INTRODUCTION TO PHYLUM Platyhelminthes
Platyhelminths (platy: flat, helminth: worm) are a phylum of simple, soft-bodied, dorsoventrally flattened invertebrates with bilateral symmetry. While there is a blind gut in some, none have a body cavity (coelom); thus, platyhelminths are classified as acoe- lomates (Roberts and Janovy 2000).
They are among the simplest animals to have a nervous system. Platyhelminths of interest for wildlife belong to the Class Cercomeridia (App. 1: Table 3). Two platyhelminth groups are of major importance in wildlife disease studies: trematodes (Subclass Trematoda), also called flukes, and cestodes (Infraclass Cestoidea), also called tapeworms (Roberts and Janovy 2000) (App. 1: Table 3). Although there are many free-living platyhelminths, all trematodes and cestodes engage in parasitic life styles.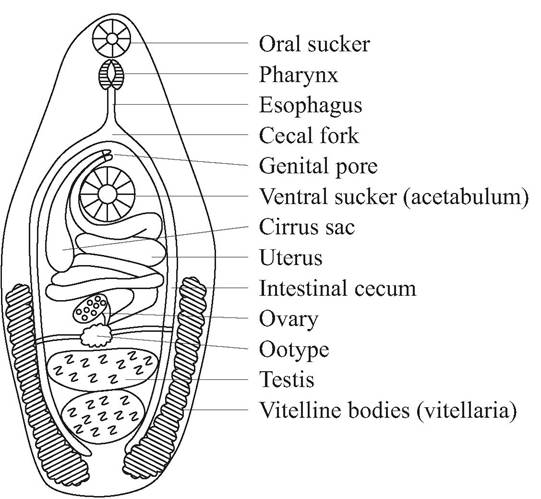
FIGURE 4.1 Generalized anatomy of a digenetic trematode (McDonald, 1981; courtesy of Malcolm McDonald and the U. S. Fish and Wildlife Service).
TREMATODES (FLUKES)
Trematodes are characterized as having an oral sucker that includes a pharynx, and with a variably placed ventral sucker (acetabulum) as a holdfast organ (Roberts and Janovy 2000) (Fig. 4.1); a few trematodes have only one sucker, usually at the anterior end (Bush et al. 2001). While lacking coeloms, trematodes have well-developed mesoderms that form the parenchyma, reproductive organs, and musculature in the adult stages (Roberts and Janovy 2000). Almost all are hermaphroditic (monoecious), with complete male and female reproductive systems in each individual; while some are capable of self-fertilization, most require cross-fertilization to produce viable progeny (Roberts and Janovy 2000).
As with many parasite groups, trematodes have undergone extensive taxonomic revisions in recent years (Littlewood and Bray 2001, Gibson et al.
2002, Jones et al. 2005), and are estimated to include between 18,000 (Cribb et al. 2001) and over 24,000 (Poulin and Morand 2004) species. At least 530 species have been identified in waterfowl alone, for example (McDonald 1981). While advanced molecular techniques to determine phylogenetic relationships grew rapidly in the 1990s, considerable uncertainty on these taxonomic relationships remains (Cribb et al. 2001). Among trematodes, the parasites of interest to wildlife diseases fall primarily into the Infraclass Digenea, the digenetic trematodes, and further discussion of trematodes will be focused on this group.Digenetic Trematodes
Digenetic trematodes are not segmented, and their intestinal tract is a bifurcated blind sac (Fig. 4.1). Digenetic trematodes parasitize all classes of vertebrates; nearly every organ of the vertebrate body can be parasitized by the adult or juvenile form of some species of trematode (Roberts and Janovy 2000). Various trematodes can cause liver disease, respiratory diseases such as pneumonia or pleurisy, intestinal inflammation with bleeding and diarrhea, neurological diseases such as encephalitis and ataxia, muscle weakness or pain, and some multisystemic diseases such as anemia. Descriptions of pathogenic trematodes found in wild mammals (Davis and Libke 1971, Pybus 2001) and birds (Wobeser 1997, Huffman 2008, Huffman and Fried 2008) are available.
