MORBILLIVIRUS INFECTIONS IN AQUATIC MAMMALS
jolianne m. rijks 1, albertd.m.e. osterhaus2 AND THIJS KUIKEN2
1Dutch Wildlife Health Centre, Utrecht, The Netherlands 2Department of Virology, Erasmus MC, and Artemis Riesearch Institute for Wildlife Health in Europe, Rotterdam, The Netherlands
AETIOLOGY
Five morbilliviruses have been reported to cause natural infections in aquatic mammals (Table 7.2):
TABLE 7.2 Method of diagnosis, time, location and impact of morbillivirus infections in aquatic mammals in Europe.
| Morbillivirus | Host species | Diagnosed by | Year(s) | Geographical location | Estimated number affected | Reference |
| Canine distemper | Caspian seal (Pusa caspica) | RT-PCR | 1997 | Caspian Sea | 47 | |
| virus | Caspian seal | RT-PCR | 2000 | Caspian Sea | >10 000 | 5, 48 |
| Phocine | Harbour seal (Phoca | Virus | 1988 | Kattegat, Skagerrak, | 23 000 | 49 |
| distemper virus | vitulina) | isolation | Baltic Sea, Wadden | 50, 51 | ||
| Sea, North Sea, N.E. Atlantic | 52 | |||||
| Harbour seal | RT-PCR | 2002 | Kattegat, Skagerrak, | 30 000 | 53, 54 | |
| (Grey seal, Halichoerus | Baltic Sea, Wadden | |||||
| grypus) | Sea, North Sea, N.E. Atlantic | |||||
| Porpoise | Harbour porpoise (Phocoena | Virus | 1988, 1990 | North Sea, N.E. | 10 | 55, 56 |
| morbillivirus | phocoena) | isolation | Atlantic | 57 | ||
| Mediterranean monk seal | Virus | 1996 | Mediterranean Sea | 1 | 58 | |
| (Monachus monachus) | isolation | (Greece) | ||||
| Dolphin | Striped dolphin (Stenella | Virus | 1990-1992 | Mediterranean Sea | > 400 | 59 |
| morbillivirus | coeruleoalba) | isolation | ||||
| Long-finned pilot whale | RT-PCR | 2006-2007 | Mediterranean Sea | > 21 | 60 | |
| (Globicephala melas) Striped dolphin | RT-PCR | 2007 | Mediterranean Sea, | >100 | 61 | |
| Striped dolphin, Bottlenose | RT-PCR | 2007-2008 | Ligurian Sea | 143 | 62 | |
| dolphin ( Tursiops truncatus), Long-finned pilot whale | Mediterranean Sea | |||||
| White-beaked dolphin | Virus | 2007 | North Sea (Germany) | 1 | 63 | |
| (Lagenorhynchus albirostris) | isolation | |||||
| Common dolphin | RT-PCR | 1994 | Black Sea | ? | 64 | |
| (Delphinus delphis) | ||||||
| Pilot whale | Fin whale (Balaenoptera | Histology | 1997-1998 | North Sea | 2 | 65 |
| morbillivirus | physalus) | and IHC | ||||
| Short-finned pilot whale (Globicephala macrorhynchu$) | Histology | 1986 | Atlantic, Canary Islands | 1 | 66 |
aPorporoise-morbillivirus-like virus
• Canine distemper virus (CDV) infection (canine distemper; synonym PDV2 infection);
• Phocine distemper virus (PDV) infection (phocine distemper; synonyms seal distemper, seal plague, PDV1 infection);
• Dolphin morbillivirus (DMV) infection;
• Porpoise morbillivirus (PMV) infection;
• Pilot whale morbillivirus (PWMV) infection.
For consistency in this section of the chapter, the terms ‘CDV disease’ rather than ‘canine distemper’, and ‘PDV disease’ rather than ‘phocine distemper’, are used.
