlled by the spontaneous activity of pacemaker cells located in the longitudinal smooth muscle layer. These noncontractile cells, called interstitial cells of Cajal, are located near the cardiac sphincter depolarize and repolarize approximately three times per minute producing slow waves, or the basic electrical rhythm. These slow waves migrate throughout the stomach via the gap junctions that electrically couple smooth muscle cells. Slow waves establish the maximum rate of smooth muscle contraction by producing subthreshold depolarizations on which depolarizations resulting in contractions are superimposed (Fig. 17.17).
Vomiting and egestion
Presence of irritants or toxins in the stomach can stimulate vomiting, or emesis. Sensory impulses sent to the emetic center in the medulla oblongata initiate a motor response that causes the diaphragm and abdominal wall muscle to contract, increasing intraabdominal pressure. As the pressure increases, the cardiac sphincter relaxes; the soft palate rises to close off the nasopharynx; and the stomach contents are forced upward through the esophagus, pharynx, and mouth. Excessive vomiting can cause metabolic alkalosis, dehydration, and electrolyte imbalances.
Egestion is a process unique to birds. During egestion, nondigestible materials such as bone, fur, or feathers are orally eliminated from the digestive tract.
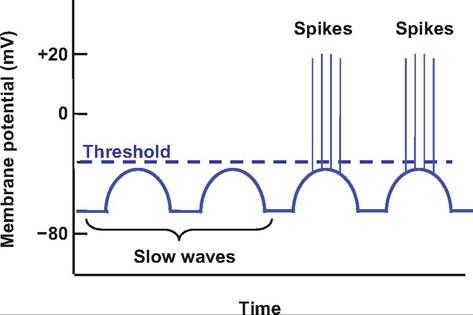
Fig. 17.17. Slow waves. Slow, rhythmic depolarizations and repolarizations, called slow waves or basic electrical rhythm, are initiated by the interstitial cells of Cajal. They establish the background rhythm for peristalsis, which involves spikes superimposed on the slow waves resulting in depolarizations above threshold that cause smooth muscle contractions.
Approximately 12 minutes prior to egestion, gizzard contractions increase, resulting in the compaction of this Undigestible material into a pellet. The pellet can contain exoskeletons of insects and indigestible plant material. Seconds before egestion, the pellet is moved orad by esophageal antiperistalis. This process does not use abdominal or duodenal muscles.
Regulation of gastric secretions and emptying
Gastric secretions are controlled by neural and hormonal mechanisms. The nervous control includes both long and short nerve reflexes involving the vagus nerve. Stimulation of the vagus nerve (i.e., parasympathetic nervous system) increases secretory activity of the stomach. In contrast, sympathetic stimulation inhibits stomach secretion.
Gastric secretions are controlled at three levels, including the central nervous system, stomach, and small intestine. Controls from these three sites are the cephalic phase, gastric phase, and intestinal phase of gastric secretion, respectively (Fig. 17.18). These control mechanisms can either increase or decrease gastric secretions.
Cephalic phase
The cephalic phase causes an increase in gastric secretions prior to the arrival of food. This stage is controlled by the central nervous system, and it prepares the stomach for the arrival of food. The sight, smell, and taste of food stimulate the parasympathetic nervous system to send signals via the vagus nerve that synapse on the submucosal plexus located in the wall of the stomach. This stimulates the postganglionic parasympathetic fibers innervating mucous cells, chief cells, parietal cells, and G cells in the stomach, thus increasing gastric secretions. This phase is short, lasting minutes. Emotional responses associated with activation of the fight-or-flight response decrease gastric secretions and gastric motility.
Gastric phase
Beginning with the arrival of food in the stomach, this phase further stimulates gastric secretion and motility. This phase accounts for about two-thirds of gastric secretions. Stimuli for the gastric phase include distention of the stomach, an increase in gastric pH, and the presence of undigested food, especially proteins and peptides. The arrival of protein in the stomach increases the pH since proteins act as buffers. Activation of stretch receptors sends signals to the myenteric plexus (short loop reflex) and the medulla via the vagus (long loop reflex). These reflexes result in the release of ACh, which stimulates gastric secretions. Chemical stimuli, such as partially digested proteins and increasing pH, also directly activate G cells to secrete gastrin, which strongly stimulates HCl but also has a weaker effect of increasing pepsinogen secretion. A decrease in the luminal pH below 2 inhibits gastrin secretion. Finally, local release of histamine in the lamina propria, presumably from mast cells, also stimulates parietal cells to secrete HCl. Therefore, there are three chemicals that can stimulate parietal cells to release HCl.
Intestinal phase
Involving neural and hormonal signals, the intestinal phase functions to decrease gastric motility. Stimulation of chemoreceptors and stretch receptors triggers the enterogastric reflex. This reflex inhibits gastrin production and gastric motility, and stimulates contraction of the pyloric sphincter, thus slowing gastric emptying into the duodenum. The enterogastric reflex has three components: (1) inhibition of vagal nuclei in the medulla, (2) inhibition of local reflexes, and (3) sympathetic stimulation of the pyloric sphincter causing it to tighten. The enterogastrone reflex is a hormonal reflex. The arrival of lipids (especially medium- and long-chain fatty acids) and amino acids (especially tryptophan) cause the release of CCK and gastric inhibitory peptide (GIP). CCK inhibits gastric secretion of acid and enzymes, while GIP inhibits gastric secretions as well as gastric motility. These reflexes act to prevent the excessive decrease in pH of the small intestine, as well as slow up gut motility in order to facilitate digestion and absorption from the small intestine, particularly in response to lipids. A decrease in duodenal pH below 4.5 also stimulates secretin release by enteroendocrine cells in the duode-
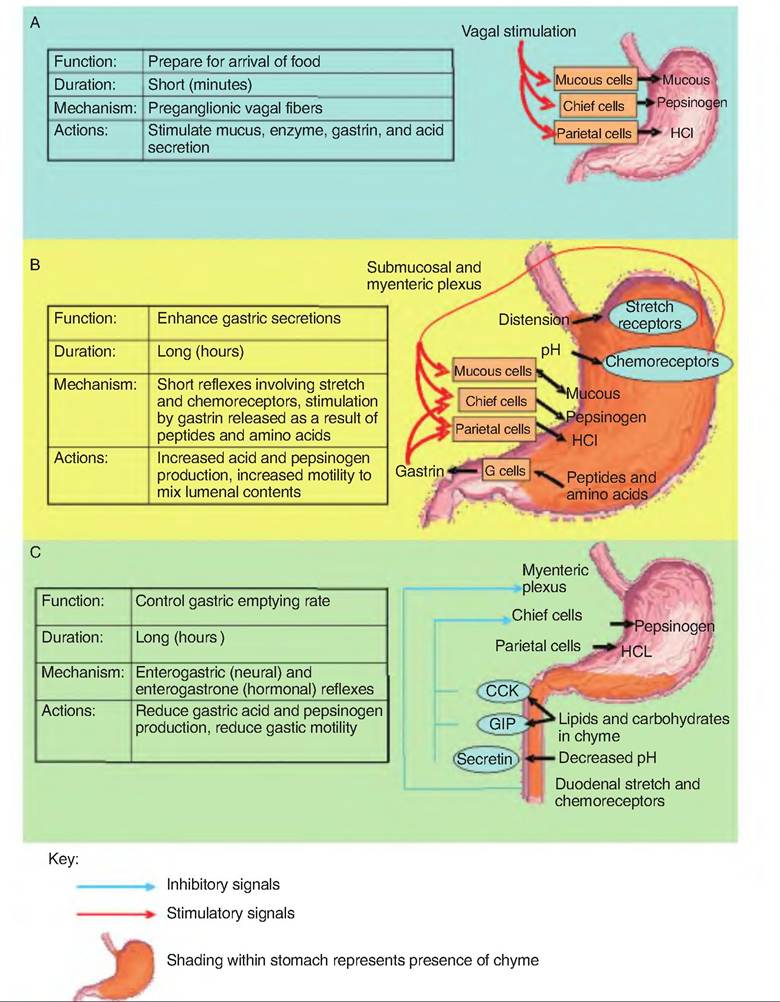
Fig. 17.18. Phases of gastric secretion. (A) Cephalic phase. (B) Gastric phase. (C) Intestinal phase. (Figure modified from Martini, 2004.)
num. Secretin further inhibits gastric HCl and pepsinogen release in the stomach. A summary of these intestinal inhibitory effects on motility is shown in Figure 17.19.
In addition to these inhibitory effects occurring during the intestinal phase, there is an excitatory component. The presence of partially digested proteins in the duodenum stimulates G cells in the duodenal wall to release gastrin that circulates to the stomach to facilitate enzyme secretion. This gastrin is referred to as intestinal (enteric) gastrin. The excitatory phase is short because it is overridden by the inhibitory intestinal phase mechanisms described in the preceding paragraph.
Anatomy of the stomach of ruminants
Ruminants are those animals that ruminate (i.e., chew their cud). They have a specially modified stomach that consists of three nonsecretory forestomachs and a secretory "true" stomach. The forestomachs include the reticulum, rumen, and omasum; the true stomach
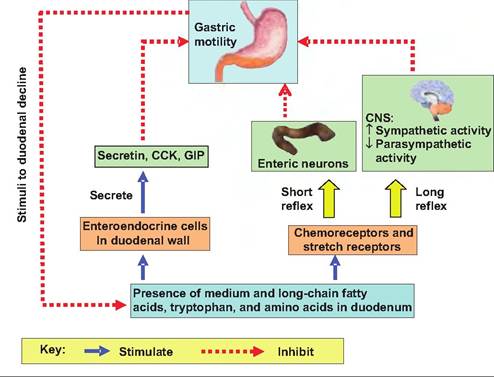
Fig. 17.19. Neural and hormonal inhibition of gastric emptying. Inhibitory signals originating in the duodenum slow gastric emptying allowing the duodenum time to buffer the acid and to absorb nutrients.
is the abomasum (Fig. 17.20). The forestomachs serve as a large fermentation chamber where microbial digestion occurs, allowing the ruminant to digest feedstuffs not available to nonruminants. The fermentation end products, such as volatile fatty acids (VFAs), are absorbed and used as metabolic substrates.
The esophagus connects with the reticulum at the cardiac opening (Fig. 17.20). The reticulum is separated from the heart by only the diaphragm. As a result, any hardware such as nails or wire entering the reticulum can puncture the pleural and pericardial spaces, or the liver (hardware disease). Often a magnet is placed in the reticulum to attract hardware and to prevent its migration to the remainder of the stomach.
