INFECTIOUS CANINE HEPATITIS
NICOLA DECARO AND CANIO BUONAVOGLIA
Department of Veterinary Public Health, Faculty of Veterinary Medicine of Bari, Valenzano (Bari), Italy
Infectious canine hepatitis (ICH), also known as Rubarths disease or hepatitis contagiosa canis, is a highly contagious, often fatal, disease of dogs and wild carnivores caused by Canine adenovirus 1 (CAdV1).
Clinical signs were first observed in farmed red foxes (Vulpes vulpes), and the disease was named epizootic fox encephalitis because of the neurological disorders observed in the affected animals. ICH was reported in dogs a few years later and, based on the similar clinical course and pathology, Rubarth suggested a common aetiology for the two diseases. The aetio- logical agent, CAdV1, was isolated a decade later and attenuated through passages on canine and swine cell lines to produce a vaccine. In 1961, another adenovirus of dogs, Canine adenovirus 2 (CAdV2), was isolated from dogs with laryngotracheitis or tracheobronchitis. The isolate, strain Toronto A26/61, was initially considered to be an attenuated strain of CAdV1, and only subsequently was it proposed as the prototype of a distinct canine adenovirus, designated CAdV2.AETIOLOGY
CAdV1, a member of the genus Mastadenovirus within the family Adenoviridae, is highly stable in the environment, surviving several days at room temperature and remaining infectious for months at temperatures below 4° C. The virus is also resistant to various chemicals such as chloroform, ether, acid and formalin. Successful virus inactivation can be obtained with some disinfectants such as iodine, phenol and sodium hydroxide, or with thermal treatment at 60°C for 5 minutes.
CAdV1 is genetically and antigenically related to CAdV2, having a 75% identity at the genetic level and inducing cross - protecting antibodies in the hosts. Although CAdV1 and CAdV2 are strictly related, they have different tissue tropisms and pathogenetic roles.
Vascular endothelial cells and hepatic and renal parenchymal cells are the main targets of CAdV1, whereas the respiratory tract epithelium and, to a limited degree, intestinal epithelium, are the targets of CAdV2. CAdV1 is responsible for a severe, often fatal, disease of domestic dogs and wild carnivores. CAdV2 is associated with infectious tracheobronchitis (also known as kennel cough or canine infectious respiratory disease), an acute, highly contagious respiratory disease of dogs affecting the larynx, trachea, bronchi, and occasionally the lower respiratory tract, that is typically a complex of diseases caused by viral pathogens (CAdVs, canine herpesvirus, canine parainfluenza virus, reoviruses) in association with bacteria, mainly Bordetella bronchisep- tica and Mycoplasma spp. The two CAdV types display different haemagglutination patterns: CAdV1 is able to agglutinate erythrocytes of different species, including birds, whereas CAdV2 haemagglutination is restricted to human and rat erythrocytes.CAdV1 is easily cultivable on a variety of cell lines of different species, including dog, ferret, raccoon and pig. However, the most suitable cell lines used for virus isolation are dog kidney cells, such as Madin-Darby canine kidney (MDCK) cells. Like other adenoviruses, CAdV replicate in the nuclei of the infected cells, inducing intranuclear inclusion bodies.
Epidemiology
CAdV1 has a worldwide distribution in domestic dogs, and several non-domestic carnivore species are susceptible to infection and/or disease. In past decades, epizootic fox encephalitis was very common in European red foxes (Vulpes vulpes) housed on northern American farms. In the wild, cases of overt disease have been reported only sporadically in foxes and other wild carnivores belonging to the families Canidae, Ursidae, Procyonidae and Mustelidae. Recently, CAdV1 was isolated in the UK from the internal organs of three free-ranging European red foxes, with gross lesions and histological changes highly suggestive of epizootic fox encephalitis.