EPIDEMIOLOGY
In 1988, about 50% of the North-East Atlantic population of harbour seals (Phoca vitulina) died within a few months. The causative agent was identified as a new morbillivirus, PDV(49-51). A PDV epidemic of similar scale occurred for a second time in 2002, after the harbour seal population had returned to pre-1988 levels(53). In 1988 a second new morbillivirus, PMV, was identified in harbour porpoises (Phocoena phocoena) that stranded on the coast of Northern Ireland(55). A third new morbillivirus, DMV, was identified as the cause of a large epidemic in striped dolphins (Stenella coeruleoalba)(,T This epidemic started in the western Mediterranean Sea in 1990 and spread eastwards in 1991 and 1992. Two years later, an epidemic of morbillivirus disease occurred in common dolphins (Delphinus delphis) in the adjacent Black Sea(64). Phylogenetic analysis based on the sequence of a nucleoprotein gene fragment showed 97% identity with dolphin morbillivirus (M. van de Bildt, personal communication). A second smaller DMV epidemic in striped dolphins in the western Mediterranean Sea occurred in 2007, and was preceded in 2006—2007 by an epidemic from the same virus in long-finned pilot whales (Globicephala melas) found stranded in the Strait of Gibraltar and southern Spain(60’61). The Caspian seal (Pusa caspica) population, which is restricted to the Caspian Sea, suffered a large CDV epidemic in 2000(5). In addition, infections with different morbilliviruses, including PWMV, have been identified in individual strandings of marine mammals in Europe (Table 7.2).
GEOGRAPHICAL DISTRIBUTION IN EUROPE
Morbillivirus infections have been diagnosed in multiple species of aquatic mammals in all the major waters of Europe, including the (North-)East Atlantic Ocean and the North, Baltic, Mediterranean, Black and Caspian Seas (Table 7.2).
Rather than being endemic, they occur as epidemics in naive populations or as isolated cases.HOST FACTORS
Marine mammal species affected by morbillivirus disease belong to the suborders Pinnipedia, Odontoceti and Mystacoceti. There also is evidence of morbillivirus infection in the order Sirenia and in polar bears (Ursus maritimus) by serology and in sea otters (Enhydra lutris) by RT-PCR(67’68). Therefore, many species of aquatic mammals should be considered susceptible to morbillivi- rus infection.
Many of the aquatic mammal species occurring in Europe are susceptible to fatal disease from morbillivirus infection: harbour seal (PDV), Caspian seal (CDV), harbour porpoise (PMV), striped dolphin (DMV), bottlenose dolphin (Tursiops truncatus; DMV and PMV)(69), long-finned pilot whale (Globicephala melas; DMV and PWMV)(70), white-beaked dolphin (Lagenorhynchus albi- rostris; DMV), common dolphin (DMV), short-finned pilot whale ( Globicephala macrorhynchus; PWMV) and fin whale (Balaenoptera physalus( unidentified morbillivirus) (Table 7.2). Some species are known to be susceptible to morbillivirus infection, but the pathogenicity of these infections is not known, e.g. Mediterranean monk seal (Monachus monachus( PMV-like virus, DMV)(58), harp seal (Pagophilus groenlandicus; PDV), hooded seal (Cystophora cristatd)771. The grey seal is unusual, because it appears relatively resistant to disease by at least one morbillivirus, PDV, based both on field and experimental data(72’73).
Aquatic mammals of all ages and both sexes appear to be susceptible to morbillivirus infection and disease, judging from their representation during reported epidemics and post- epidemic serological studies. However, age and sex may affect risk of infection and the severity of disease. PDV appeared to spread through the subadult fraction of the harbour seal population before the juvenile and adult fraction in the Dutch 2002 epidemic(74), indicating different probabilities of disease transmission among different age cohorts(75).
DMV affected mainly juvenile striped dolphins during the 2007 epidemic. A likely explanation is that the older animals that survived the 1990 epidemic had developed protective immunity(62). PDV appeared to spread faster among males than among females of all age categories during the 2002 epidemic; possible reasons were higher contact rates among males and higher contaminant levels in tissues of male seals(54’74).ENVIRONMENTAL FACTORS
Pollution, climate and fisheries activities have been associated with triggering the occurrence of morbillivirus epidemics in aquatic mammals, with enhancing mortality, or both. The tissues of harbour seals that died during the 1988 epidemic had high levels of pollutants, including polychlorinated biphenyls (PCBs) and dichlorodiphenyltrichloroethane (DDT). Based on a semi-field study comparing the immunological parameters in harbour seals fed contaminated or clean fish, it was concluded that the pollutant levels found in 1988 were sufficiently high to have caused immune suppression1-76). The same may have occurred during the 2002 epidemic in harbour seals, because the levels of PCBs had not decreased significantly (Kajiwara et al., unpublished data). Pollution may also have exacerbated the 1990 epidemic in striped dolphins, because tissues of animals that died during the epidemic had higher pollutant levels than those that stranded before(77). By contrast, the tissue levels of pollutants in Caspian seals that died during the 2000 epidemic were similar to those found in apparently healthy animals from previous years(48), as was also the case for striped dolphins stranded during the 2007—2008 epidemic, and before and after it(77).