The lining of the reticulum has a honeycomb arrangement of ridges (Fig. 17.21). The reticulum is separated from the rumen by the ruminoreticular fold, or groove. While this separates the two chambers, there remains an opening connecting the two. Therefore, the rumen and reticulum act as a functional unit, the reticulorumen, which is lined with keratinized, stratified squamous epithelium.
In young animals, the reticuloomasal fold (Fig. 17.22), sometimes called the reticular groove, prevents food from entering the rumen and instead directs it to the omasum. Since the milk ingested during suckling does not require fermentation, it is shunted directly to the abomasum through the omasal canal. This groove closes as a result of a reflex initiated by stimulation of receptors in the mouth and pharynx. This reflex diminishes with age, thus attenuating the reticular groove.
The rumen, sometimes called the "pouch," occupies almost the entire left side of the abdominal cavity. The rumen is divided into a ventral and dorsal sac by the cranial and caudal pillars as well as by the right and left longitudinal pillars (Fig. 17.23). The dorsal sac is further divided into the cranial sac found between the ruminoreticular fold and cranial pillar, the dorsal sac, and a Caudodorsal blind sac. The luminal surface of the rumen is lined with papillae to increase its surface area.
The omasum is kidney-shaped, and lies to the right of the ruminoreticulum and is located between the rumen-reticulum and abomasum. Its lining consists of many Ieaflike folds, or plies, attached to the greater curvature with their free edges extending into the omasal canal. It is therefore sometimes called the "book stomach" since its interior looks like pages of a book. The leaves have small papillae, thus further increasing the surface area. Food enters the omasum via the reticuloomasal orifice and exits to the abomasum via the Omasoabomasal orifice.
The abomasum consists of two glandular regions equivalent to the fundus and pyloric region of the monogastric stomach. The cardiac region is confined to the area adjacent to the Omasoabomasal orifice. The interior of the abomasum has about 12 rugae (folds) that spiral over the fundus and body, but are absent from the pylorus. A constriction in the pylorus separates this region from the duodenum.
The ruminant stomach provides several advantages compared to the monogastric stomach: (1) It allows animals to use feedstuffs too fibrous for monogastrics; (2) cellulose can be broken down and used by ruminants; (3) it allows the use of nonprotein nitrogen sources (urea and uric acid), which are converted by the rumenal microbes to high-value organic nitrogen compounds; and (4) it provides B complex vitamins due to the action of microbes as long as cobalt is present in the diet. However, there are also disadvantages associated with ruminant digest: (1) Animals must spend a considerable part of each day ruminating (chewing); (2) a large amount of alkaline saliva is necessary; and (3) considerable amounts of volatile acids are released into the environment.
Motility of the ruminant stomach
The mixing, or A, sequence spreads across the reticulorumen in a "Z" pattern. It begins with a double contraction in the reticulum that proceeds across the dorsal rumen to the Caudodorsal area. The contraction then propagates through the ventral region of the rumen. This sequence provides extensive mixing of the rumen contents, which disrupts the layering of luminal contents that would allow gas to collect at the top, with coarse solids floating on the surface and finer particles suspended below. Soil and sand gathers in the ventral region. See Figure 17.24 for further explanation of this mixing sequence.
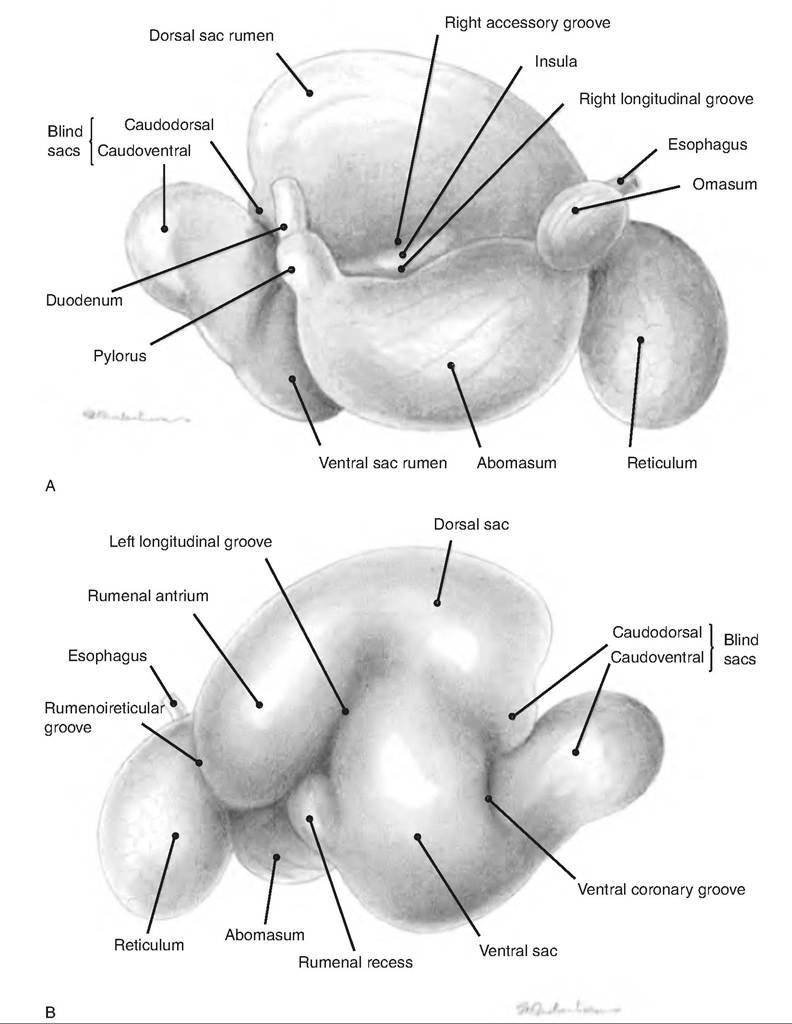
Fig. 17.20. Ruminant stomach. The right (A) and left (B) aspect of the stomach of the goat. (Reprinted from Constantinescu, 2001. Used by permission of the publisher.)
With fermentation comes the production of gas, which must be removed from the animal. The B, or eructation, sequence moves gas from the rumen toward the oral cavity (Fig. 17.24). This sequence allows the formation of a gas bubble, which is eventually forcibly ejected into the esophagus by contraction of the ventral rumen. Excess accumulation of gas in the reticulum and rumen leads to bloat.
While grazing, ruminant animals quickly move feed into the rumen before it is completely masticated. This feed is then returned to the oral cavity through a process called rumination. Rumination is a series of coordinated events involving the respiratory muscles, larynx, pharynx, esophagus, oral cavity, and reticulum. At the height of a single contraction of the reticulum, the animal inhales while the glottis
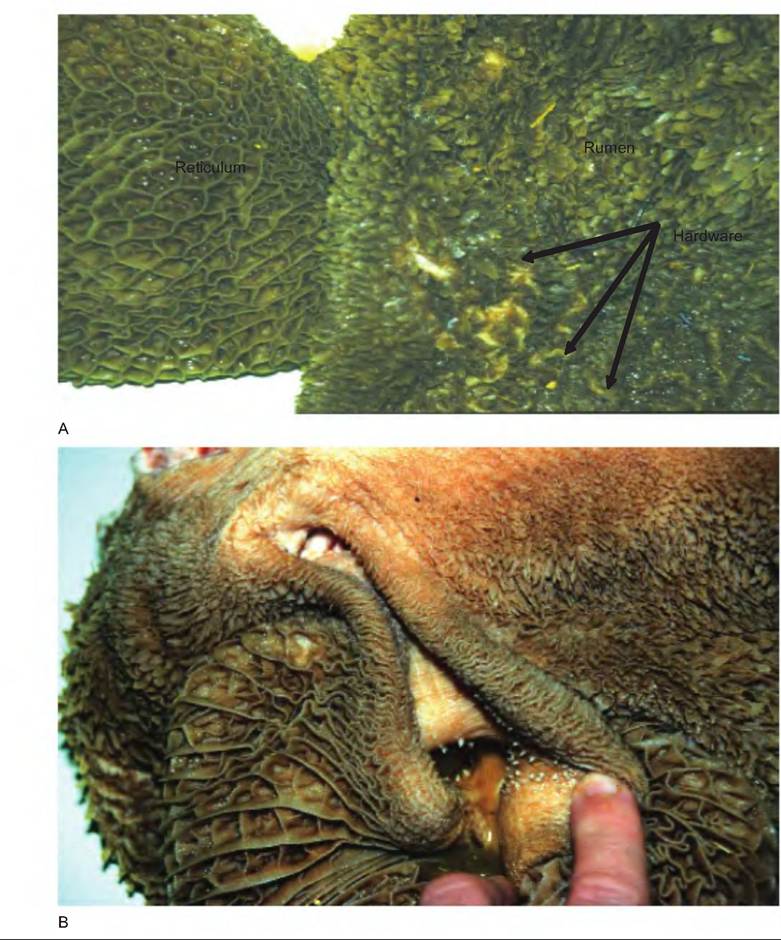
Fig. 17.21. I nside of ruminant stomach. (A) The inside of the reticulum (honeycomb) and rumen. Note the hardware found in the rumen that was ingested as the animal grazed. (B) The reticuloomasal fold (reticular groove). Continued on next page.
is closed so that air cannot flow into the lungs. This generates great negative pressure in the thorax. The transfer of this negative pressure to the esophagus allows a bolus of reticular contents to move through the cardia and, by a process of reverse peristalsis, to move into the oral cavity. Immediately, there is a swallowing event that carries the liquid portion of the bolus back to the rumen. The remaining residue is chewed, saliva is added, and it is again swallowed. Time spent ruminating varies with the diet. A cow consuming a coarse hay diet will spend approximately 8h∕day.
Rumenal microbial fermentation
Fermentation involves the anaerobic action of bacteria and protozoa with bacteria accounting for about 80% of rumen metabolism. Primary bacteria are those that break down the dietary constituents; secondary bacteria further break down the end products of the primary bacteria. Secondary bacteria include those that produce propionate from lactate, and methane-producing bacteria. The protozoa consume rumenal bacteria, plant starch granules, and perhaps linoleic and linolenic acids.

Fig. 17.21. Continued. (C) The many plies shown in the omasum.
