SAPROPHYTIC FUNGI
Systemic Diseases
ASPERGILLUS SPP. (ASPERGILLOSIS)
CAUSATIVE AGENT (CLASSIFICATION, morphology) The genus Aspergillus is a member of the Family Trichocomaceae, Order Eurotiales, Class Ascomycetes (App.
1: Table 6) (Howard 2003). This genus includes at least 185 species, of which 20 are known to be pathogenic to plants or animals (Converse 2007) The most common animal pathogens are A. fumiga- tus and A. niger, which cause aspergillosis, and A. flavus, which produces an aflatoxin (Chute et al. 1965). Aspergillus fumigatus is the most commonly reported species and is most pathogenic for both wild birds (Converse 2007) and mammals (Burek 2001). Pathogenic members of the genus Aspergillus include anamorphs among at least seven teleomorph genera: Emericella, Eurotium, Fennellia, Hemicarpenteles, Neopetromyces, Neosartorya, and Petromyces (Howard 2003). Because species of Aspergillus often are not clearly identified, the genus will be addressed as a whole.host range and geographic distribution Aspergillus spp. have a worldwide distribution (Burek 2001) and commonly are found among free-living birds (McDiarmid 1955, Beer 1963, Rosen 1964, O'Meara and Witter 1971) and mammals (Burek 2001). In North America, infection by Aspergillus spp. are considered common in waterfowl, gulls, and corvids; occasional in songbirds, upland game birds, and blackbirds; and infrequent or rare in wild raptors, herons, and shorebirds (Friend 1999a). On a worldwide basis, waterfowl, raptors, and gulls represent the majority of free-living hosts reported with aspergillosis (Converse 2007). Although less frequently reported in mammals, Aspergillus spp. have occurred in farmed deer (Odocoileus spp.) (Jensen et al. 1989), alpacas (Vicugna pacos) (Pickett et al. 1985, Severo et al. 1989), camels (Camelus dromedarius) (El-Khouly et al. 1992), dolphins (Tursiops truncatus) (Reidarson et al.
1998b), and wild felids (Peden et al. 1985). It also is reported in domestic birds and mammals (Fraser and Mays 1986, Acha and Szyfres 2001).LIFE CYCLES AND VARIATIONS Aspergillus spp. reproduce asexually by forming aerial fruiting bodies that bear conidiospores, asexual spores formed on the tip of specialized hyphae called conidiophores; these spores are easily inhaled into lungs and air sacs (Converse 2007). Some Aspergillus spp. produce mycotoxins (aflatoxins) (Converse 2007); this topic is addressed later in this chapter.
reservoirs and transmission Aspergillus spp. generally are ubiquitous saprophytes found in soil, decaying vegetation, and agricultural wastes such as spilled grain and corn silage (O'Meara and Witter 1971, Converse 2007). Aspergillus spp. generally grow over a wide temperature range (18 to 30°C); most will spor- ulate at 23 to 26°C (Raper and Fennell 1965). Growth and sporulation of Aspergillus spp. are encouraged by moist conditions and decaying vegetation (Burek 2001). Spores commonly are released when fungal fruiting bodies are broken by animal movements (Bellrose 1945).
Primary transmission is through inhalation, which typically requires one to several million air-borne spores (Austwick 1968, Atasever and Gumussoy 2004). Disturbance of infected soil and vegetation results in aerosols, allowing for aspiration and deposition of spores in bronchioles and pulmonary alveoli (Burek 2001). Transmission also can occur by puncture wounds contaminated with hyphae or spores, especially if the lungs or air sacs are penetrated (O'Meara and Witter 1971). Aspergillus spp. spores can penetrate intact egg shells, resulting in mortality of the embryos or birds hatched (O'Meara and Chute 1959). Ingestion is an unlikely route of transmission (Chute et al. 1965).
CLINICAL EFFECTS AND IDENTIFICATION Aspergillosis is a noncontagious disease usually caused by inhalation of Aspergillus sp. spores (Converse 2007). Most reports are among birds. Two hypotheses for the development of avian aspergillosis are that the initial dose of spores is very high, exceeding the natural resistance of a host, or that birds may carry the fungi but become ill after stresses lower the immune response (Friend and Trainer 1969).
Both acute and chronic forms of aspergillosis occur (Converse 2007). In acute cases, birds die in a few days from severe respiratory compromise following a generalized lung infection; birds generally are in good body condition with no other evident problems (O'Meara and Witter 1971, Redig et al. 1980).