A serological investigation revealed that CAdV1 infection was probably responsible for a dramatic epizootic in red foxes in England and Scotland1-1). Free-ranging red foxes display the highest seroprevalence rates against CAdV, with values of 35% in Germany1-2), less than 20% in Spain(3) and 59.6% in Norway(4). Arctic foxes (Vulpes lagopus) were found to be seropositive to CAdV, with a seroprevalence of 37.8% in the high-arctic islands of Svalbard, and a 67.7% seroprevalence for CAdV was found in the joint Swedish-Norwegian wolf ( Canis lupus) population1-4). Antibodies to CAdV were also detected in free-ranging mustelids in south-western France, including European mink (Mustela lutreola), polecats (Mustela puto- rius), American mink (Mustela υison) and stone martens (Martes foina), with seroprevalences ranging from 2 to 10%(5). A recent serological survey carried out in Spain showed the presence of specific antibodies in the Iberian lynx (Lynx pardinus) and Egyptian mongoose (Herpestes ichneumon)3. ICH was also reported in a captive Eurasian river otter (Lutra lutra)(6). Outbreaks of ICH were also observed in brown bears (Ursus arctos) in Hungary(7). A serological survey carried out on 22 European brown bears from Croatia detected the presence of antibodies against human adenovirus in one animal(8), but a possible cross- r eaction with CAdV1 cannot be ruled out definitively. Outside Europe, reports of ICH in non-domestic carnivores are much more numerous.CAdV1 is shed by animals with active infection through all biological fluids, including saliva, faeces and urine. Recovered animals do not shed CAdV1 in the saliva and faeces, but domestic dogs are recognized to shed the virus in their urine for up to 9 months, so feral dogs may contribute to the spread of the virus to wildlife. Although it is not known how long viruria lasts in non-domestic carnivores, a potential role of these animals as long- term shedders should be considered.
The mortality rates are about 10-30% in domestic dogs and farmed foxes, with peaks of 80% mortality in pups. The high seroprevalence rates found in unvaccinated wildlife may account for a lower pathogenicity of CAdV1 in free-ranging nondomestic carnivores. Therefore, these animals may act as reservoirs of the virus in nature, although there are no specific studies assessing the epidemiological role of nondomestic species.PATHOGENESIS, PATHOLOGY AND IMMUNITY
The main route of infection is by ingestion of virus- contaminated material, although CAdV1 may also be contracted by inhalation of aerosol produced during coughing from animals with CAdV1 respiratory bronchopneumonia. After ingestion or inhalation of infected material, CAdV1 replicates primarily in the tonsils and spreads to the organs and central nervous system via the bloodstream. The main target cells for viral replication seem to be the same regardless of the host species, i.e. the endothelial cells of blood vessels and the hepatocytes. Five days after infection, the virus can be isolated from lungs, liver, spleen, kidney and brain of infected animals. Virus replication in parenchymal cells of liver and lungs and in reticulo-endothelial cells of liver and lymphoid tissues together with infiltration of leucocytes leads to the early appearance of necrotic lesions. CAdV1 infection can cause haemorrhages in many tissues through different pathogenic mechanisms, mainly through extensive vascular damage, impaired hepatic synthesis of clotting factors, thrombocytopenia and disseminated intravascular coagulation. Neurological signs are uncommon in domestic dogs but very frequent in non-domestic carnivores, mainly foxes, as a consequence of cerebral vasculitis. Extensive endothelial damage in the central nervous system may cause hyperacute death without the appearance of observable clinical signs, with the exception of hyperexcitability and convulsions.
An adequate immune response is mounted from 7 days after infection, with production of neutralizing antibodies that are able to clear the virus from the blood and liver, so most animals may recover spontaneously.
However, high- titre antibodies can bind to viral antigens, forming immune complexes that precipitate in the renal glomeruli and irido-corneal angle and cause glomerulonephritis, corneal oedema and uveitis, which are common complications. In the late stages of CAdV1 infection, the virus disappears from the glomerular endothelium but can persist in the tubular epithelium for 6-9 months, causing long- term viruria.Recovery from infection in domestic dogs and foxes is followed by a long- term, probably life-long, protection against CAdV1 disease and infection.
At necropsy, animals that die from hyperacute disease may appear in good nutritional state and with few gross lesions. In acute cases, external examination can reveal ecchymoses and petechial haemorrhages, whereas the abdominal cavity contains abundant clear or serosanguine- ous fluid. The liver is enlarged, yellowish brown, congested and spotted with small circular areas of necrosis; the gallbladder appears thickened, oedematous and greyish or bluish-white opaque in colour. Oedema of the gallbladder wall is a consistent finding. Congestion and haemorrhagic lesions are observed in the spleen, lymph nodes, thymus, pancreas and kidneys. The lungs show patchy areas of consolidation due to bronchopneumonia. Haemorrhagic enteritis can also be observed.