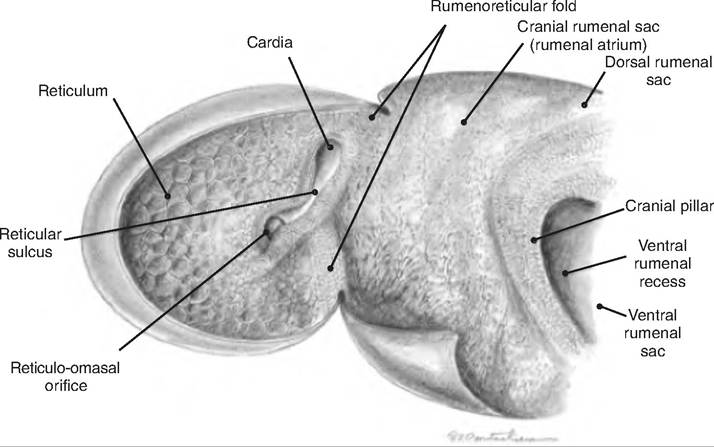
Fig. 17.22. Internal configuration of the reticulum and rumenal atrium of the goat. (Reprinted from Constantinescu, 2001. Used by permission of the publisher.)
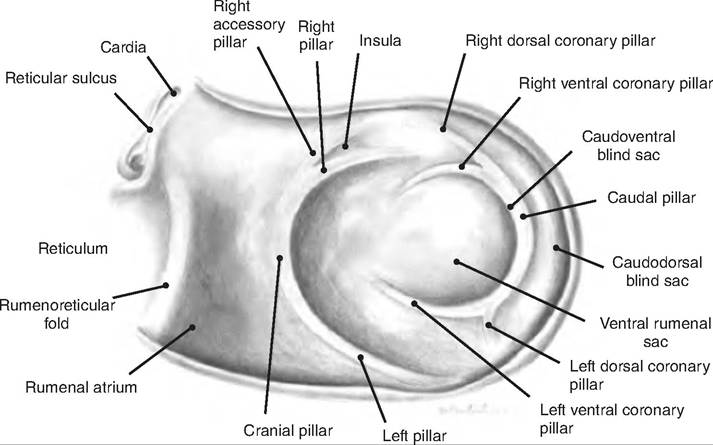
Fig. 17.23. Internal configuration of the rumen of the goat. (Reprinted from Constantinescu, 2001. Used by permission of the publisher.)
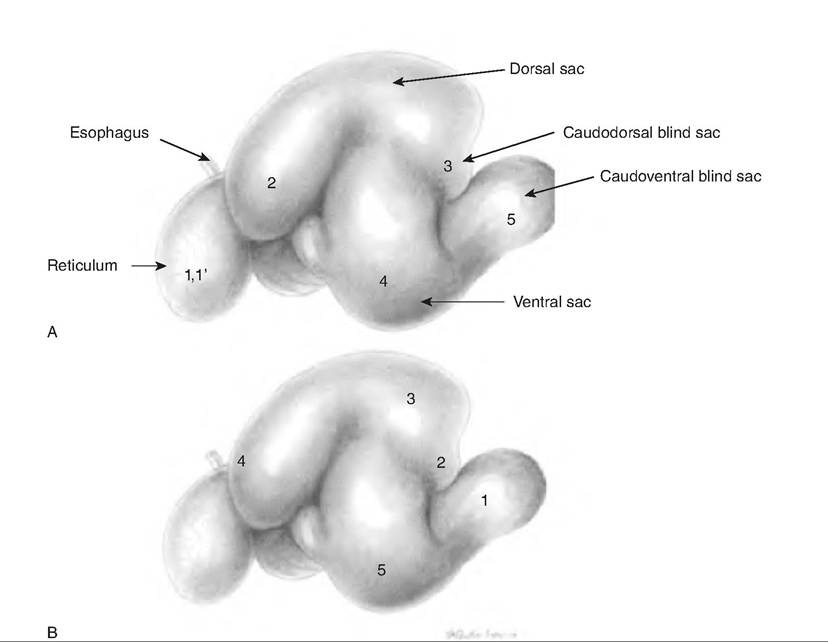
Fig. 17.24. Mixing and eructation sequence of rumen motility. (A) The mixing, or A, sequence, begins in the reticulum in a biphasic, or double, contraction (1,1 ')■ The contraction is next seen in the rumen in the following sequence: anterior dorsal sac (2), caudal region (3), ventral rumen (4), and finally, in the Caudoventral sac (5). (B) The eructation sequence allows elimination of gas produced by fermentation. The sequence begins in the Caudodorsal blind sac (1) and proceeds to the Craniodorsal blind sac (2) and dorsal sac area (3), the cardia (4), and then the ventral rumen (5). (Reprinted from Constantinescu, 2001. Used by permission of the publisher.)
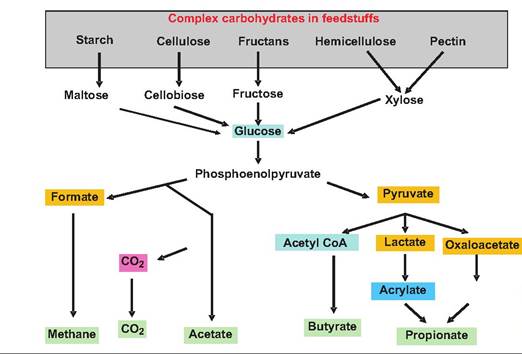
Fig. 17.25. Rumenal carbohydrate fermentation. Complex carbohydrates are fermented by microorganisms within the rumen. Those compounds displayed in blue do not accumulate; those shown in green are the end products.
Rumenal carbohydrate digestion
Products of the bacteria and protozoa carbohydrate digestion include short-chain VFAs, carbon dioxide, and methane (Fig. 17.25). The major VFAs are acetic, propionic, and butyric acids, which are produced in the following proportions: 60-70% acetic acid, 15-20% propionic acid, and 10-15% butyric acid. The percentage of propionic acid increases when the animal is fed concentrates with soluble sugars or starch. The rumen epithelium can absorb glucose and VFAs.
Rumenal protein digestion
Rumen microorganisms hydrolyze dietary proteins to peptides and amino acids. In addition, these microorganisms can make amino acids from nonprotein nitrogen sources such as uric acid, urea, and ammonia. As a result, as much as 50% of the diet of ruminant animals can include poultry waste since the rumenal bacteria will convert the uric acid into amino acids. These amino acids are then absorbed and used by the animal.
Rumenal lipid digestion
Triglycerides are hydrolyzed by rumenal bacteria yielding glycerol and fatty acids. The glycerol is generally metabolized to propionic acid while the fatty acids pass to the duodenum where they are absorbed.
Anatomy of the stomach of birds
In mammals, the stomach is a single chamber. Birds have a two-chambered stomach, including the proventriculus (pars glandularis) and gizzard (pars muscularis) (Fig. 17.26). The proventriculus is the glandular or true stomach and is located orad to the gizzard.
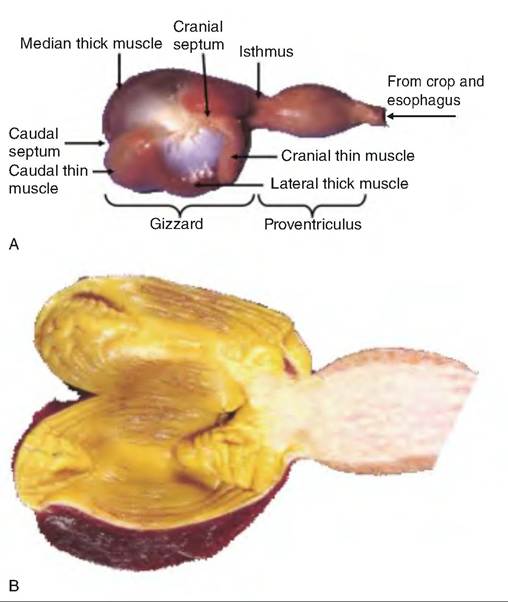
Fig. 17.26. Avian stomach. (A) The avian stomach has two chambers. The gizzard is the most orad and is the muscular, Itonglandular portion. The proventriculus is the glandular portion. (B) The inside aspect of the gizzard and proventriculus.
The interior of the gizzard is lined with a cuticle, occasionally called koilin, produced by the mucosal glands. The cuticle protects the gizzard from acid and proteolytic enzymes secreted from the proventriculus, as well as abrasion from grinding of hard feedstuffs. The gizzard consists of two pairs of opposing circular muscles called thick and thin pairs. Alternating contraction of these muscles results in a grinding motion much like placing a large nut in one's palm and trying to crack it open while squeezing and twisting the hands.
Small intestine
The small intestine is the area where most digestion and 90% of absorption occur. Digestion occurs with the aid of accessory organs that produce necessary enzymes, buffers, and other secretions.
Anatomy of the small intestine
The small intestine extends from the pylorus to the large intestine and is divided into three sections: (1) duodenum, (2) jejunum, and (3) ileum. The duodenum is the first section past the stomach, and is largely a retroperitoneal organ. It is the area where the chyme from the stomach is mixed with the secretions from the pancreas and liver. The duodenum has a descending and ascending segment separated by the caudal flexure (Fig. 17.27 and Fig. 17.28).
The pancreatic and bile ducts empty into the descending duodenum. The bile and pancreatic ducts combine at a point in the wall of the duodenum called the hepatopancreatic ampulla, which opens into the duodenum via the duodenal papilla. The entry of fluid into the duodenum is controlled by the hepatopancreatic sphincter, or sphincter of Oddi (Fig. 17.29).
The longest part of the small intestine, the jejunum runs ventrally and caudally within the abdomen,
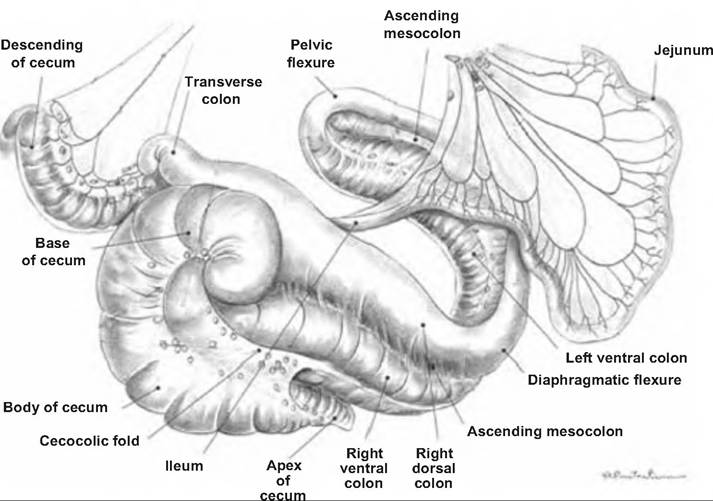
Fig. 17.27. Horse intestinal tract. The small intestine, beginning with the jejunum, and large intestine of the horse. (Reprinted from Constantinescu and Constantinescu, 2004.)