With chronic aspergillosis, birds develop a slowly progressing disease and may become severely debilitated before dying. Signs often include emaciation, reduced activity, inability to fly, dyspnea, and diarrhea (Converse 2007). Characteristic lesions include firm nodules and sometimes colored conidiospores in the respiratory tract (O'Meara and Witter 1971, Redig et al. 1980). There typically is a strong inflammatory response with many nodules disseminated through the respiratory tract; in time any internal organ can be infected via the vascular system. Birds with chronic aspergillosis also may have evidence of trauma, parasitism, and other diseases, as well as a history of capture, captivity, or rehabilitation (Converse 2007). Avian aspergillosis losses often are associated with changing environmental conditions, such as increases in temperature and moisture, which can enhance development of fungi on waste grain in agricultural fields (Converse 2007).
Among mammals, clinical signs and pathology vary considerably by the organ system involved (Burek 2001). With a primary respiratory disease, there are miliary nodules in the lungs containing fungal hyphae; in disseminated disease, nodules may be scattered throughout the body. The fungi can cause considerable tissue damage associated with vascular invasion and thrombosis; the hyphae proliferate in necrotic tissue (Burek 2001).
Antemortem (before death) diagnosis is based on clinical signs, serological tests, culture of tracheal washes, endoscopy, and radiographs (Converse 2007). Diagnostic tests include an enzyme-linked immunosorbent assay (ELISA) (Converse 2007) and polymerase chain reaction (PCR) (Katz et al.
1996). Adult free-ranging female birds can be tested indirectly by testing for IgG antibodies in their eggs against Aspergillus spp. (Graczyk and Cranfield 1995); titers in eggs are correlated with specific antibody levels in the mothers (Graczyk and Cranfield 1996). Because Aspergillus spp. are common contaminants, positive culture should correlate with appropriate histopathologic evidence of tissue invasion (Chandler and Watts 1987).Postmortem diagnosis of aspergillosis is based on the presence of compatible lesions at necropsy and identification of Aspergillus spp. in tissues by isolation or identification of characteristic fungal hyphae (Converse 2007). The fungi can be grown on several media (Kunkle and Richard 1998). However, the fungal hyphae need to be distinguished from other invasive fungi such as Fusarium spp. (Chandler and Watts 1987). Monoclonal and polyclonal antibodies can be used with an immunohistochemical label to identify Aspergillus spp. (Carrasco et al. 1993).
population effects Epizootics of acute aspergillosis have been reported among free- living waterfowl (Bowes 1990), including mallards (Anas platyrhynchos) (Neff 1955, Adrian et al. 1978), as well as common crows (Corvus brachyrhynchos) (Zinkl et al. 1977). Mortality may disproportionately affect younger animals (Brand et al. 1988), and may be influenced by severity and duration of exposure, and stress from concurrent infections of other parasites (O'Meara and Witter 1971). Chronic aspergillosis is associated with individual bird deaths rather than group mortality (Converse 2007).
Aspergillosis was the most common (23%) finding among dead or moribund common loons (Gavia immer) in New York from 1972 to 1999, with ingestion of lead fishing weights being the second most common diagnosis (21%) (Stone and Okoniewski 2001). This prevalence of lead may be significant as aspergillosis has been associated with lead poisoning in other areas (Locke et al. 1969, Bair et al. 1988, Souza and Degernes 2005).
Aspergillosis also has been associated with tracheal trematodes (Pennycott 1999) and thiamine deficiency (Friend and Trainer 1969).Aspergillosis has caused mortality in a number of marine mammals (Sweeney et al. 1976). Pneumonia caused by Aspergillus spp. may account for most pulmonary mycoses in marine mammals (Reidarson et al. 1998b).
special problems No specific persistent problems are reported.
control Immunity to Aspergillus spp. can develop among birds. Adult birds are more resistant than chicks to aspergillosis; there is evidence for innate resistance, but little definitive data are available (Converse 2007). Birds are able to produce both IgM and IgG antibodies to Aspergillus spp. (Martinez-Quesada et al. 1993), and mammals produce IgE (Kwon-Chung and Bennett 1992), but the role of antibodies in protecting infected animals or enhancing recovery is not clear (Converse 2007). There is evidence among birds that maternal antibodies are protective for hatchlings (Graczyk and Cranfield 1995).