Histological changes in the liver are characterized by Centrolobular necrosis along with neutrophilic and mononuclear cell infiltration and intranuclear inclusions in the Kupffer’s cells and hepatocytes. Intranuclear inclusion bodies can also be seen in the vascular endothelium, renal glomeruli and urinary bladder. Multifocal areas of congestion, haemorrhage and leucocyte infiltration can be observed in several organs, mainly in liver and kidney, owing to vascular damage and inflammation. Nondomestic carnivores usually display multifocal haemorrhages in the central nervous system. Interstitial nephritis and iridocyclitis with corneal oedema are present in dogs recovering from ICH, whereas there is little information in wildlife in Europe about the histological changes associated with CAdVl immune-mediated disease.
CLINICAL SIGNS AND TREATMENT
Clinical signs of ICH are well known in dogs and farmed foxes, whereas there are only sporadic reports in nondomestic carnivores. Virus detection in the faeces of healthy free-ranging wild animals suggests that nondomestic carnivores may become infected without displaying overt disease. Nevertheless, some carnivores (foxes) may be affected by fulminating ICH, especially when a hyperacute neurological form occurs.
The incubation period is usually 4—6 days after ingestion of infectious material. The earliest clinical signs in dogs are represented by a biphasic hyperthermia with temperatures commonly exceeding 40° C. Common clinical signs in all animal species are loss of appetite, depression, vomiting, diarrhoea (often haemorrhagic), abdominal pain, coughing and harsh lower respiratory sounds. Pulse and respiratory rates are accelerated. A serous or purulent ocular and nasal discharge may be present. Haemorrhagic diathesis, as a consequence of the vascular damage, is demonstrated by widespread petechial and ecchymotic haemorrhages and epistaxis. Neurological signs consisting of hyperexcitability, nystagmus and convulsive seizures are rare in dogs but very common in foxes and other nondomestic carnivores. In foxes, the early involvement of the central nervous system may lead to sudden death with few, or no, overt clinical signs. Corneal opacity (‘blue eye’) and interstitial nephritis may occur 1—3 weeks after recovery as a result of the tissue deposition of immune complexes. However, these immune-mediated clinical signs, commonly occurring in domestic dogs, are infrequently seen in wild carnivores. Haematological changes have been described in detail for dogs, whereas tests in non-domestic carnivores have not been reported. Leucopenia with total white blood cell (WBC) counts dogs. Increase in the serum transaminases can be seen only in the severe forms of the disease. Proteinuria (albuminuria) can reach values of >50mg∕dl due to immunomediated glomerulonephritis.
Treatment of ICH is based primarily on treating the clinical signs together with supportive therapy. Dehydration and DIC require administration of fluids, plasma or whole blood transfusions and anticoagulants. Hyperam- monaemia due to hepatic and renal damage can be corrected by oral administration of non-absorbable antibiotics and lactulose, and by oral or parental administration of potassium and urinary acidificants (e.g. ascorbic acid). Supportive therapy may facilitate the clinical recovery of the infected animals, provided that there is sufficient time for hepatocellular regeneration. Broad-spectrum antibiotics can be useful to control bacterial complications.
Passive immunization through administration of hyperimmune sera does not have a therapeutic effect in clinically ill animals, as the onset of clinical signs quickly succeeds to viraemia, and in that stage of infection circulating antibodies are ineffective. By contrast, hyperimmune sera administered to uninfected animals may prevent further spreading of the virus to other animals.
DIAGNOSIS
Clinical laboratory investigations aiming to assess hepatic enzyme activity, WBC counts, haematocrit and proteinuria are of uncertain value in wild species. However, detection of coagulation abnormalities, including thrombocytopenia, altered platelet function and marked prolongation of one-stage prothrombin time, activated partial prothrombin time and thrombin time, may have a certain diagnostic relevance. In dead animals, gross lesions are highly suggestive of CAdV1 infection. Histopathology of liver, spleen, kidney and brain may show hepatic centrilobular necrosis, disseminated vascular damage and intranuclear inclusion bodies. Immunohistochemical techniques applied to formalin- fixed, paraffin-embedded tissues can detect viral antigens in the liver and other parenchymatous organs. CAdV1 can be readily isolated on cell lines of canine origin, such as Madin Darby Canine Kidney (MDCK) cells. CAdV1-induced cytopathic effects consist of clustering of infected cells, detachment from the monolayer and formation of intranuclear inclusions. However, the cyto- pathic effect produced by CAdV2 is indistinguishable from that associated with CAdV1 replication, and CAdV2 may be isolated from the same tissues as CAdV1. Immunofluorescence can detect viral antigens in infected cell cultures and in acetone-fixed tissue sections or smears, but crossreaction with CAdV2 are possible. The adenoviral isolates can be distinguished from their haemagglutination patterns or, more easily, by restriction fragment length polymorphism analysis on viral genomes using the endonucleases PstI and HpaII, which generate differential electrophoretic patterns. Polymerase chain reaction assays have been established that are able to amplify a fragment of the adenoviral genome and to differentiate the two CAdV genotypes.