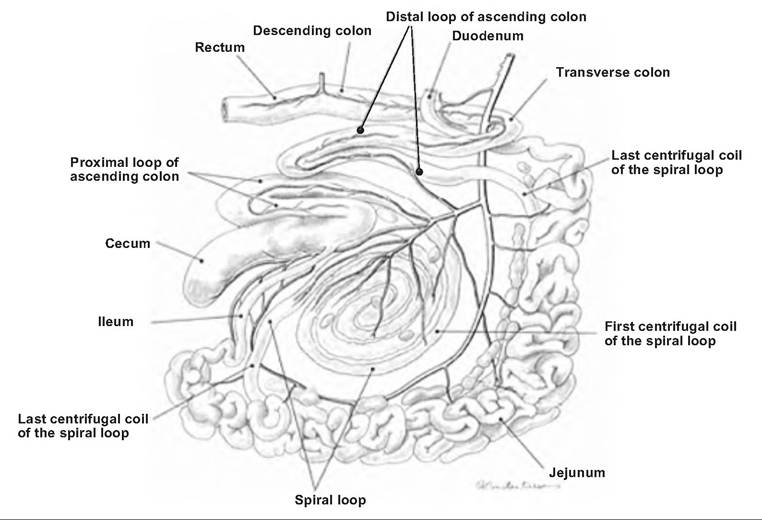
Fig. 17.28. Goat intestinal tract. The large and small intestine of the goat, left aspect. (Reprinted from Constantinescu and Constantinescu, 2004. Used by permission of the publisher.)
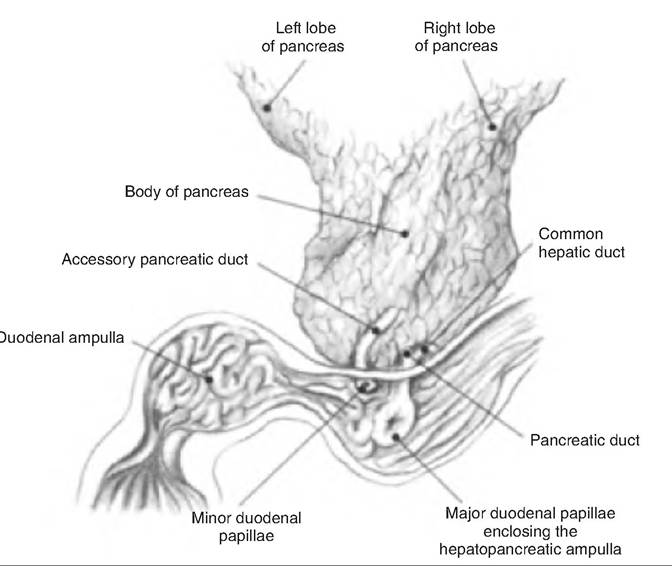
Fig. 17.29. Bile and pancreatic ducts of the horse. The hepatopancreatic sphincter, or sphincter of Oddi, is found surrounding the hepatopancreatic ampulla. (Reprinted from Constantinescu and Constantinescu, 2004. Used by permission of the publisher.)
forming many loops and coils. This is the site of the bulk of chemical digestion and absorption.
The ileum is the short, terminal segment of the small intestine. On gross examination, it is indistinguishable from the jejunum. In birds, the ileum is generally separated from the jejunum at the yolk stalk (diverticulum Vitellinum), formally called MeckeTs diverticulum. The ilium ends at the ileocecal valve, a sphincter controlling the movement of digesta from the ileum into the cecum.
An extensive mesentery attaches the jejunum and ileum to the dorsal abdominal wall. Blood vessels, lymphatics, and nerves reach the small intestine through the mesentery. The blood supply is from the cranial and caudal mesenteric artery (Fig. 17.30). The venous blood from the small intestine drains into the hepatic portal vein that runs to the liver.
Histology of the small intestine
The interior of the small intestine contains transverse folds called plicae, or plicae circulares (Fig. 17.31). Although the rugae of the stomach are transient depending on the stretch in the lumen, the plicae are permanent. They increase the surface area.
The mucosa has fingerlike projections called intestinal villi. The villi are covered with simple columnar epitheliums that have microvilli on their surface. The microvilli make up the brush border. As discussed further, enzymes are embedded in the walls of the plasma membranes of the microvilli. These enzymes are involved in mucosal digestion of carbohydrates and proteins. The combination of plicae, villi, and brush border increases the surface area of the small intestine approximately 600-fold.
The lamina propria within each villi has an extensive network of capillaries originating in the submucosa. These capillaries carry absorbed nutrients to the liver. In addition, there is a lymphatic capillary called a lacteal, or central lacteal, which transports materials not entering the blood capillaries. The Iacteals carry lipid-protein mixtures called chylomicrons to the thoracic ducts where they enter the venous blood supply at the left subclavian vein.
Contraction of smooth muscle located in the muscularis mucosa allows the intestinal villi to move back and forth. Such movement assists in moving the liquefied intestinal contents into contact with the mucosa aiding in digestion and absorption. Such movement also assists with squeezing the lymph along the lacteal and out the villi.
Goblet cells are located between the columnar epithelial cells lining the villi. These cells secrete mucus into the lumen. At the base of each villi are entrances to intestinal glands, or crypts of Lieberkuhn. These glands secrete a watery mucus-containing intestinal juice that assists with absorption of nutrients. The stimulus for secretion is distension or irritation of the
Fig. 17.30.
publisher.)
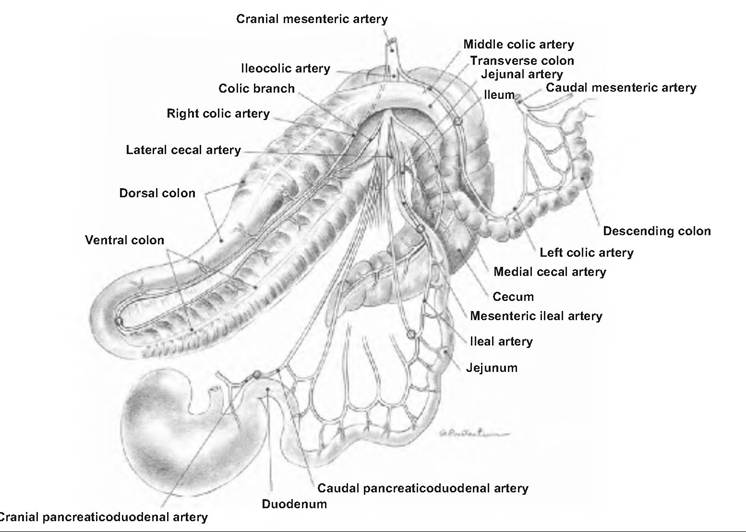
Blood supply to the intestines of the horse. (Reprinted from Constantinescu and Constantinescuz 2004. Used by permission of the
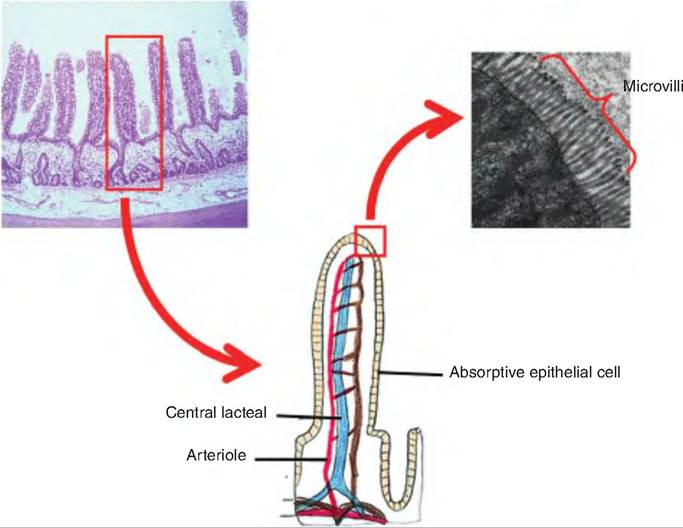
Fig. 17.31. Intestinal wall. Histological section of the wall of the small intestine showing a villi, and then expanded to show microvilli found on the columnar epithelia cells lining the villi.
intestinal mucosa. Located at the base of each gland are stem cells that produce new epithelial cells that migrate up the villi. These cells replace cells sloughed off near the villi tips and constantly replace brushborder enzymes. Also located within the crypts are Paneth cells, which secrete lysozyme and are part of the immune system.
The submucosa consists of areolar connective tissue. It contains aggregates of lymphatic tissue called Peyer's patches that become more numerous toward the end of the small intestine. These function to prevent bacteria from entering the bloodstream. Also in the submucosa are duodenal glands, sometimes called submucosal glands or Brunner's glands. These glands produce large amounts of alkaline mucus to protect the epithelium from the acidic chyme arriving from the stomach.
The duodenum has a bilayered muscularis externa containing circular and longitudinal smooth muscle. Since the bulk of the duodenum is retroperitoneal, it is lined with an adventitia. When located within the peritoneal cavity, the external surface is covered with the peritoneum. The serosal layer of the peritoneum lines the intestinal surface, and the visceral portion lines the peritoneal cavity.
Intestinal juices and brush-border enzymes
Intestinal juices are secreted from the mucosal lining of the small intestine. They contain water and mucus, and are slightly alkaline (pH 7.6). This liquid aids in absorption of substances from the digestive tract lumen. Also embedded in the microvilli of the absorptive epithelial cells lining the small intestine are enzymes called brush-border enzymes. These enzymes include the carbohydrate-digesting enzymes α-dextrinase, maltase, sucrase and lactase, the proteindigesting enzymes aminopeptidase and dipeptidase, and nucleotide-digesting enzymes nucleosidases and phosphatases, as well as enterokinase, the enzyme that activates trypsinogen.