Among mammals, itraconazole can be used for treatment of systemic aspergillosis (Denning and Stevens 1990), but cure is uncommon (Burek 2001). Decreasing the numbers of spores contaminating an environment and controlling other predisposing conditions also are considered important (Pier and Richard 1992).
COCCIDIOIDES IMMITIS AND C. POSADASII (coccidioidomycosis)
causative agent (classification, morphology) Coccidioides spp. are ascomycetes found in the Family Onygenaceae, Order
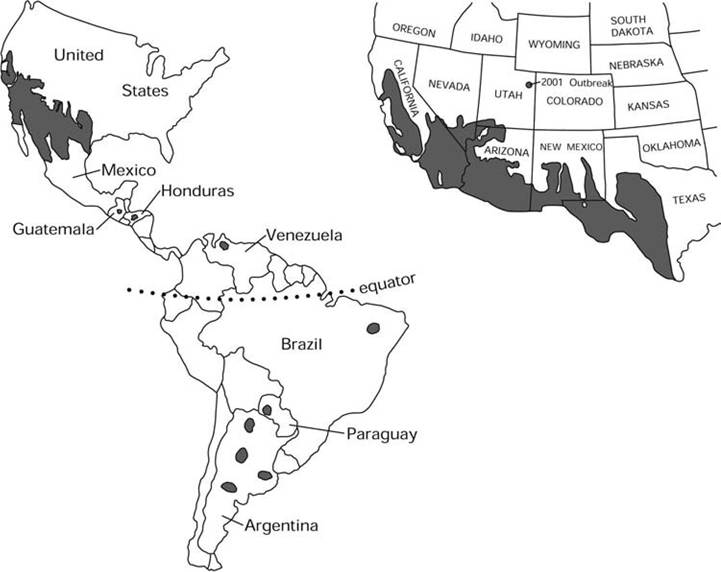
MAP 7.1. Distribution of endemic Coccidioides immitis and C. posadasii (Bultman et al. 2004; courtesy of M. W. Bultman and the U.S. Geological Service).
Onygenales (App. 1: Table 6) (Howard 2003, Clemons et al. 2007). Until recently only one species, C. immitis, was recognized. However, there now is phylogenetic evidence for a second species, C. posadasii (Fisher et al. 2002, Tintelnot et al.
2007). The teleomorph form is not certain (Saubolle et al. 2007), but Uncinocarpus spp. has been proposed (Sigler 2003).HOST RANGE AND GEOGRAPHIC DISTRIBUTION Coccidioides spp. can infect many mammal species (Burek 2001, Laniado-Laborin 2007) and rarely reptiles (Timm et al. 1988). No reports have yet been made of avian infections (Laniado-Laborin 2007). Most reports from wild species are from captive animals in zoos or exhibits in enzootic regions; death from coccidioidomycosis has been reported from a variety of nondomestic canids and felids, as well as bats, wallabies, kangaroos, tapirs, and several primates (Shubitz 2007). Among free-living wildlife, Coccidioides spp. has been reported from desert bighorns (0vis canadensis nelsoni) (Jessup et al. 1989), mountain lions (Puma concolor) (Clyde et al. 1990, Adaska 1999), and coyotes (Canis latrans) (Straub et al. 1961). Coccidioidomycosis also has been reported from free-living marine mammals, including California sea lions (Zalophus californianus) (Fauquier et al. 1996), a bottle-nosed dolphin (Tursiops truncatus) (Reidarson et al. 1998a), and a sea otter (Enhydra lutris) (Cornell et al. 1979). Among domestic mammals, lesions containing Coccidioides spp. have been found in cattle, sheep, and swine, but these species rarely develop overt disease (Maddy 1959).
Coccidioides immitis and C. posadasii are endemic in the Western Hemisphere, almost exclusively between 40°N and 40°S latitudes (Burek 2001) in the Lower Sonoran Life Zone (Map 7.1); these areas seldom have freezing temperatures during the year, and the climate is arid, with yearly rainfall ranging from 11 to 44 cm (Galgiani i993)∙ Coccidioides spp. survives well in saline soil (Fauquier et al. 1996), and elevated salinity may inhibit some microbial competitors (Egeberg et al. 1964). Presence of fine sand and silt in the soil is the single characteristic common in all areas in which the organism is found (Saubolle et al. 2007). In enzootic areas, the mean annual soil temperatures of enzootic regions range from 15°C to over 22°C; the soil is alkaline, with adequate pore space in the upper 20 cm for moisture, oxygen, and room for growth.