The winters preceding the 1988 PDV, the 1990 DMV, and the 2000 CDV epidemics were unusually mild. It is speculative, but possible, that such weather events may trigger morbillivirus epidemics in aquatic mammals.
Hypotheses include increased hauling-out patterns of seals, resulting in higher contact rates, and changes in fish abundance and seal species distribution, the latter resulting in a higher risk of cross-species transmission of morbillivirus(48).Fisheries activities were speculated to have triggered the 1988 PDV epidemic indirectly. An unusual southward migration in the preceding winter brought harp seals — considered, on the basis of serological studies, to have been exposed and therefore a possible source of PDV — into the range of the North-East Atlantic harbour seal population. This movement may have been triggered by overharvesting of the fish stocks around Greenland, where harp seals normally forage(54).
The marine mammal populations in Europe that have undergone morbillivirus epidemics are probably not the reservoirs for these viruses; these reservoirs are unknown. The North-East Atlantic harbour seal population of about 50,000 is probably not a reservoir for PDV, as sera from before 1988 lacked specific antibody, seroprevalence declined between the 1988 and 2002 epidemics, and no infections were reported in this inter-epidemic period. Instead, the reservoir is speculated to be in aquatic mammals, e.g. harp seals, from the Northern Atlantic or Arctic Oceans, based on high seroprevalence in those populations, and one probable clinical case in a juvenile harp seal from Canada(78). In any case, the P- and H-gene sequences of the 1988 and 2002 PDV isolates were virtually identical and thus probably had the same source1-53’54).
The striped dolphin population of the Mediterranean Sea is probably not the reservoir of DMV, based on declining seroprevalence and lack of reported DMV infections between the 1990—1994 and 2007 epidemics. The reservoir of DMV is speculated to be in aquatic mammal populations in the Atlantic Ocean, e.g. in long- finned pilot whales, in which the seroprevalence is high(79). Long- finned pilot whales with DMV infection in which the virus is genetically nearly identical to that in striped dolphins were found stranded along the Strait of Gibraltar just before the 2007 epidemic, and may have introduced DMV into the Mediterranean Sea via this route1-60’61).
The reservoir for CDV that affected Caspian seals in 2000 could be the Caspian seal population itself, which was estimated at 111,000 in 2005(80). Antibody to CDV was detected in Caspian seal sera as far back as 1993, and P- and F- gene sequences of the CDV from 2000 were identical to those of a CDV found in a Caspian seal in 1997. Alternatively, CDV could have been reintroduced repeatedly into Caspian seals from adjacent populations of terrestrial carnivores, e.g. domestic dogs or wolves(5,48).
It is not clear whether the North-East Atlantic Ocean harbour porpoise population of about 385,000(80) is a reservoir for PMV. Specific antibody was detected in harbour porpoises collected between 1991 and 1999 from the North and Baltic Seas and from the North-East Atlantic Ocean(79,81). However, PMV has not been detected in harbour porpoises since its report in 10 harbour porpoises from the North and Irish Seas between 1988 and 1992(56,57,67), despite examination of over 500 carcasses found stranded or by-caught in the North Sea and NorthEast Atlantic Ocean between 1990 and 2000(82-84).
The little information available about morbillivirus infections in aquatic mammals is skewed towards coastal species in temperate climates. Epidemics occurring in more pelagic species, such as white-beaked dolphins and fin whales, may go unnoticed because their carcasses are less likely to reach a coast. Epidemics occurring in polar species, such as harp seals, also may go unnoticed because of low human densities in their range. Therefore, it is even more difficult to make any conclusions on the role of these pelagic and polar species in the epidemiology of morbil- livirus infections.