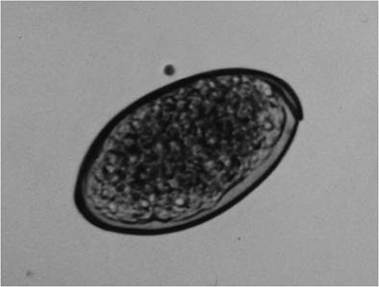
FIGURE 4.3 Trematode ova from the liver fluke, Fasciola hepatica (Courtesy of W. Frank, Universitat Hohenheim, Germany).
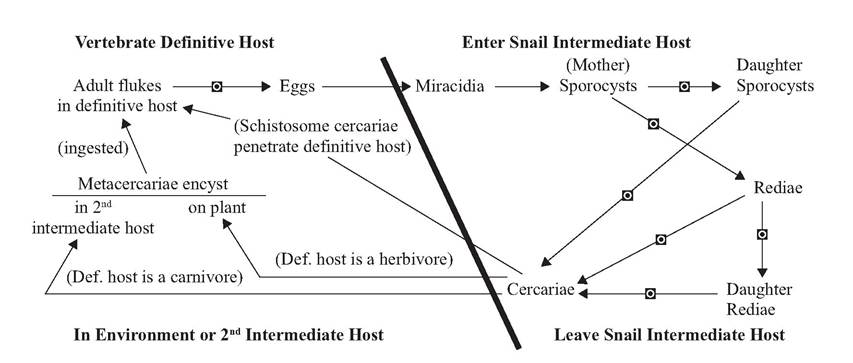
FIGURE 4.2 Life cycle of a digenetic trematode. Ξ: Indicates life cycle transitions in which multiplication occurs.
DIGENETIC TREMATODE LIFE CYCLES Digenetic refers to two (“di”) generations/produc- tions (“gen”), the characteristic that the life cycle of these parasites involves at least two hosts; that is, they are heteroxenous and have an indirect life cycle, involving a definitive host in which the sexually mature stage of the parasite occurs, and at least one intermediate host, in which asexual development and multiplication of an immature larval stage occurs (Fig.
4.2) (Carus 1863). The first intermediate host is a mollusc such as a snail and, for many digenetic trematodes, there is only one intermediate host; however, some digenetic trematodes may have a second or occasionally even a third intermediate host (Roberts and Janovy 2000).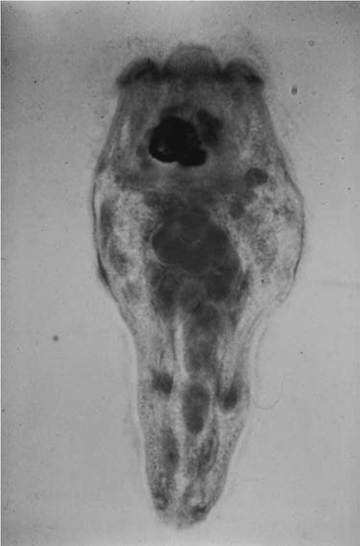
FIGURE 4.4 Miracidium of Fasciola hepatica (Courtesy of W. Frank, Universitat Hohenheim, Germany).
Adults produce operculated eggs (Fig. 4.3), with up to 25,000 per day for some liver flukes. The eggs escape with the feces, urine, or sputum, depending on the site of the adult in the host.
A larval stage called the miracidium (Fig. 4.4) develops in the egg. There are two strategies used by miracidia to infect their snail
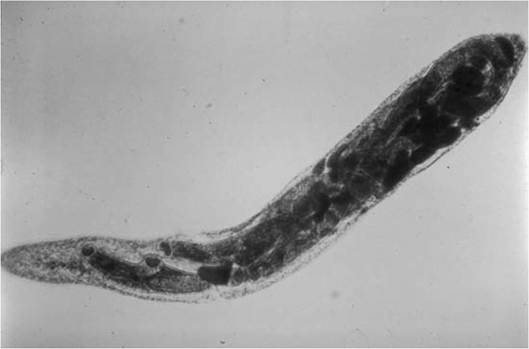
FIGURE 4.5 Redia of Fasciola hepatica; this redia contains developing cercariae (Courtesy of W. Frank, Universitat Hohenheim, Germany).