Mechanical digestion and motility in the small intestine
Small intestine motility is regulated mainly by the enteric reflex responding to the presence of materials in the intestinal lumen. There are two types of movements within the small intestine: segmentation and peristalsis. Segmental contractions are a nonpropagating type of movement that results in churning and mixing of the luminal contents with digestive juices (Fig. 17.1). Segmental contractions, the prominent motility pattern in the small intestine, consist of oscillating ringlike contractions separated by relaxed areas containing a bolus of chyme. The constant formation and then relaxation of these contractual rings along the length of the small intestine result in a mixing action, as if kneading dough, thus forcing the chyme to move from a previously relaxed area into a previously contracted area.
The segmental contractions are initiated by intrinsic pacemaker cells located in the longitudinal smooth muscle layer. The pacemaker cells produce a basic electrical rhythm similar to the slow waves discussed in the stomach. Various factors can change the resting potential of this basic electrical rhythm, moving it either closer or farther away from the threshold. Parasympathetic stimulation enhances, and sympathetic stimulation reduces, segmental contractions. The presence of ingesta in the duodenum moves the resting potential toward threshold, thus allowing segmental contractions to increase. Segmental contractions are simultaneously increased in the ileum, even though ingesta is not present. This latter effect is caused by gastrin produced in response to food in the stomach (gastroileal reflex).
Late in the intestinal phase, when most nutrients have been absorbed, segmental contractions are replaced by peristaltic contractions. Peristaltic contractions propel chyme along the length of the digestive tract. Peristalsis in the small intestine is controlled by the migrating myoelectric complex (MMC), which is a type of slow wave characterized by three phases. Phase 1 involves quiescence; Phase 2 has irregularly spaced spike activity superimposed on slow waves; and Phase 3 is characterized by high-amplitude, regular spike activity superimposed on slow waves. The MMC begins near the caudal end of the stomach and pushes contents along a short segment of the small intestine before decaying. MMCs slowly migrate down the small intestine. These complexes strengthen as nutrients and water are absorbed, thereby decreasing the distension of the wall of the small intestine.
The MMC is thought to serve a "housekeeping" role by sweeping residual undigested material through the digestive tract. In dogs and man, the cycle recurs every 1.5-2 hours. The MMC is thought to be controlled from the central nervous system and may be initiated in part by motilin. The growling sound that is occasionally heard from the gastrointestinal tract is caused by the MMC. Ingesting food will switch this pattern to the quieter segmental contractions.
Birds display a unique type of intestinal motility involving intestinal refluxes. This motility involves the reflux of intestinal contents into the proventriculus.
Chemical digestion in the small intestine
Some starch digestion occurs in the mouth by the action of salivary amylase. In the stomach, some chemical protein digestion occurs as pepsin converts proteins to peptides. However, most chemical digestion occurs within the small intestine.
Carbohydrate digestion
Since food remains in the mouth only a short time, little starch is totally digested. Instead, pancreatic amylase completes this process. Starch and glycogen
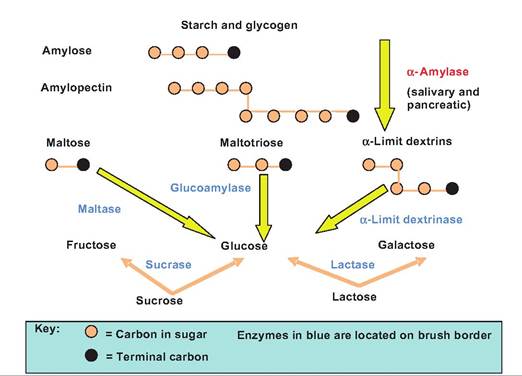
Fig. 17.32. Carbohydrate digestion in the small intestine. Lumenal digestion of carbohydrates results in the production of maltose, maltotriose, and α-dextrins. Mucosal digestion is then completed by the brush-border enzymes. The shaded circles represent the monosaccharides found in the polysaccharides.
are acted upon by salivary and pancreatic amylase to form maltose, maltotriose, and «-dextrins (Fig. 17.32 and Table 17.4). This is the Iumenal phase of carbohydrate digestion since it occurs within the lumen. The smaller di- and trisaccharides then move into contact with the brush border where mucosal digestion by brush-border enzymes digests these sugars to monosaccharides.
Protein digestion
Chemical digestion of protein begins in the stomach by the action of pepsin. Pepsin, which works optimally at a pH of 1.5-2.5, cleaves bonds involving tyrosine and phenylalanine. Pepsin digests approximately 10-15% of dietary protein before being inactivated in the lumen of the small intestine.
Once in the small intestine, trypsin and chymotrypsin secreted by the pancreas break down proteins
Table 17.4. Digestive enzymes.
| Enzyme | Source | Substrate | Products |
| Saliva | |||
| Salivary amylase | Salivary glands | Starch and glycogen | Maltose (disaccharide), maltotriose (trisaccharide), and α-dextrins |
| Lingual lipase | Gland in the tongue | Triglycerides and other lipids | Fatty acids and diglycerides |
| Gastric Secretions | |||
| Pepsin | Chief cells | Proteins | Peptides |
| Gastric lipase | Chief cells | Short-chain triglycerides | Fatty acids and monoglycerides, α-dextrins |
| Pancreatic Secretions | |||
| Trypsin | Pancreatic acinar cells | Proteins, chymotrypsinogen, procarboxypeptidase | Peptides |
| Chymotrypsin | Pancreatic acinar cells | Proteins | Peptides |
| Elastase | Pancreatic acinar cells | Proteins | Peptides |
| Carboxypeptidase | Pancreatic acinar cells | Terminal amino acid at carboxyl end of peptides | Peptides and amino acids |
| Pancreatic lipase | Pancreatic acinar cells | Triglycerides | Fatty acids and monoglycerides |
| Ribonuclease | Pancreatic acinar cells | Ribonucleic acid | Nucleotides |
| Deoxyribonuclease Brush-Border Enzymes | Pancreatic acinar cells | Deoxyribonucleic acid | Nucleotides |
| Cx-Dextrinase | Membrane microvilli | Cx-Dextrins | Glucose |
| Maltase | Membrane microvilli | Maltose | Glucose |
| Sucrase | Membrane microvilli | Sucrose | Glucose and fructose |
| Lactase | Membrane microvilli | Lactose | Glucose and galactose |
| Enterokinase | Membrane microvilli | Trypsinogen | Trypsin |
| Aminopeptidase | Membrane microvilli | Terminal amino acid | Peptides and amino acids |
| Dipeptidase | Membrane microvilli | Dipeptides | Amino acids |
| Nucleosidases | Membrane microvilli | Nucleotides | Nitrogenous bases, pentoses, and phosphates |
| Phosphatases | Membrane microvilli | Phosphorylated nutrient molecules |
into peptides. Carboxypeptidase, another pancreatic enzyme, cleaves one amino acid at a time from the carboxyl and amino end of a polypeptide, respectively, while the brush-border enzymes aminopeptidase and dipeptidase further cleave the proteins.
Lipid digestion
Triglycerides are the most abundant lipid in the diet. Triglycerides and phospholipids are digested by lipases. Although most lipid digestion occurs in the small intestine, lingual and gastric lipases begin the process. Bile salts assist in emulsifying dietary lipids within the aqueous environment found in the small intestine lumen. During this emulsification process, large lipid masses are dispersed into small droplets in which the polar (ionized) portion of the lipids face the outside of the droplet, while the nonpolar (hydrophobic) portions face the inside of the droplet. This increase in surface area produced by reducing the droplet size allows the water-soluble pancreatic lipase to act more efficiently. Pancreatic lipase cleaves off two fatty acids from triglycerides producing two free fatty acids and monoglyceride (Fig. 17.33).
Nucleic acid digestion
DNA and RNA are part of ingested foods. Pancreatic nucleases digest these molecules to their nucleotide monomers. The nucleotides are then acted upon by brush-border nucleosidases and phosphatases that release free bases, pentose sugars, and phosphate ions.
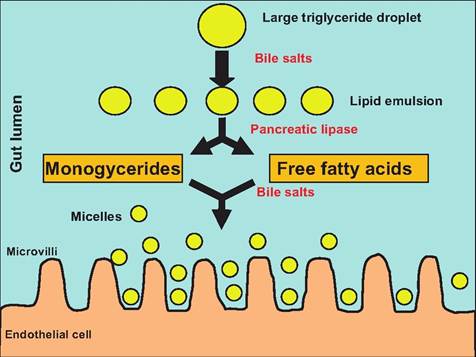
Fig. 17.33. Lipid digestion in the small intestine. Lumenal digestion of lipids results in the production of monoglycerides and free fatty acids. With the help of bile salts, these then form water-soluble micelles that can move toward the microvilli located on the epithelial cells lining the villi of the small intestine.
Absorption in the small intestine
Having decreased the size of the particles through mechanical and chemical digestion, nutrients are now in a form suitable for absorption. Absorption is the process whereby compounds and ions move through the epithelial cells lining the mucosa and pass into the bloodstream or lymphatic system.
About 90% of absorption occurs within the small intestine, with the rest occurring in the stomach and large intestine. Absorption occurs via diffusion, facilitated diffusion, osmosis, and active transport (Fig. 17.34).
Absorption of monosaccharides
The result of Iumenal and mucosal digestion of carbohydrates is the production of monosaccharides, the only form of carbohydrates absorbed. Fructose, a monosaccharide found in fruits, is absorbed by facilitated diffusion, and therefore can only move down its concentration gradient.
Glucose and galactose are absorbed via secondary active transport. These latter two sugars are cotransported across the apical epithelial membrane along with two molecules of Na+. Since Na+ and the sugars are moving in the same direction, this is a symporter. All three binding sites must be occupied for transport
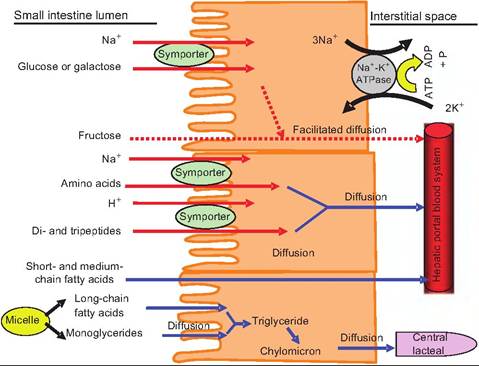
Fig. 17.34. Absorption of nutrients in the small intestine. Carbohydrates, amino acids, and peptides are absorbed by secondary active transport systems. As Na+ is actively pumped out of the epithelial cells and into the interstitial space, this creates a Na+ concentration gradient in which there is a higher concentration in the small intestine lumen than within the epithelial cell. As Na+ enters the epithelial cells moving down its concentration gradient, it carries glucose, galactose, or amino acids with it. Peptides enter epithelial cells via a H+ symporter. Long-chain fatty acids and monoglycerides enter epithelial cells and are re-esterified into triglycerides. These are packaged into chylomicrons and transported into the central Iacteals, and eventually into the subclavian vein.
to occur. While transport across the apical membrane is passive, the driving force for this movement is derived from the Na+-K+-ATPase that actively transports Na+ out of the cell at the basolateral membrane and into the interstitial space. The active transport of Na+ out of the cell produces a lower concentration of Na+ inside the cell than that found in the small intestine lumen. Glucose, galactose, and Na+ can therefore move down their concentration gradient from the intestinal lumen into the epithelial cells. Once inside the cell, the sugars move out of the cell at the basolateral membrane by facilitated diffusion, and into the hepatic portal vein.