Serological tests developed for detection of CAdV antibodies are the virus neutralization test, indirect immunofluorescence, haemagglutination inhibition and enzyme-linked immunosorbent assays. These tests are not useful for indirect diagnosis of CAdV1 infection through antibody detection, owing to the cross-reactions with the low pathogenic CAdV2. It is noteworthy that the detection of antibodies in wild animals may reflect the widespread circulation of CAdV1 in wildlife, which is not always associated with clinical disease. In free-ranging wild carnivore populations, serological tests are suitable for epidemiological investigations.
MANAGEMENT, CONTROL AND REGULATIONS
Use of vaccines has greatly reduced the effects of ICH in canine populations. Initial attempts were made with CAdV1-inactivated vaccines, which required repeated inoculations. CAdV1-based modified-live virus (MLV) vaccines proved to be highly effective, but were associated with interstitial nephritis and corneal opacity. Administration of CAdV1 in conjunction with canine distemper virus vaccines was also associated with post-vaccinal encephalitis in domestic dogs. As CAdV1 and CAdV2 are able to induce cross-protection, all current vaccines against ICH are prepared with MLV CAdV2, which does not cause renal or ocular damage.
The CAdV2-attenuated strain Toronto A26/61 is used in most vaccine formulations. In non-domestic carnivores, however, administration of MLV vaccine should be evaluated very carefully, because there have been no extensive vaccination studies to prove the complete safety of MLV formulations. MLV CAdV2-based vaccines have been tested in farmed foxes, proving to be safe and fully protective against CAdV1 infection. A recombinant CAdV2 vaccine expressing glycoprotein G of rabies virus was tested in wildlife species (skunks and raccoons), proving to be completely safe and providing evidence of its promise as a candidate for an oral rabies vaccine(9). Recently, a DNA vaccine was developed against ICH, but its safety and efficacy were evaluated only in mice(10).
Maternally derived antibody (MDA) is likely to interfere with vaccinal immunity in young animals. Oral administration of CAdV2 to non-domestic carnivores resulted in the development of high virus-neutralizing antibody titres. However, further studies are needed to assess whether oral vaccination of wild animals against ICH is effective and safe. All wild animals that are vaccinated against CAdV infection should, where possible, be tested serologically 15—20 days after vaccine administration to determine whether immunization has been successful.
In domestic dogs, vaccination is usually repeated yearly, although after administration of two doses of CAdV2 vaccine, immunity appears to persist for more than 3 years.
PUBLIC HEALTH CONCERN
Although antibodies against CAdV have been found in humans, this seropositivity could be due to cross-reactions with human adenoviruses rather than infection with CAdV1 or CAdV2. Thus, at the moment, CAdV1 is not considered a zoonotic agent.
SIGNIFICANCE AND IMPLICATIONS FOR ANIMAL HEALTH
A CAdV1 carrier status has been suggested for some wild carnivore species and may play a role in the spread of infection to domestic dogs. Feral dogs may have contact with non- domestic carnivores living in suburban areas, such as European red foxes or grey wolves, causing repeated virus introduction into wildlife. Epidemiological and pathogenetic studies would help to define the role of wild carnivores as reservoirs for CAdV1 infection, the susceptibility to CAdV1 disease of the different species of European wild carnivores and the impact of CAdV1 infection in wildlife populations.
AVIAN ADENOVIRUS INFECTIONS
KEVIN EATWELL
Exotic Animal and Wildlife Service, Royal (Dick) School of Veterinary Studies, Edinburgh, Scotland
Adenoviruses that infect avian species have been placed in the genus Aviadenovirus. These are further subdivided into serotypes based on virus neutralization tests. A variety of serotypes have been identified in chickens, quail, turkeys, ducks, pheasants, pigeons, passerines, psittacines, raptors and waterfowl. These are typically classified into three groups, with a number of isolates remaining unclassified.
Group I causes inclusion body hepatitis and hydropericardium in chickens but also disease in quail, turkeys, pigeons, passerines and raptors. These generally are transmitted both horizontally and vertically and lead to severe diseases in birds under 1 month old.