TRANSMISSION
The route of transmission of morbilliviruses in aquatic mammals is speculative and experimental data is scant, but it is assumed to be similar to that in terrestrial mammals. PDV was isolated from nasal fluid of a harbour seal on day 11 post-inoculation(72).
The transmission of PDV in harbour seals may occur when they are in the water, where they have more direct contact than when they are hauled out on land. On land, direct contact is less common, but transmission by indirect contact might take place via infectious aerosols, secreta and excreta. Wind gusts could favour horizontal and concentrated movement of aerosolized virus.
Transmission of DMV in dolphins may take place by direct contact, e.g. during coitus, or by indirect contact, e.g. by aerosol(67). Transmission by inhalation of expired blowhole secreta might be important because of the combination of aggregating in large schools, forceful expiration and inspiration, and breathing in a synchronized fashion (85).
PATHOGENESIS, PATHOLOGY AND IMMUNITY
Little is known about the pathogenesis of morbillivirus disease in aquatic mammals. Current concepts come largely from pathogenesis studies of morbillivirus infections in terrestrial mammals, supplemented by observations from both natural and experimental infections in aquatic mammals. Although morbillivirus in terrestrial mammals may enter via other sites (e.g. conjunctiva, digestive tract, urogenital tract and skin wounds), the main site of entry is assumed to be the respiratory tract. Harbour seals were successfully infected experimentally by droplets on the nasal mucosa(72). Even in the respiratory tract, the primary target cells are probably immune cells expressing signalling lymphocyte activation molecule (SLAM), the receptor for morbillivirus. Peripheral blood leucocytes of aquatic mammals also express SLAM(86). The first tissues found positive after infection are the lymph nodes of the head and lower respiratory tract.
Morbillivirus has a high infectivity, demonstrated for PDV in harbour seals, where a low dose is sufficient to cause fatal infection(87). After amplification in the regional lymph nodes of the head and lower respiratory tract, the virus spreads via the blood to other lymphoid tissues, where a second cycle of amplification takes place. In experimentally infected harbour seals, leucocyte-associated viraemia occurs from 5 to 12 days post-inoculation, and sometimes beyond(72). The resulting infection of the lymphoid tissues weakens host immunity, demonstrated in aquatic mammals by the severe lymphoid depletion of lymphoid organs associated with virus infection. The third stage of virus spread is to the epithelia of multiple organ systems (respiratory, digestive, urogenital), to the central nervous system (for some morbillivirus species), and to a variety of miscellaneous cell types such as fibroblasts. This systemic spread of morbillivirus also occurs in aquatic mammals(72). Because not all these cell types express SLAM, morbilliviruses presumably have an alternative mechanism to enter cells.
Morbillivirus-induced immune depression allows facultative pathogens to spread and cause severe secondary disease. An example in PDV- infected seals is Bordetella bronchiseptica, which can spread to the lower respiratory tract and cause bronchopneumonia1-88’89). Spread of morbil- livirus to the central nervous system causes necrosis of neurons and glial cells and associated inflammation, resulting in severe nervous dysfunction. Spread of morbillivirus to epithelial cells in various organ systems may lead to the development of intranuclear inclusion bodies (INIB) and intracytoplasmic inclusion bodies (ICIB), which are not necessarily associated with severe disease.
The speed and level of the immune response may influence the extent of morbillivirus spread throughout the body. Despite lymphoid depletion, infected hosts can mount an immune response. In harbour seals experimentally infected with PDV, neutralizing antibodies were first detected at 7 days post-infection and reached a maximum 3 weeks later(72). An experimentally infected grey seal also developed antibodies1-87). Harbour seal pups, of naturally infected mothers, had detectable maternal antibodies until 3 months after birth, at which time they were fully susceptible to PDV infection(72).