intermediate host (Fig. 4.2). For most Digenea, the ciliated miracidium hatches from the egg and actively seeks out the proper intermediate host. It has about 24 hours to find and penetrate its snail host before depleting its energy reserves. In contrast, miracidia of some species of Digenea remain in the egg and wait to be eaten. They have no control over whether an appropriate intermediate host will ingest them, but the miracidia often can survive for months or years in the egg.
Once a miracidium reaches a proper intermediate host, it settles in the appropriate tissue (e.g., intestine, gonad, digestive gland) and metamorphoses into a simple sac-like form called the sporocyst. In some species, the first generation (mother) sporocyst will develop similar cells, called daughter sporocysts. But within sporocysts of most species of trematodes, a number of asexual embryos develop, called rediae (Fig. 4.5), which have a few rudimentary appendages such as a pharynx and a sac-like gut.
Some trematodes also develop daughter rediae. Eventually the rediae (or daughter sporocysts) develop additional embryos that become cercariae (Fig. 4.2).Cercariae (Fig. 4.6) undergo no multiplication; they are juvenile forms that will develop into the adult. They have a digestive tract and suckers, and often have genital primordia present (Roberts and Janovy 2000). Cercariae emerge from the snail intermediate host and must eventually reach the definitive host; most cercariae have tails for swimming.
Cercariae use one of three strategies to find their definitive hosts (Fig. 4.2). In a few cases, as with important human parasites of the Superfamily Schistosomatoidea, cercariae actively seek out and penetrate the definitive host. However, for most members of the Digenea, the cercariae first encyst in the environment or in a second intermediate host with a stage called the meta- cercaria. If the definitive host is an herbivore, the cercariae usually swim to plants in the environment and encyst on them to await their ingestion by the herbivorous host. If the definitive host is a carnivore, the cercariae seek out and encyst on or in potential prey species to form metacercariae and await their ingestion by the definitive host. Second intermediate hosts can be other molluscs, crustaceans, fish, amphibians, reptiles, or other mammals; occasionally a bird may serve as the second intermediate host. The predatory behaviors of freshwater invertebrates may reduce success of cercariae in completing their life cycles (Schotthoefer et al. 2007).
Two types of glands can be associated with cercariae. Cystogenous (cysto: cyst; gen: producing) glands are used by all cercariae forming metacercariae. Histolytic (histo: tissue; lytic: to lyse) glands can lyse host tissues and are used by cercariae to escape from their snail hosts through solid tissue as well as to penetrate the next intermediate or definitive host, as needed. Once metacercariae become fully developed, they are infective to the definitive host and can
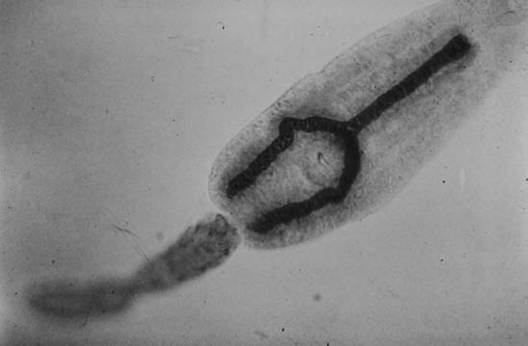
FIGURE 4.6 Cercaria of Fasciola hepatica (Courtesy of W.
Frank, Universitat Hohenheim, Germany).complete development into adult trematodes (Roberts and Janovy 2000).
With such complicated life cycles, it would seem improbable for any trematode to ever be able to complete all stages. The primary strategy used by trematodes to overcome these odds is extensive multiplication—flooding the environment with parasite stages (Fig. 4.2). Thus, a single adult liver fluke may lay up to 25,000 eggs per day and produce eggs for many years (Happich and Boray 1969). One egg leads to one miracid- ium. But one miracidium entering a snail may produce from hundreds to millions of cercariae. Each cercaria will produce only one adult trematode. Those numbers are needed for one life cycle completion to replace one parent trematode!