Absorption of amino acids, dipeptides, and tripeptides
Although it was once believed that only amino acids are absorbed, it is now well established that di- and tripeptides are also actively absorbed in the small intestine. Some amino acids enter the epithelial cells by a secondary active transport system similar to that described for glucose and galactose. There are some amino acids that utilize a sodium-independent cotransporter in which the amino acids enter along with H+ instead of Na+. In this case, H+ is pumped into the intestinal lumen in exchange for Na+. The Na+ is then pumped out of the cell by the Na+-K+-ATPase on the basolateral membrane. This creates a concentration gradient for H+, which is now in high concentrations within the lumen. As H+ enters the epithelial cells, selected amino acids are cotransported. Peptides are absorbed via this sodium-independent cotransporter. Once inside the epithelial cell, the peptides are hydrolyzed to single amino acids, which then move by diffusion into the hepatic portal vein.
Absorption of lipids
Since lipids are not water soluble, lipid absorption and transport within the body pose unique challenges compared to carbohydrate and protein absorption. Within the small intestine lumen, triglycerides are broken down into fatty acids and monoglycerides. The bile salts within the gut lumen help emulsify the lipids by forming water-soluble particles, which helps the lipids migrate within the aqueous chyme found in the gut. Since lipids are fat soluble; once the micelles come in contact with the lumen wall, the monoglycerides and free fatty acids can cross the epithelial membrane by simple diffusion. The bile salts that helped form the micelles continue to form new micelles down the length of the small intestine. Upon reaching the ileum, the bile salts are reabsorbed via active transport and are recycled. This allows a small amount of bile salts to assist with the absorption of large amounts of lipids.
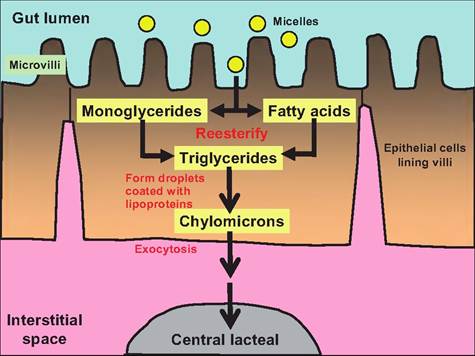
Fig. 17.35. Transport of lipids through the enterocytes. As the micelles produced in the lumen of the small intestine come in contact with the microvilli on the enterocytes lining the villi, the monoglycerides and fatty acids passively pass into the enterocytes. Inside the enterocytes, they re-esterify forming triglycerides. These form droplets that are then coated with lipoproteins, thus forming chylomicrons. The chylomicrons pass through the basal surface of the enterocytes into the interstitial space and then enter the central lacteal found in each villi.
Once inside the epithelial cells lining the gut, the short-chain fatty acids, those having fewer than 12 carbons, pass into the hepatic portal system similarly to amino acids and monosaccharides. The remaining triglycerides and monoglycerides are resynthesized into triglycerides (Fig. 17.35). These triglycerides combine with cholesterol and proteins formed in the rough endoplasmic reticulum to form droplets called chylomicrons. The phospholipids and cholesterol are oriented in the chylomicrons so that their hydrophobic ends face the interior of the droplet and their hydrophilic ends face the surface, thus making these droplets water soluble. The proteins found on the chylomicron surface stabilize the structure.
The chylomicrons are secreted by a process of exo- cytosis into the interstitial fluid of the villus. In mammals, these chylomicrons then enter the central lacteal, which is part of the Iympathic system, and are carried to thoracic ducts where they merge with the venous blood supply at the left subclavian vein. In birds, re-esterified lipids are packaged into portomi- crons. After leaving the enterocytes, portomicrons pass directly into the hepatic portal blood supply and are carried directly to the liver.
Although fat absorption into the epithelial cells is a passive process, it still requires energy. Bile salts are actively secreted by the liver, and the re-esterification of monoglycerides and fatty acids into triglycerides requires energy.
Accessory organs
Pancreas
Chyme passes from the stomach to the small intestine. The chemical digestive processes that occur within the small intestine depend upon accessory organs, including the pancreas, liver, and gallbladder.
The pancreas is a V-shaped retroperitoneal organ lying posterior to the greater curvature of the stomach. It is composed of two lobes joined by a body. The body is the middle portion of the pancreas and is in contact with the pyloric part of the stomach. The right lobe is in the mesoduodenum next to the descending duodenum, and the left lobe lies in the greater omentum. The pancreatic duct (Wirsung's duct) opens with the common bile duct on the major duodenal papilla. There are also accessory pancreatic ducts opening on the minor duodenal papilla. Pigs have only accessory ducts, whereas small ruminants have only pancreatic ducts.
The pancreas has both endocrine and exocrine functions. Its endocrine function includes the synthesis and release of insulin and glucagons, which is discussed in Chapter 12. Its exocrine function is to release enzymes involved in the digestion of all nutrients, including carbohydrates, lipids, proteins, and nucleic acids.
Histology of the pancreas
The pancreas is composed of small clusters of glandular epithelium. Approximately 99% of these cells are arranged in clusters called acini composed of secretory cells surrounding ducts, and are involved in the exocrine portion. These cells have an extensive endoplasmic reticulum and dark-staining zymogen granules. The remaining 1% of the cells are scattered among the acini and are called pancreatic islets (islets of Langerhans) involved in the endocrine function. They secrete glucagons, insulin, somatostatin, and pancre- ateic polypeptide.
Composition and function of pancreatic juice
Pancreatic juice is a clear, colorless liquid. Containing mostly water, it also has salts, sodium bicarbonate, and enzymes. Sodium bicarbonate serves as a buffer to neutralize stomach acid within the small intestine, thus stopping the action of gastric pepsin. Neutralizing gastric acid also allows pancreatic enzymes to function. Pancreatic enzymes include the carbohydrate-digesting enzyme pancreatic amylase; several protein-digesting enzymes, including trypsin, chymotrypsin, carboxypeptidase, and elastase; the triglyceride-digesting enzyme pancreatic lipase; and
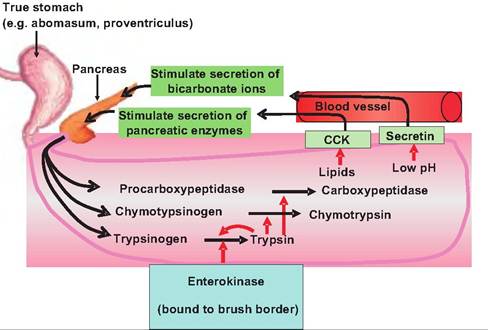
Fig. 17.36. Activation of protein-digesting enzymes. Trypsinogen is secreted into the small intestine lumen where it is converted to trypsin by the enzyme enterokinase found on the brush border. Trypsin then activates the other pancreatic zymogens including chymotrypsinogen and procarboxypeptidase.
the nucleic acid-digesting enzymes ribonuclease and deoxyribonuclease.
These protein-digesting enzymes are produced within the pancreas in an inactive form so that they do not digest the pancreas. Pancreatic acinar cells secrete a protein called trypsin inhibitor that prevents the activation of trypsinogen (Fig. 17.36). Upon entering the duodenal lumen, trypsinogen is acted upon by the brush-border enzyme enterokinase, which splits off a small part of the trypsinogen molecule, thus activating it. Trypsin then activates the remaining zymogens chymotrypsinogen, procarboxypeptidase, and proelastase producing the respective active enzymes.
Regulation of pancreatic secretions
Similar to gastric secretions, pancreatic secretions are controlled by both neural and hormonal mechanisms:
1. During the cephalic and gastric phases of gastric secretion, parasympathetic signals carried via the vagus nerve (cranial nerve X) increase secretion of pancreatic enzymes.
2. Partially digested lipids and proteins within the duodenal lumen stimulate the secretion of CCK from enteroendocrine cells in the duodenal wall. CCK stimulates the secretion of pancreatic enzymes.
3. Decreased pH in the duodenal lumen stimulates the release of secretin from enteroendocrine cells in the duonenal wall. Secretin stimulates release of bicarbonate ions from the pancreas.
Liver and gallbladder
The liver and gallbladder are accessory organs of the digestive system. The liver is the largest gland of the body. It resides just under the diaphragm mostly in the right hypochondriac and epigastric region, although it can extend to the left hypochondriac and umbilical regions (Fig. 17.37). The liver receives blood from the intestines and the general circulation. The gallbladder, which stores bile, is a thin-walled muscular green sac found on the ventral surface of the liver. Water is absorbed, thus concentrating the bile as much as 10-fold. Bile formed in the bile canaliculi moves into the bile ducts, which fuse to form the common hepatic duct.
Deep fissures divide the liver into four lobes. The right lobe is the largest, and is separated from the left lobe by a deep fissure. The quadrate lobe is located between the right and left lobes and ventral to the liver porta, or hilus, the area where blood vessels and nerves enter the organ on the visceral surface. The caudate lobe is located dorsal to the porta.
The liver is covered with visceral peritoneum that closely adheres to its surface. There are also several attachments holding the liver in place. The falciform ligament connects the ventral liver to the sternal part of the diaphragm and ventral abdomen. The round ligament located on the free border of the falciform ligament is a vestige of the umbilical vein. The right and left triangular ligaments attach the liver to the right and left crus of the diaphragm, respectively. In addition, the lesser omentum extends from the liver porta to the lesser curvature of the stomach. The hepatic artery, hepatic portal vein, and other structures enter the liver at the porta, having traveled through the lesser omentum.
Histology of the liver
Each lobe of the liver has approximately 100,000 liver lobules, the functional unit of the liver (Fig. 17.38). Lobules are approximately hexagonal in shape and are separated from each other by an interlobular septum. The cells of the liver, hepatocytes, are arranged in plates that radiate longitudinally outward from the central vein like the spokes of a wheel.