In host species susceptible to morbillivirus disease, the vast majority of morbillivirus infections end abruptly within days after the onset of clinical signs, either because the host dies, or because the host has mounted an immune response that clears the virus from tissues. The infectious period is therefore generally short. It is not always fully understood how death is caused. Some hosts may die from neurological complications resulting directly from virus infection of the central nervous system, and others from disease caused by a facultative pathogen secondary to virus-induced immune depression. Many seals and dolphins dying during morbillivirus epidemics had a morbillivirus-associated encephalitis, a secondary bacterial bronchopneumonia, or both(88,89). In hosts that completely clear the infection, the immunity is long-term, often lifelong, which explains why virtually no harbour seals older than 14 years stranded during the 2002 PDV epidemic, and why most of the striped dolphins that stranded during the 2007 DMV epidemic were juveniles(61,90).
Occasionally, viral clearance is incomplete. In particular, this involves animals with virus infection in the brain, which usually die within a few months. They are not known to be infectious beyond 2 months after infection. Very rarely, neurological disease only develops years after infection. In such cases, virus is restricted to the brain and may
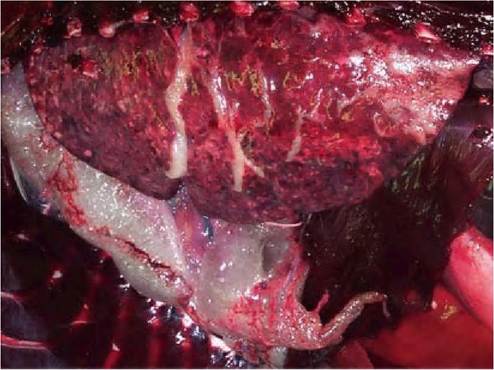
FIGURE 7.4 Pulmonary and mediastinal emphysema in a harbour seal with PDV infection. The ventral part of the lung is consolidated. Reprinted with permission from Rijks et al., 2008.(89)
be defective. Neurological cases after epidemics have been documented in marine mammals. Up to 4 years after the 1990 DMV epidemic, striped dolphins were found stranded dead with a DMV infection restricted to the central nervous system and associated with encephalitis (91). Also, the authors have diagnosed encephalomyelitis in an approximately 14-year-old harbour seal in 2001, 13 years after the 1988 PDV epidemic (unpublished data). In this animal, infection was restricted to the central nervous system with a PDV variant that had a genetic signature that indicated it was defective, which suggests that it had survived for 13 years after becoming infected with PDV in 1988.
Aquatic mammals that die with morbillivirus infection have a nutritional state ranging from poor to good, and generally no food in the stomach1-48’60’64’89’92-94). The main gross lesions in several, but not all, aquatic mammal species with morbillivirus infection are in the lungs. Harbour seals with PDV infection, Caspian seals with CDV infection, and harbour porpoises with PMV infection have mutifocal pulmonary consolidation (Figure 7.4) characterized by irregular, red or grey, firm areas that sink in water. This is caused at least partly by secondary bacterial infections, of which B. bronchiseptica is most important in harbour and Caspian seals. Striped dolphins with DMV infection have multifocal atelectasis. A related lesion in harbour seals with PDV infection but not in other aquatic mammal species is marked emphysema, which is usually mediastinal, pericardial, retroperitoneal and subcutaneous (Figure 7.4). The occurrence of emphysema in harbour seals may be due to well-developed interlobular septa that prevent collateral ventilation in case of airway obstruction1-89).
No gross pulmonary lesions were recorded in striped dolphins that stranded after the main part of the 1990 DMV epidemic, long-finned pilot whales and a white- beaked dolphin with DMV infection, and common dolphins with DMV infection.
Other gross lesions, which are often caused by secondary infections, include: enlargement, oedema and multifocal necrosis of the lymph nodes in long-finned pilot whales; multifocal erosive or ulcerative stomatitis of unknown aetiology in striped dolphins), pilot whales and common dolphins; similar lesions in the oesophagus, stomach, intestine or a combination of these in long- finned pilot whales, common dolphins and harbour porpoises; multifocal necrotizing encephalitis from Aspergillus sp. infection in striped dolphins; and suppurative bacterial balanoposthitis in Caspian seals and harbour porpoises.