Group II includes haemorrhagic enteritis virus of turkeys, marble spleen disease of pheasants and splenomegaly of chickens. These generally are spread horizontally and cause diseases in older birds, over 1 month of age.
Group III causes egg drop syndrome of chickens and clinical signs such as tracheitis and bronchitis in goslings.
In many species infection can be subclinical, although it has been suggested that adenoviruses form part of a multiple aetiology, rather than acting as primary agents(11). In general adenoviruses are more virulent in non- host- adapted species than in their typical host. Some of these viruses cause significant diseases of wild birds in Europe and others could spill into wild populations from captive birds. In many species, such as raptors, the presence and role of adenoviruses in causing disease is largely unknown, although there are a number of case reports outside of Europe. Adenoviral infections are typically acquired via the faecal-oral route.
I n captive geese high mortality with no premonitory signs is seen in 4- to 11-day-old goslings. Birds die following a brief period of respiratory distress or diarrhoea. More recently a novel presentation of goose adenovirus has emerged in Hungary(12), where affected birds suffered from hepatitic necrosis, focal myocardial necrosis, haemorrhage and hydropericaridium. Intranuclear inclusion bodies were identified on histopathology in the liver.
Marble spleen disease in pheasants occurs worldwide and was first reported in Italy in 1966. However, this disease has only been reported in captive pheasants. The virus can cause sudden death or lead to a brief clinical history of anorexia, depression, diarrhoea and dyspnoea, most commonly in pheasants between 3 and 8 months of age. Post mortem examination reveals splenic enlargement with splenic mottling, and pulmonary oedema. Histopathology reveals reticulum-endothelial cell hyperplasia and lymphoid necrosis. Intranuclear inclusion bodies can be identified in reticulum-endothelial cells.
In Muscovy ducks (Cairina moschata) Duck adenovirus 1 can lead to acute death in captive ducklings up to 35 days of age and is found throughout Europe(13). It has also been identified in captive animals in Canada(14). Clinical signs include coughing, dyspnoea and gasping. Exudates and tracheal plugs can be seen on close examination. Tracheal and bronchial epithelium can be hyperplastic and show eosinophilic inclusion bodies(14).
In 1998 an adenovirus was identified in common eider (Somateria mollissima) in the Baltic Sea(15). The only clinical signs noted were of general illness and moving into shallow waters preceding sudden death of primarily male eiders. At the time of the outbreak the males were courting the females. As courting males can lose considerable body weight during this period, this was believed to be a predisposing factor. It was estimated that 1,000 birds, which was 25% of the local population, died over a period of 6 weeks. The females were observed to continue to lay and incubate their eggs and as a result have minimal social interaction, reducing their risk of disease transmission. The signs noted with this outbreak were similar to those reported before in the Baltic Sea. In this outbreak males in poor body condition developed intestinal disease during the breeding season(16). The birds were in poor body condition with minimal fat reserves. The main lesion, observed in the ten eiders necropsied, was impaction of the proximal small intestine with mucosal necrosis. By histopathology, necrosis of single hepatocytes was observed in two eiders and focal hepatic necrosis in one. No inclusion bodies were seen. There was renal tubular degeneration and congested lungs. Adenovirus was isolated from cloacal swabs from all the dead male birds; it demonstrated a cytopathic effect in Muscovy duck cells within 6 days. Virus neutralization against reference antisera to adenovirus groups I, II and III led to negative results. Virus was not found in any swabs from live females or healthy eider killed by hunting the previous season. Experimental infection of mallard ducklings with the adenovirus from one of the eiders led to positive antibody titres 14 days after inoculation. These ducklings displayed transient shivering, gasping and watery diarrhoea. They were euthanized, and at necropsy they showed distended small intestinal loops with mild chronic inflammation histologically. It was hypothesized that predisposing environmental stressors required to induce mortality were not present in the experimental set-up.
Confirmation of suspected adenoviral disease in dead birds should initially be based on histopathological changes. Virus isolation and viral neutralization can be used for confirmation in both live and dead birds.
There is currently no specific treatment for adenoviral infections in birds. Supportive care measures, reducing stocking density (in rehabilitation facilities) and ensuring that good basic hygiene standards and biosecurity measures are maintained can together limit the spread of disease. As adenoviruses may not be the primary agents involved with this mortality, appropriate investigation of any deaths should include a thorough evaluation of other pathogenic agents involved.