Endemic foci have been identified in semi- arid areas of the United States and Mexico, as well as some areas of Central and South America (Laniado-Laborin 2007). There is evidence that Coccidioides spp. may have had a larger range at one time in North America, or that migratory animals from other regions were infected as they passed through enzootic areas (Morrow 2006). With the division of species, C. immitis now is geographically restricted to the Central Valley of California and southern California; in contrast, C. posadasii occurs in all other regions of North, Central, and South America where infections have been reported (Fisher et al. 2002).
Both species appear to be have considerable genetic diversity with frequent genetic recombination; both species also have biogeographically distinct populations (Barker et al. 2007). The genetic diversity of Coccidioides spp. in North America has clear geographic partitions, whereas the populations of Coccidioides spp. in South America are genetically similar and appear to be founded on a North American population centered in Texas; one hypothesis is that South America was colonized by Amerindians carrying this fungus (Fisher et al. 2001).
LIFE CYCLES AND VARIATIONS The life cycle of Coccidioides spp. is unique among pathogenic fungi (Kirkland and Fierer 1996). These dimorphic fungi are considered true pathogens and readily change from a saprophytic mycelial form in soil into a round, thick-walled spher- ule/endospore form in an animal host (Shubitz 2007). In soil, the fungus grows on decaying organic matter and is characterized by mycelia with branching segmented hyphae. Here the fungus reproduces asexually by disarticulating the hyphae into small, environmentally resistant structures called arthrospores, also called arthroconidia; the arthroconidia are released from mycelia and dispersed in the air (Hector and Laniado-Laborin 2005). The saprophytic mycelial cycle does not depend on a reversion to the parasitic form as it contains large amounts of arthroconidia that are dispersed by the wind to colonize new sites. However, upon inhalation by a host, the conidia are converted by isotropic growth into spherules (Cole and Hung 2001). Each spherule divides to produce hundreds of endospores, which then disperse into surrounding tissue and produce a second generation of spherules (San-Blas and Nino- Vega 2004). The endospores again can revert to the saprophytic mycelial stage if they reach the soil through death of the infected animal or by body secretions (Bultman et al. 2004).
Reservoirs and transmission The reservoirs are the soils and infected animals of the Lower Sonoran Life Zone. Coccidioides spp. spores present in dry soil first may require moisture to germinate and grow, and then require a dry period for the fungal hyphae to desiccate and form arthroconidia (Kolivras and Comrie 2003). Cases of coccidioidomycosis are linked to both the amount of dust in the air and the cumulative rainfall over the preceding several months (Park et al. 2005). In one study, precipitation during the normally arid periods in late spring and early summer 1.5 to 2 years before the season of exposure was the dominant predictor of coccidioidomycosis cases for all seasons (Comrie 2005). Work is continuing to clarify the environmental factors leading to the success of Coccidioides spp. throughout its range (Baptista-Rosas et al. 2007, Fisher et al. 2007).
Inhalation of arthrospores is the principal means of transmission (Davis 1981); in the mouse, only a few arthrospores administered intranasally are required for infection (Galgiani 1993). The arthroconidia developing in the Coccidioides spp. hyphae commonly are separated by empty, thin-walled brittle cells (disjunctors) formed by autolysis. The spores are easily released into the air by soil disruption and wind (Saubolle et al. 2007), and spread of the fungus by wind is an important local process (Bultman et al. 2004). However, there are distinct genetic clades of Coccidioides spp.; thus arthroconidia probably do not travel over long distances to new sites (Fisher et al. 2001).
CLINICAL EFFECTS AND IDENTIFICATION Coccidioidomycosis, the disease resulting from Coccidioides spp. infection, is not to be confused with the disease “coccidiosis,” caused by members of the sporozoan protozoa (App. 1: Table 5) (Galgiani 1999). Virulence of Coccidioides spp. is a function of a spherule outer wall glycoprotein that reduces cell-mediated immunity, reduces glycoprotein occurrence on endospores to prevent host recognition, and produces of chemicals contributing to tissue damage at the sites of infection (Hung et al. 2007). Clinical signs vary greatly, ranging from a benign, selflimited upper respiratory infection, to chronic pulmonary disease, to disseminated fatal disease (Burek 2001).