The main organs in which microscopic lesions of mor- billivirus infection are observed in aquatic mammals are lymphoid organs, brain and lungs, although lesions containing characteristic inclusion bodies may frequently be found throughout the body. Although there is an overall similarity in these lesions, characteristic changes may be more pronounced in one morbillivirus- aquatic mammal combination than in another (Table 7.3). Another factor to consider is that the morbillivirus-associated lesions in lungs and lymphoid organs often are confounded by coinfections from lungworms and secondary infections from bacteria and fungi.
Lymphoid organs (spleen, lymph nodes, Peyer’s patches, thymus) have generally marked lymphoid depletion and lymphocytic necrosis. In long-finned pilot whales with DMV infection, however, the changes in the lymph nodes, considered to be the main microscopic lesion, have been described as multifocal necrotizing lymphadenitis1-60). In these pilot whales, as well as in striped dolphins, common dolphins and fin whales, syncytia have been observed in the remaining lymphoid tissue. Many histiocytes and some dendritic cells in lymphoid organs of Caspian seals have ICIB, and some syncytia in lymphoid organs of common dolphins and the fin whale have INIB. By immunohistochemistry for morbillivirus antigen, there is specific staining of lymphocytes, macrophages, dendritic cells, syncytia and (in common dolphins) fibroblasts.
Lungs have a multifocal broncho- interstitial pneumonia. Syncytia in alveolar and bronchiolar lumina are more
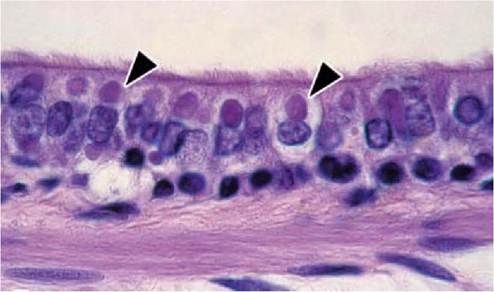
FIGURE 7.5 Acidophilic intracytoplasmic inclusions (arrowheads) in ciliated epithelial cells of a bronchiole from a Caspian seal with CDV infection. Haematoxylin and eosin. Reprinted with permission from Kuiken et al., 2006(48).
TABLE 7.3 Comparison of morbillivirus-specific microscopic lesions in aquatic mammals infected with morbillivirus.
Lesion Tissue Abundance of lesions per host—morbillivirus pair
| Harbour seal PDV (88,89,95) | Caspian seal CDV (48) | Striped dolphin DMV early (59) | Striped dolphin DMV late (91) | Pilot whale DMV (60) | White-beaked dolphin DMV (63) | Common dolphin DMV (64) | Harbour porpoise PMV (56,94) | ||
| Inflammation | Lymphoid | — | — | — | — | ++ | n.d. | — | — |
| Cerebrum | + | — | ++ | ++ | ++ | ++ | — | ++ | |
| Lung | ++ | + | + | — | + | + | + | ++ | |
| Other | — | — | — | — | — | — | — | + | |
| Syncytia | Lymphoid | — | — | + | — | + | n.d. | + | — |
| Cerebrum | + | — | + | — | + | — | — | — | |
| Lung | + | + | ++ | — | — | — | + | ++ | |
| Inclusions | Lymphoid | — | + | — | — | — | n.d. | + | — |
| Cerebrum | + | + | ++ | + | + | + | + | ++ | |
| Lung | + | ++ | ++ | — | — | — | + | ++ | |
| Other | + | ++ | + | — | — | n.d. | + | ++ | |
—, absent; +, present; ++, prominent; n.d., not done
prominent in striped dolphins and harbour porpoises than in harbour seals, Caspian seals and common dolphins. Inclusion bodies, both intracytoplasmic and intranuclear, have been observed in macrophages and syncytia in alveolar and bronchiolar lumina, and in epithelial cells lining the alveoli, bronchioles, bronchi and trachea. These inclusion bodies are abundant and prominent in bronchiolar epithelium of Caspian seals (Figure 7.5) and in syncytia in striped dolphins and harbour porpoises, but rarer in common dolphins and harbour seals. By immunohistochemistry for morbillivirus antigen, there is specific staining of the same cell types as those in which inclusion bodies are observed.