At each corner of the hexagonal lobule is a portal triad consisting of a branch of the hepatic artery, a branch of the hepatic vein, and a bile duct. Instead of capillaries, between the hepatic plates are cavities called sinusoids. They resemble fenestrated capillaries except that they have spaces between adjacent endothelial cells and the basal lamina is thinner or absent. The sinusoids allow even large plasma proteins to
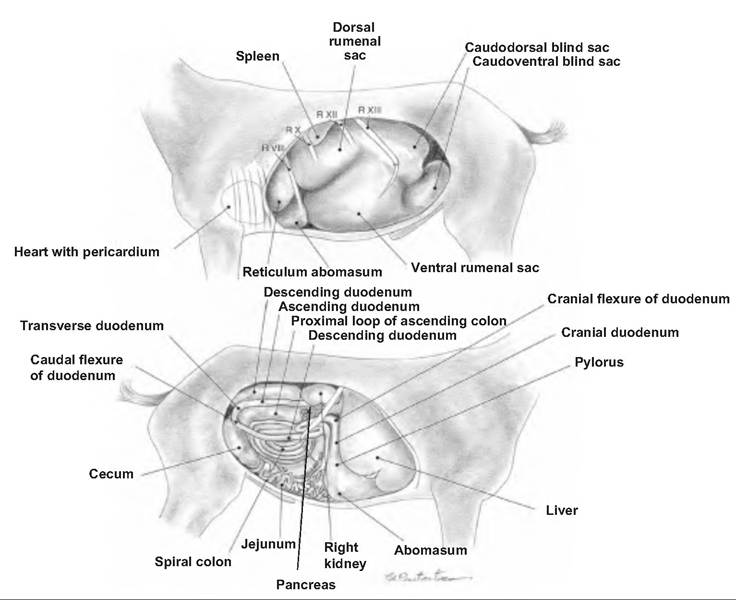
Fig. 17.37. Abdomen of a goat. (A) Right view of the abdominal cavity. (B) Right view of the abdominal visceral. (Reprinted from Constantinescu, 2001. Used by permission of the publisher.)
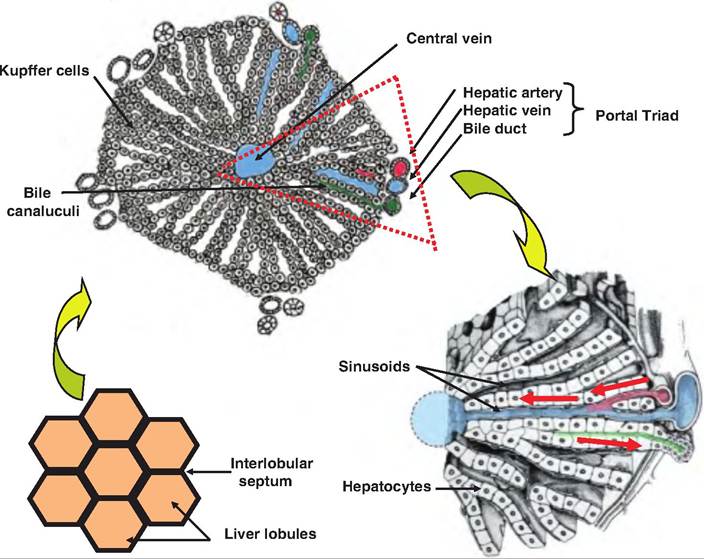
Fig. 17.38. Liver lobules. The functional unit of the liver is the lobule. Lobules are hexagonal in shape and are separated by the interlobular septum as shown in the lower left-hand corner. The portal triad consists of the portal arteriole and venule that bring blood to the liver sinusoids, and the bile duct that collects bile from the bile canaliculi. Liver macrophages, called Kupffer cells, are also found in the sinusoids. Blood from the portal artery and portal vein flow into the sinusoids and toward the central vein. Material filters through the hepatic plates into the bile canaliculi and then flows into the bile ducts. (Adapted from http://curesure.com/lmage_3.jpg.)
pass out of the bloodstream and into the spaces surrounding the hepatocytes. In addition to the endothelial cells lining the sinusoids, there are also star-shaped fixed macrophages called Kupffer cells that are confined to the liver. They phagocytize pathogens, cell debris, and damage red and white blood cells.
Bile is secreted by hepatocytes and enters the bile canaliculi, which are narrow intercellular canals between the hepatocytes. The bile empties into the bile ducts located at the periphery of the lobules. Bile ducts merge to form the right and left hepatic duct, which then unite to form the common hepatic duct that travels toward the duodenum. Before entering the duodenum, the cystic duct from the gallbladder fuses with the common hepatic duct to form the common bile duct. Note that the horse does not have a gallbladder.
Blood supply of the liver
As stated earlier, the liver receives oxygenated blood from the hepatic artery and also receives deoxygenated blood from the hepatic portal vein that contains nutrients, drugs, and toxins from the digestive tract. Branches from both of these arteries enter the liver sinusoids where the hepatocytes remove some nutrients and toxins. As the blood passes through the sinusoids, metabolites from the hepatocytes are secreted into the blood. This blood then passes to the central vein and then into the hepatic vein (Fig. 17.39).
Histology of the gallbladder
The mucosa of the gallbladder is lined with simple columnar epithelium. The surface has rugae similar to those found in the stomach. The middle layer of the gallbladder consists of smooth muscle that allows for expulsion of the bile into the small intestine. The outer layer of the gallbladder is the visceral peritoneum.
Bile composition and function
Bile consists of water, bile salts, bile acids, cholesterol, the phospholipid lecithin, bile pigments, and ions. The bile salts include sodium and potassium salts of bile acids, mostly cholic acids and Chenodeoxycholic acid. These salts assist in emulsification of lipids within the small intestine lumen. This decreases the surface area to volume ratio of the lipids, thus increasing the efficiency of pancreatic lipase as it digests triglycerides. Bile salts, free fatty acids, and lecithin combine to form micelles within the small intestine lumen. The polar ends of the molecules are arranged to the outside of the micelle, and the nonpolar portions face the inside of the micelle. Cholesterol and fat-soluble vitamins are packaged inside the micelle.
As aged red blood cells are phagocytized, iron, globin, and bilirubin are liberated. Bilirubin is the main waste product of heme breakdown. The globin and iron are recycled, but bilirubin is absorbed from the blood by the hepatocytes and excreted into the bile conjugated to glucuronic acid. When in the small intestine, it is metabolized by bacteria. Stercobilin, one of its breakdown products, gives feces their dark brown color. Most bile salts are reabsorbed via active transport in the ileum whereupon they enter the hepatic portal blood and return to the liver.
Regulation of bile secretion
In those animals that have a gallbladder, bile is stored between meals. The sphincter of the hepatopancreatic ampulla (sphincter of Oddi) restricts the entrance to
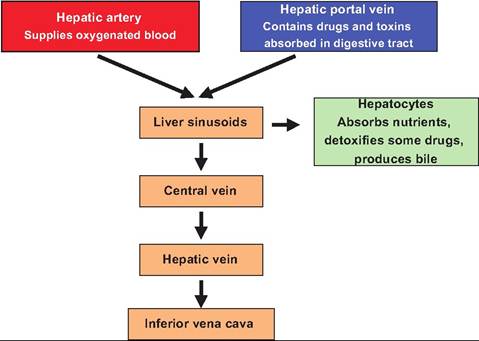
Fig. 17.39. Blood flow through the liver lobule.
the duodenum. There are neural and hormonal stimuli that can stimulate bile secretion:
1. Parasympathetic signals traveling along the vagus nerve can stimulate bile production by the liver.
2. Fatty acids, particularly medium- and long-chain, and amino acids in the chyme cause duodenal enteroendocrine cells to secrete CCK. CCK causes the smooth muscle cells of the gallbladder to contract, and bile is squeezed into the cystic duct and through the common bile duct. CCK also relaxes the sphincter of the hepatopancreatic ampulla. The action of hormones controlling digestion is summarized in Table 17.5.
Functions of the liver
The liver is a vital organ. It performs many functions:
1. Carbohydrate metabolism. The liver plays a vital role in maintaining blood glucose levels. When blood glucose levels are high, the liver converts glucose to glycogen (glycogenesis) and triglycerides so that energy can be stored until needed. When blood glucose levels drop, the liver can break down glycogen to glucose (glycogenolysis) and release the glucose into the bloodstream. In addition, the liver can convert certain amino acids to glucose (gluconeogenesis), as well as lactic acid to glucose.
2. Lipid metabolism. Hepatocytes can store triglycerides as well as use fatty acids to synthesize ATP. In addition, hepatocytes synthesize lipoproteins that carry fatty acids, triglycerides, and cholesterol throughout the body. Cholesterol can be synthesized in the liver, and cholesterol is used to make bile salts.
3. Protein metabolism. Hepatocytes remove the amino group (deamination) of amino acids so they can be used for ATP synthesis. Hepatocytes also synthesize carbohydrates and fats from certain amino acids. Hepatocytes can synthesize various plasma proteins such as albumin, prothrombin, fibrinogen, and alpha and beta globulins (needed for hemoglobin synthesis).
Table 17.5. Hormones controlling digestion.
| Hormone | Site of Secretion | Stimulus for Secretion | Actions |
| Gastrin | Enteroendocrine G cells in the pyloric antrum mucosa, also secreted from duodenal mucosa | Stomach distention, partially digested protein, high pH in the stomach | Increases secretion of gastric juice, increases gastric motility, stimulates growth of gastric mucosa, constricts lower esophageal sphincter while relaxing ileocecal sphincter |
| Secretin | Enteroendocrine S cells in duodenal mucosa | Acidic pH in duodenal lumen | Stimulates pancreatic secretion rich in bicarbonate ions |
| Cholecystokinin (CCK) | Enteroendocrine CCK cells in duodenal mucosa | Partially digested proteins, medium and long-chain fatty acids | Stimulates pancreatic secretion rich in enzymes, gall bladder bile ejection, opens hepatopancreatic ampulla, inhibits food intake peripherally and in the central nervous system |
4. Removal of waste products. The liver detoxifies substances such as alcohol and antibiotics, and can alter and excrete steroid hormones. The liver is an important site of detoxification of ammonia, which is converted to the less toxic urea, which is excreted in the urine. The waste product of red blood cell destruction, bilirubin is eliminated via the bile.