In humans, symptoms of coccidioidomycosis usually begin 7 to 21 days after inhalation of arthroconidia and may include fever, cough, chest discomfort, malaise, and fatigue (Saubolle et al. 2007). Symptoms of pneumonia generally last under 3 weeks, but can be longer. Skin rashes can be seen in 10 to 50% of infected humans. Disseminated coccidioidomycosis occurs in less than 5% of symptomatic patients; those of black or Filipino ethnic backgrounds, pregnant women, and immunocompromised patients have higher risks for disseminated disease (Saubolle et al. 2007).
Detection and diagnosis in animals, including humans, often is based on direct microscopy, culture, and serologic findings (Saubolle 2007, Parish and Blair 2008). Fungal identification can be based on phenotypic characters such as the presence of spherules in hosts, the presence of arthroconidia in culture, or genotypic characteristics (Saubolle 2007). Enzyme immunoassays and immunodiffusion methods commonly are used for detection of both IgM and IgG antibodies (Saubolle 2007). Polymerase chain reaction tests (PCR) and restriction fragment length polymorphism (RFLP) have been used to identify and differentiate Coccidioides spp. (Umeyama et al. 2006, Tintelnot et al. 2007).
Detection of C. immitis in the soil is difficult. Traditionally, mice have been inoculated with isolates from suspect soil and their organs later evaluated for evidence of the unique spherules characteristic of the parasitic form of the fungus (Bultman et al. 2004). Although DNA analysis is used, there are no standardized procedures as yet (Bultman et al. 2004).
population effects There are no known ongoing significant population impacts among wildlife.
special problems Coccidioidomycosis is a common infectious disease among humans in the southwestern United States, Mexico, Central America, and South America (DiCaudo 2006), with upwards of 100,000 primary coc- cidial infections of humans each year in the United States alone. While recent increases in human cases in Arizona were thought to be linked to development of a hypervirulent strain, this does not appear to be the case (Jewell et al. 2008). About 3,000 inmates were required to be moved out of the Central Valley of California to other prisons in 2013 to reduce the risk of coccidioidomycosis among high-risk prisoners (http://articles.latimes.com/2013/apr/30/local/ la-me-prisons-valley-fever-20130430).
The occurrence of coccidioidomycosis in marine mammals is intriguing but most likely is the result of marine mammals inhaling arthroconidia blown from land over the water (Shubitz 2007). Coccidioides spp. can survive for an extended period in seawater (Fauquier et al. 1996).
control Macrophages producing cytokines against spherules are one important host defense (Viriyakosol et al. 2005). Humoral antibodies generally are not considered protective against Coccidioides spp. (Saubolle et al. 2007); however, B-lymphocytes may play a role in vaccine-induced immunity (Magee et al. 2005).
Orally administered azole and other antifungal agents are the primary therapy used for captive animals (Shubitz 2007). Among humans, coccidioidomycosis usually is treated with azole or amphotericin B (Parish and Blair 2008).
Recovery from coccidioidal infection confers lifelong protection against reinfection (Smith and Pappagianis 1961, Pappagianis 2001), and there is considerable interest in developing effective vaccines, especially for humans (Johnson et al. 2007). A vaccine based on whole killed spherules is effective for some nonhumans (Levine et al. 1965).
Other Saprophytic Systemic Fungi Occasionally Affecting Wildlife
EMMONSlA spp. (CH RYSOSPO Rl UM PARVUM) Emmonsia crescens and E. parva are ascomyce- tes (Family Onygenaceae, Order Onygenales) (App. 1: Table 6) that infect small mammals. These parasites also have been classified as subspecies of Chrysosporium parvum (Burek 2001), but currently are identified as two distinct species of Emmonsia (Sigler 2003). Both species are dimorphic fungi that live in the soil and produce self-limiting pulmonary mycotic infections in a wide variety of small mammals that burrow or produce ground nests; they also infect mustelids (Dvorek et al. 1973, Jellison 1981). They appear to have a worldwide distribution (Jellison and Lord 1964, Dvorek et al. 1973, Jellison 1981, Hubalek et al. 2005), with the exception of Australia and Antarctica (Emmons and Jellison 1960).
Transmission is primarily through inhalation of spores, called adiaspores, from the free- living soil forms (Burek 2001). The predatorprey relation may help in the spread of the fungi since adiaspores passed in the feces are viable and germinate (Krivanec and Otcenasek 1977).