The brain, mainly cerebrum, has a non-suppurative encephalitis in many striped dolphins(91’93) and a white- beaked dolphin with DMV infection1-63), many harbour porpoises with PMV infection1-56’94), some harbour seals with PDV infection(92’95), but not in Caspian seals with CDV infection1-48). Characteristic changes in the grey matter are neuronal degeneration and necrosis, gliosis, perivascular cuffing and oedema. In the white matter, demyelination is observed in many harbour seals with PDV infection, some striped dolphins with DMV infection, but not in harbour porpoises with PMV infection. Acidophilic or amphophilic INIB and ICIB have been observed in neurons, astrocytes, microglia and ependymal cells. By immunohistochemistry for morbillivirus antigen, there is specific staining of generally the same cell types as those in which inclusion bodies are observed, as well as perivascular lymphocytes.
Besides lymphoid organs, lungs and brain, microscopic lesions of morbillivirus infection may occur in many other tissues. These lesions are characterized by the presence of acidophilic or amphophilic ICIB and (less often) INIB. These inclusion bodies have been described in epithelial cells of the urinary tract, digestive tract, bile duct, pancreatic collecting duct, eye, lacrimal gland, mammary gland
and skin. The presence of inclusion bodies may be associated with degeneration and necrosis of affected epithelial cells, as well as with inflammation from secondary bacterial or fungal infections.
Clinical signs
Clinical signs of morbillivirus disease are general malaise with or without involvement of the respiratory, digestive and reproductive tracts, central and peripheral nervous system including eyes, and skin. The clinical signs are partly caused by secondary disease from infection with facultative pathogens.
Most information about clinical signs of morbillivirus disease in pinnipeds comes from harbour seals, owing to detailed observation in rehabilitation centres. Harbour seals with PDV infection show clinical signs affecting the whole body (depression, lethargy, fever or hypothermia), central and peripheral nervous systems (abnormal posture, paresis, ataxia, seizures, head tremors and myoclonus of flipper or facial muscles), eyes (mucopurulent ocular discharge, redness of conjunctiva and sclera, corneal opacity, unilateral or bilateral blindness and nystagmus) (Figure 7.6), respiratory system (mucopurulent nasal discharge, coughing, tachypnea, dyspnea and subcutaneous emphysema), digestive system (diarrhoea), reproductive system (abortion) and skin (dermatitis)1-67’96). A subset of these

FIGURE 7.6 Mucopurulent discharge from eyes and nose of a harbour seal with PDV infection. Courtesy of J. Philippa.
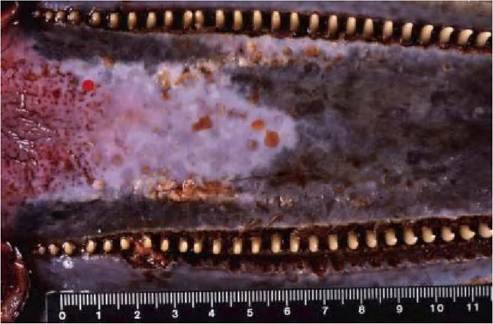
FIGURE 7.7 Multifocal erosive and ulcerative stomatitis in a striped dolphin with DMV infection. There are multiple erosions and vesicles in the hard palate, and linear ulceration in the gingiva around the teeth. Reprinted with permission from Domingo et al., 1992(93).
clinical signs has also been seen in Caspian seals with CDV infection1-48) and harp seals and hooded seals with PDV infection1-71). There is scant information about clinical signs of morbillivirus disease in cetaceans: most are found stranded dead or moribund. Striped dolphins with DMV infection can show clinical signs affecting the whole body (poor body condition leading to reduced flotation, crani- odorsal subcutaneous oedema, tachycardia, weak sound emission), nervous systems (disorientation, striking against rocks, no interest in swimming, muscle tremors), respiratory system (abnormal respiratory rates), digestive system (ulcers in the oral mucosa) (Figure 7.7) and skin (increased burdens of external parasites)1-61’67’93). Common dolphins with DMV infection can exhibit seizures, uncontrolled trembling and dyspnea.
There is no specific treatment for morbillivirus disease. Antibiotic treatment may be given against secondary bacterial infections.