ADENOVIRUS INFECTION IN SQUIRRELS
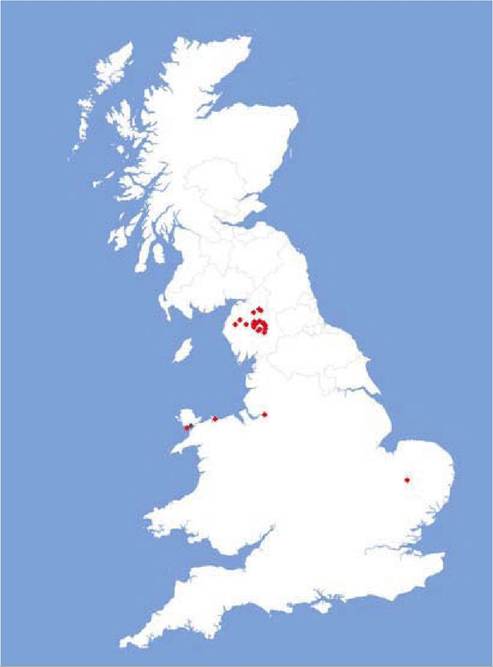
FIGURE 14.1 Red and grey squirrel adenovirus-positive cases in the UK diagnosed up to October 2009 with scanning electron microscopy and/or polymerase chain reaction (PCR). The one grey squirrel case was subclinical, PCR- positive. Reproduced with kind permission of J. Paul Duff and David Everest of the Veterinary Laboratories Agency and Mark Wilkinson of the Northumberland Wildlife Trust.
The disease caused by SqAdV is characterized by acute enteritis and death in red squirrels (Sciurus vulgaris).
Enteric adenovirus infection of wild red squirrels has so far been reported only from various parts of the UK (e.g. Cumbria, Norfolk, the island of Anglesey and Scotland), often associated with translocation/ reintroduction events and in captive animals(18-20) (see Figure 14.1). The fatal disease caused by SqAdV affects red squirrels of all age classes, but the same virus has also been identified in healthy grey squirrels (Sciurus carolinensis) on Anglesey. There is no information about the presence of this virus in continental Europe or elsewhere. Reports indicate a tendency towards the clustering of cases, suggesting local outbreaks of disease, and the most likely route of viral transmission is through direct contact with infected animals. It is unclear yet which species serve as a reservoir of the virus, one possible candidate is the grey squirrel(20) — although other rodents or the red squirrel itself cannot be excluded.
A review of 36 cases of adenovirus enteritis diagnosed from Animal Health Veterinary Laboratory Agency Diseases of Wildlife Scheme data in the UK (Duff, unpublished observations) showed that there was no obvious seasonality in this admittedly limited dataset. From the cases where age was discernable, 21 were adults and 12 were immature, supporting the premise that both young and adult animals are susceptible. The disease appears to be acute in nature — only five animals were found ill, weak and collapsed, and these died within hours of capture; the remainder were found dead, often in nest boxes. An additional indicator of the acute nature of the enteritis was that only 15 of the 36 had diarrhoeic staining of fur around the anus.
The infection appears to affect primarily the mucosae of the small and large intestine, but the presence of SqAdV was also demonstrated by polymerase chain reaction (PCR) in the spleen of a healthy grey squirrel. Gross lesions are few and limited to the gastrointestinal tract. The most consistent findings included mildly congested and distended intestines with profuse yellow coloured watery/ mucoid content, often stained with blood. Disc- shaped hyperaemic areas 2—3 mm in diameter were also observed on serosal surfaces of both small and large intestines in a few individuals1-21). In some animals there was splenic congestion, but no other consistent gross visceral lesions were recorded. Histological lesions include acute exfoliating enteropathy associated with villous atrophy, crypt hyperplasia, and the presence of solid amphophylic intranuclear inclusion bodies in the villous enterocytes surrounded with a mild infiltrate of mononuclear inflammatory cells(21). Acute splenitis and lympholysis can also be observed in some cases.
Laboratory diagnosis is based on the demonstration of SqAdV by SEM, virus isolation and PCR on clinical samples (faeces, faecal swabs). The PCR targets either the hexon or polymerase gene(17’20) and the product is further characterized by sequencing. Post mortem examination and histopathology support the diagnosis of SqAdV infection. Serological methods have not yet been developed. Treatment has not been described, and it is not known whether affected animals may respond to supportive treatment.
The risk of spreading SqAdV by translocation and reintroduction of red squirrels should be taken into consideration, along with potential danger to naive, susceptible animals introduced to an infected area. Adenovirus infection of red squirrels may have significant impact on the survival and conservation of this seriously threatened species in Britain.