The fungi are associated with mammalian species using forest-grassland ecotones (Leighton and Wobeser 1978), especially near windbreaks (Hubalek et al. 1991, Hubalek et al. 1998), but can occur in other uncultivated soils (Fischer 2001). Peak fungal proliferation occurs when mean monthly soil temperatures 5 cm below the surface range between 3 and 12°C (Hubalek et al. 1993).
In mammalian tissues, the adiaspores develop into thick-walled, gray-white fungal spherules which are surrounded by granulomatous infiltrates; the disease is called adiaspiromycosis. There appears to be no multiplication of the fungus in host species, even though extensive infections have been observed in small mammals (Burek 2001). Lesions usually are restricted to lung inflammations. In heavy infections there may be some pulmonary compromise (Chandler and Watts 1987). No control methods are proposed.
Histoplasma capsulatum Elistoplasma capsulatum, the cause of histoplasmosis, is an ascomycete (Family Onygenaceae, Order Onygenales) (App. 1: Table 6); this dimorphic fungus develops hyphae in soil and a yeast stage in host tissues (Burek 2001). This parasite has a worldwide distribution and is found in both temperate and tropical zones (Sanger 1981). Only mammals are known to be naturally infected; of these, rodents and bats are most important (Otcenasek et al. 1967). Currently three subspecies are recognized (McGinnis et al. 1999), of which H. capsulatum var. capsulatum is of greatest significance and is the focus of this summary.
The soil is the primary natural habitat and source of infection of H. capsulatum (Otcenasek et al. 1967). Interestingly, the fungi often are associated with soils enriched by bird feces (Ajello 1964) or with bat guano (DiSalvo et al. 1970). Despite past justification of histoplasmosis as a basis for instituting “blackbird control” programs (Anonymous 1975), birds are not able to sustain infections of H. capsulatum, likely because of their high (42°C) body temperatures (Ajello 1967); overall, there is no evidence that birds regularly carry the fungus or can seed it into the soil (Sanger 1981).
Bats also are susceptible to histoplasmosis (Ajello 1967), and the frequent involvement of the intestinal tract in bats is a mechanism for seeding the environment with the fungus (Shacklette et al. 1967, Greer and McMurray 1981). In Mexico, H. capsulatum has been found primarily in caves, abandoned mines, and houses with bat guano (Velasco-Castrejon and Gonzalez-Ochoa 1977). Further, the fungi can grow in bat guano; thus bats can excrete H. capsulatum in a medium in which the fungus can propagate itself (Isbister et al. 1976). It is likely that bats are the animals of greatest significance in the epizootiology of the disease, but other factors also are important (Ajello 1967), including temperatures under 40°C and soil pH of 5 to 10 (Goodman 1965). Histoplasma capsulatum also may parasitize and grow in soil amebae; parasitism of soil amebae may select for features that increase pathogenicity of H. capsulatum for mammals (Steenbergen et al. 2004).
Transmission to susceptible hosts occurs by inhalation of spores from soil (Otcenasek et al. 1967), and rarely by ingestion or skin contact with infective spores of H. capsulatum (Burek 2001). Bat caves can have high concentrations of H. capsulatum in their atmosphere (Shacklette et al. 1967). Foci of infection occur in the lungs and regional lymph nodes. In disseminated histoplasmosis, many tissues are affected, including lymph nodes, spleen, bone marrow, liver, and adrenals (Burek 2001). Many infected animals on enzootic sites have clinically inapparent infections (Burek 2001); various stresses have been implicated as contributing factors for immunosuppression and emergence of overt disease in wildlife (Quandt and Nesbit 1992).
Diagnosis is based on finding organisms in monocytes and macrophages of blood smears, bone marrow aspirates, or biopsy samples of lymph nodes, liver, intestine, or rectum (Burek 2001). Thoracic radiographs, a Histoplasma skin test, and serologic methods can be used (Quandt and Nesbit 1992), as well as ELISA and radioimmunoassay (Durkin et al. 1997). Restriction fragment length polymorphisms and polymerase chain reaction (PCR)-based DNA fingerprinting also have been successful tools to identify and characterize strains (Kersulyte et al. 1992, Bialek et al. 2002).