DIAGNOSIS
The demonstration of acidophilic or amphophilic ICIB and INIB in typical tissues by microscopic examination is sufficiently specific to make the definitive diagnosis of morbillivirus disease.
The diagnosis of morbillivirus infection to genus may be confirmed by immunohistochemistry or serology. A two- fold increase in virus- neutralizing antibody (mainly IgG) titre in serial serum samples is diagnostic1-97), as well as a single positive IgM titre to morbillivirus by enzyme- linked immunosorbent assay (ELISA)(53). Both the virus neutralization (VN) test and ELISA may give false negatives, because not all infected animals mount an adequate immune response1-96).
The diagnosis of morbillivirus infection may be confirmed to species by PCR or virus isolation. Samples of choice are nasal and pharyngeal swabs from live animals and brain, lung, tracheo- bronchial lymph node, spleen, liver and kidney from carcasses. Brain samples should not be omitted because infection may be limited to this tissue(89). Besides standard cell cultures, Vero cells expressing dog-SLAM may be used(98). Morbilliviruses grow faster and to a higher titre on Vero-SLAM cells, and isolation of field viruses is more sensitive in these cells.
Population screening for morbillivirus infection is often performed on live animals by serology, and on dead stranded animals by PCR or immunohistochemistry on tissues. Blood samples from dead stranded animals also can be used for serologic screening, but because of haemolysis the more specific VN is then precluded and results are often not reproducible.
MANAGEMENT, CONTROL
AND REGULATIONS
Management of morbillivirus infections in aquatic mammals depends on the susceptibility and conservation status of the host species, and on the virological and immunological status of the host population. In free-living aquatic mammals, population management needs to account for potential reduction of population size from recurrent epidemics and for risk of virus transmission between separate populations. Such virus transmission can occur directly through translocation of individual animals and indirectly through fishing or other human activities that influence the home range and migratory patterns of aquatic mammal populations. Vaccination by injection is impractical for free-living aquatic mammals. It is only considered when species are highly endangered, for example, the endangered Mediterranean monk seal(99).
I n captive aquatic mammals, it is important to minimize the risk of virus introduction from both terrestrial and aquatic mammals by restricting access of other species and by following appropriate hygienic and quarantine procedures. Captive aquatic mammals can be vaccinated with an inactivated vaccine against the homologous or heterologous virus. Harbour seals brought into rehabilitation centres during PDV epidemics have been successfully vaccinated with inactivated CDV vaccines, including commercially available vaccines1-100). Recently, DNA vaccines have been developed1-101). The use of live attenuated vaccines is contra-indicated because of the risk of disease to the vaccinated animal and because of the risk of virus spread in the population.
CDV, PDV, DMV, PMV and PWMV infections are not notifiable, but the wildlife group of the World Organisation for Animal Health (OIE) does request their reporting. The morbillivirus OIE reference laboratory is the Institute for Animal Health, Pirbright Laboratory.
PUBLIC HEALTH CONCERN
The primary public health concern from morbillivirus infections in aquatic mammals is not from the virus itself but from secondary infections that are exacerbated because of the morbillivirus-induced immunosuppression. For example, morbillivirus-infected aquatic mammals have been diagnosed with toxoplasmosis(93), systemic Streptoc- cocus spp., Staphyloccus aureus or Escherichia coli infec- tions(88,89), which pose a risk to people handling such animals.
CDV, PDV, DMV, PMV and PWMV infections are currently not considered to be zoonoses. However, as for CDV, it cannot be ruled out that in the event of eradication of measles, humans could become a new ecological niche for a morbillivirus species from aquatic mammals(102).
SIGNIFICANCE AND IMPLICATIONS FOR ANIMAL HEALTH
Some morbilliviruses from aquatic mammals present a threat to domestic animals. Although DMV and PMV did not cause clinical disease in experimentally infected domestic dogs and ruminants(57), PDV did cause clinical disease in experimentally infected domestic dogs and experimentally or naturally infected mustelids(103’104).
Morbillivirus infections present a serious threat to endangered aquatic mammals, e.g. the Mediterranean monk seal. The high mortality they cause may reduce numbers to a level that results in extinction. Morbillivirus infections may also cause high mortality in less endangered aquatic mammals and need to be accounted for in their population management.