5. Synthesis of bile salts. The bile salts necessary for lipid emulsification within the small intestine are synthesized in the liver.
6. Storage. The liver is the primary storage site of fat-soluble vitamins (A, D, E, and K), as well as vitamin B12. Glycogen and certain minerals (Fe and Cu) are also stored in the liver.
7. Phagocytosis. Kupffer cells destroy aged blood cells and microbes that may have entered via the hepatic portal blood.
8. Activation of vitamin D. The liver combines with the skin and kidneys to synthesize the active form of vitamin D.
Large intestine
The terminal portion of the digestive tract is the large intestine. It extends from the end of the ilium to the anus. The primary functions of the large intestine are electrolyte and water absorption, microbial digestion and vitamin production, formation of feces, and expulsion of feces. These functions require considerable time, so transit time through the large intestine is slow.
Anatomy of the large intestine
The large intestine is divided into the cecum, colon, rectum, and anal canal. The ileum attaches to the large intestine at the ileocecal valve (sphincter), which controls the movement of chyme into the large intestine.
The cecum is a blind diverticulum that extends off of the colon at the ileocecal valve (Fig. 17.40). It is located on the right side of the abdomen, except in the pig, where it is located on the left. In those animals that are hindgut fermenters (e.g., the horse), the cecum is a major site of digestion. Extending from the cecum is another twisted, coiled blind pouch called the appendix, or vermiform appendix.
The colon consists of three segments called the ascending, transverse, and descending colon. The ascending colon ascends on the right side of the abdomen until it reaches the inferior surface of the liver. It then turns sharply to the left at the right colic (hepatic) flexure, continuing as the transverse colon. Upon reaching the left side of the abdomen near the spleen, it again abruptly bends at the left colic (splenic) flexure and passes inferiorly as the descending colon to the level of the iliac crest. From this point, the sigmoid colon projects medially to the midline, terminating as the rectum. The rectum extends from the large intestine to the anal canal. The digestive tract terminates at the anal canal, which has an exterior opening called the anus.
Histology of the large intestine
Unlike the small intestine, the mucosal lining of the large intestine lacks villi and plicae (circular folds). Since almost no chemical digestion occurs in the large intestine, the lining also lacks cells that secrete digestive enzymes. The colon is lined with simple columnar epithelium and mucus-secreting goblet cells. The anal canal is lined with stratified squamous epithelium that merges with the skin surrounding the anus.
Unique to the large intestine, portions of the longitudinal smooth muscle found in the muscularis externa layer are thicker, forming three visible longitudinal bands called the teniae coli. The areas between the teniae coli are thinner or lack longitudinal muscle. Tonic contractions of the teniae coli result in a series of pouch-like expansions along the length of the colon called haustra (sing. = haustrum). The outermost layer of the large intestine is the visceral peritoneum. Small fat-filled pouches of visceral peritoneum called epiploic appendages are attached to the teniae coli.
Mechanical digestion in the large intestine
The ileocecal valve controls the movement of chyme into the large intestine. Normally closed, the gas- Iroileal reflex initiated immediately after a meal opens this valve so that peristalsis allows chyme to move from the ileum into the large intestine. Gastrin also relaxes this valve.
Haustra churning is characterized by the slow filling of a haustrum until it is distended, at which time its walls contract, squeezing the contents into the next haustrum. Peristalsis also occurs along the large intestine, although at a slower rate than in the small intestine. Finally, mass peristalis occurs when a strong peristaltic wave begins near the middle of the transverse colon and quickly forces the contents to the rectum. Such movement normally occurs near the beginning or end of a meal.
Chemical digestion in the large intestine
Although no chemical digestion occurs in the large intestine, considerable fermentation takes place,
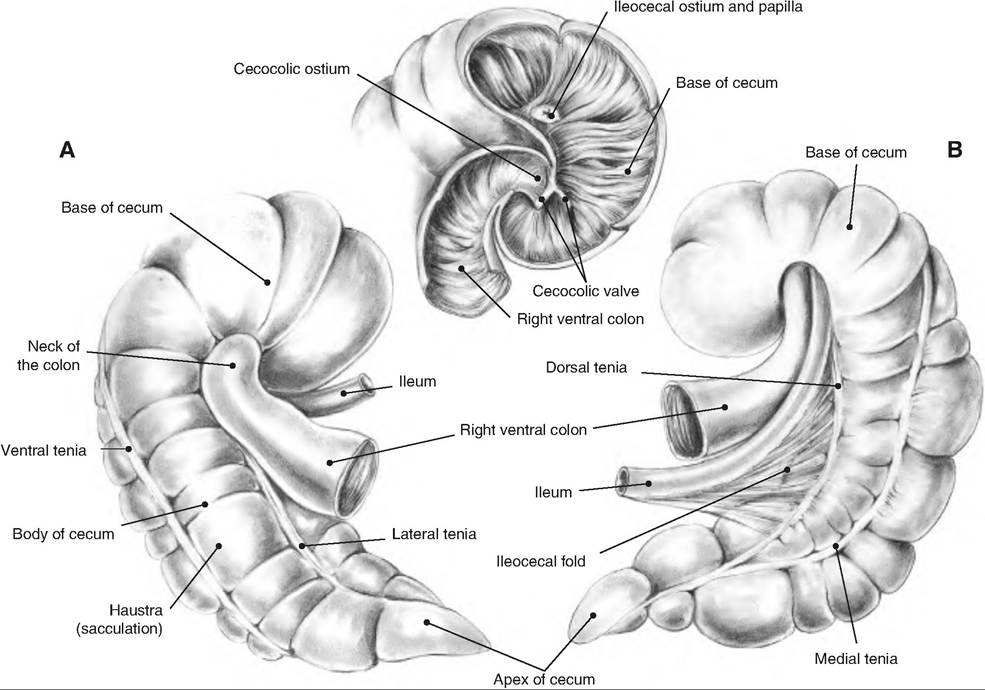
Fig. 17.40. Cecum of the horse. (A) Right aspect. (B) Left aspect. (C) Communications of the cecum. (Reprinted from Constantinescu and Constantinescu, 2004. Used by permission of the publisher.)
especially in those animals that are hindgut fermenters. Microbes ferment any remaining carbohydrates releasing hydrogen, carbon dioxide, and methane. These gases lead to flatulence in the large intestine. Amino acids are converted to indole, skatole, hydrogen sulfide, and fatty acids. Indoles and skatole are eliminated in the feces, contributing to its odor, while some of the remaining products are absorbed and transported to the liver. The microbes synthesize B vitamins and vitamin K, which are also absorbed by the large intestine.
Gut microflora
Gut microflora is more complex than that of its host, and it is now known that there are about 150 times more microbial genes (3.3 million vs. 20,000-25,000) in the human gut than in the human genome. While it is well documented that some gut microflora can cause disease (i.e., Escherichia coli, clostridium, and salmonella), other microflora are necessary. It has been known since the 1980s that the enzymes that make vitamin B12 are produced by bacteria. Bacteroides tlιe- taiotaomicron can break down complex carbohydrates found in plants. Studies show that mice raised in a sterile environment and fed a complex polysaccharide diet, those colonized with B. Bietaiotaomicron, can derive more energy from their diet than those devoid of bacteria.
Recent evidence suggests that other microorganisms, in fact, regulate an animal's genes and immune system (Lee and Mazmanian, 2010). It is now believed that commensal microbiota may "program" aspects of T-cell differentiation, thereby influencing the host genome and the function of the adaptive immune system. Again, using mice raised in a sterile environment, the lack of intestinal microbiota results in defective MALT, fewer Peyer patches, less lamina propria in the small intestine, and less mesenteric lymph nodes There are also extraintestinal changes including reduced numbers of CD4+ T cells in the spleen, and fewer and smaller germinal centers within the spleen. In addition, there are decreased systemic antibody levels. All these changes suggest that the microbiota is capable of shaping systemic immunity.
While Helicobacter pylori has been shown to cause peptic ulcers, it has also been shown to regulate stomach acid levels. If the stomach secretes too much acid, strain of H. pylori containing the gene cagA produce proteins that reduce acid secretion. H. pylori might also affect food intake. When present, these bacteria cause a postprandial reduction in ghrelin, a hormone secreted from the stomach, and induce increased food intake in mammals.
Another example of how the microbiota may interact with an animal's genome is revealed by recent studies on Bacteroides fragilis. Normally, regulatory T cells produce proteins that restrain proinflammatory T cells, thus restricting inflammation and preventing the immune system and preventing the body from attacking itself. B. fragilis have sugar molecules called polysaccharide A (PSA) that protrude from their surface. PSA induces the immune system to produce more regulatory T cells, which reduces proinflammatory T cells, and this allows B. fragilis to reside in the mucosal lining of the gut. In an experimental mouse model for colitis, oral PSA has also been shown to protect against weight loss and decrease proinflammatory cytokine production in the gut. PSA is also able to protect animals for illness in an animal model for autoimmune encephalomyelitis and an animal model for multiple sclerosis (Fig. 17.41).
Fig. 17.41. How the microbiome and the human genome contribute to inflammatory disease. In a simplified model, the community composition of the human microbiome helps to shape the balance between immune regulatory (Treg) and proinflammatory (Th1 7) T cells. The molecules produced by a given microbiome network work with the molecules produced by the human genome to determine this equilibrium. (A) In a healthy microbiome, there is an optimal proportion of both pro- and anti-inflammatory organisms (represented here by segmented filamentous bacteria SFBs and B. fragilis), which provide signals to the developing immune system (controlled by the host genome), leading to a balance of Treg and Th17 cell activities. In this scenario, the host genome can contain "autoimmune-specific" mutations (represented by the stars), but disease does not develop. (B, C) The genomes of patients with multiple sclerosis, type I diabetes, rheumatoid arthritis, and Crohn's disease contain a spectrum of variants that are linked to disease by genome-wide association studies. Environmental influences, however, are risk factors in all of these diseases. Altered community composition of the microbiome due to lifestyle, known as dysbiosis, may represent this disease-modifying component. An increase in proinflammatory microbes (e.g., SFBs in animal models) may promote Th17 cell activity to increase and thus predispose genetically susceptible people to Th1 7-mediated autoimmunity (B). Alternatively, a decrease or absence in anti-inflammatory microbes—for example, B. Iragilis in animal models—may lead to an underdevelopment of Treg cell subsets (C). The imbalance between Th17 cells and Tregs ultimately leads to autoimmunity. (Modified from Lee and Mazmanian, 2010. Used by permission of the publisher.)