With such a fixed and complicated life cycle, there are several points at which the life cycle can be interrupted for purposes of trematode control. Among nonhuman animals, two general strategies commonly are used in trematode control: control of the molluscan intermediate host, and immunization or treatment of the definitive host. Control of the molluscan host typically has involved one of three strategies: molluscicides, predators, and habitat manipulation. Of these, molluscicides have been most common. For many years, various forms of copper, a heavy metal, were applied to habitats with snail intermediate hosts (Gordon et al. 1959, Faust and Russel 1964). While very effective at killing snails and generally not toxic for mammals at low levels, copper can be toxic to many vertebrates and invertebrates, especially if application is not carefully controlled.
The World Health Organization defined characteristics of suitable molluscicides to include safety in respect to acute and chronic mammalian toxicity, not producing adverse effects if it enters the food chain, and evidence that it is stable in storage for at least two years (McCullough and Mott 1983). However, even under the best of circumstances, molluscicides affect species beyond those of just the target snail intermediate hosts.
A proposed alternative, biological method of mollusc control has been use of predators such as ducks or crayfish (Jobin et al. 1973); however, this approach has had inconsistent results and has not been well developed. Interestingly, some trematodes can castrate their snail intermediate hosts. For example, infections of snails by the trematode Ribeiroia marini cause complete castration of Biom- phalaria glabrata, a prominent snail host in the life cycle of human schistosomes (Golvan et al. 1974, Nassi et al. 1979). Large-scale introduction of Ribeiroia marini can eliminate populations of this planorbid snail. Rediae of Ribeiroia spp. also actively consume schistosome sporocysts, further reducing the production of Schistosoma mansoni cercariae (Johnson et al. 2004). While unusual, such insights can lead to further innovations in trematode control in the future.
Habitat manipulation has been used to control trematodes. In Lake Victoria, Africa, vegetation along water channels around the lake was cleared to remove snail habitat; soon afterward predatory birds and frogs further depressed the snail population (Ogamlo-Ogoma 1972). One treatment effectively removed predators for about 8 to 9 weeks, but the snail population returned to pretreatment levels by about 11 weeks. Thus, treatment every 8 weeks would be needed to be effective. And, of course, drastic changes in a habitat affect far more species than just the targeted snail intermediate hosts.
Treatment and immunization of infected or at-risk definitive hosts are other options. A considerable number of chemicals are available to treat trematodes in domestic or captive definitive hosts (Fraser and Mays 1986, Fowler et al. 2003). In the United States, the Animal Medicinal Drug Use Clarification Act (1994) allows limited use of some licensed drugs for off-label use in wildlife that are not potential food animals (Waldrup 1998) (www.fda.gov/ora/compliance_ref/cpg/ cpgvet/cpg615-115.html). Such regulations can vary considerably among countries. However, there appears to be adequate opportunity for further research on the topic of chemical treatment of wildlife to control helminths. In Europe, chemicals such as rafoxanide reduce liver flukes, Fasciola hepatica, among roe deer (Capreolus capreolus) and red deer (Cervus elaphus) through incorporation into feeding stations; this chemical is also effective against some nematodes and nasal bot flies (Barth and Schaich 1973a, 1973b). Additional chemicals, including praziquantel, triclabendazole, and albendazole, have had varying success against other trematodes affecting wildlife (Foreyt and Gorham 1988a, 1988b; Qureshi et al. 1989, 1990).
Although less readily applicable to wildlife hosts, immunization is a possibility. Wildlife exposed to some trematodes can develop immune responses to them (Macy 1973). Vaccines continue being developed against liver flukes (Spithill and Dalton 1998).
Although most trematodes do not cause significant mortality among wildlife, some species can be of concern to managers. A number of these trematodes affecting wildlife will be considered.
Liver Trematodes