There are no control methods for histoplasmosis in wildlife. However, ketoconazole has been used with domestic pets (Legendre 1995), and several other antibiotics appear to have promise (Li et al. 2000).
Subcutaneous Diseases
A number of fungi cause subcutaneous infections among mammals (Burek 2001), including humans (Schell et al. 1999), and birds (Friend 1999b). Until recently, most of these fungi were classified among the Fungi Imperfecti, but the relationship for many of them to other well- established groups is becoming clearer (App. 1: Table 6). Two species causing some pathology among wildlife are Sporothrix schenckii and Lacazia (Loboa) loboi.
Sporothrix schenckii Sporotrichosis is a subcutaneous mycosis caused by Sporothrix schenckii (Order Ophiostomatales, Class Asco- mycetes), a thermally dimorphic fungus that forms mycelia in decaying vegetation and fungal media at 25 to 30°C, but yeast-like stages in tissues and fungal media at 37°C (Schell et al. 1999). The parasite has a worldwide distribution, but is more common in temperate and tropical climates (Lopes-Bezerra et al. 2006). Based on mitochondrial DNA analysis using restriction fragment length polymorphism patterns, there are 24 different genotypes divided into two main types (Mesa-Arango et al. 2002). Type A is found predominantly in North and South America and South Africa (Ishizaki et al. 1998, 2000); Type B is found primarily is Spain, Asia, and Australia (Ishizaki et al. 2000, Mesa-Arango et al. 2002).
Sporothrix schenckii occurs in both marine (Migaki et al. 1978) and terrestrial mammals (Jones and Hunt 1983, Costa et al. 1994, Werner and Werner 1994). Though it has also been isolated from birds (Ziolkowska and Tokarzewski 2007), S. schenckii is not known
to cause pathology or mortality among birds. This fungus typically is inoculated through a scratch or into a previous wound by plant material, particularly of a grassy or straw-like nature (Mackinnon et al. 1969, Windingstad 1990).
In its most common form, lymphocutane- ous lesions are characterized by small, firm nodules on the head, extremities, and tail that ulcerate and discharge hemorrhagic or serous exudate (Burek 2001). Diagnosis is based on clinical signs, histopathology, fungal cultures, and serologic tests (Goad and Pecquet-Goad 1986, Hu et al. 2003, Bernardes-Engemann et al. 2005). In captive hosts, the infection can be successfully treated with potassium iodide (Scott et al. 1995) or several antibiotics such as itraconazole and ketoconazole (Legendre 1995).
LACAz∣A (LOBOA) loboi Lacazia (Loboa) loboi is an ascomycete (Family Onygenaceae, Order Onygenales) (Herr et al. 2001) that is closely related to the fungal parasite Paracoccidioides brasiliensis (Vilela et al. 2005). Lacazia loboi causes subcutaneous mycoses in bottle-nosed dolphins (Tursiops truncatus) (Migaki et al. 1971, Caldwell et al. 1975, Simose-Lopes et al. 1993) and the Guiana dolphin (Sotaliafluviatilis guianensis) (DeVries and Laarmann 1973), as well as humans (Burns et al. 2000, Opromolla et al. 2000). It is considered enzootic to the South and Central American tropics (Pfaller and Diekema 2005). Natural infections have been reported only in dolphins and humans, but experimental infections occur in hamsters (Opromolla and Nogueira 2000) and armadillos (Euphractus sexcinctus) (Sampaio and Braga-Dias 1977).
There may be both an aquatic and a terrestrial reservoir. The occurrence of lobomycosis in dolphins from marine and freshwater habitats of Florida (Reif et al. 2006), Texas (Cowan 1993), the Spanish-French coast (Symmers 1983), the south Brazilian coast (Simose-Lopes et al. 1993), and the Surinam River Estuary of South America (DeVries and Laarmann 1973) is evidence that an aquatic reservoir is important. Lacazia loboi also may be a saprophyte of soil or vegetation (Pfaller and Diekema 2005). Based on consistent differences in fungal cell size between dolphins and humans, there may be strain differences between the human (terrestrial) and dolphin (aquatic) strains (Haubold et al. 2000). Cutaneous trauma is the likely mode of infection (Pfaller and Diekema 2005). There is one possible case of dolphin-to-human transmission (Borelli 1961).
In dolphins and humans, the disease is characterized by slowly developing cutaneous nodules and hard plaques arising on injured areas of skin (Caldwell et al. 1975, Simose- Lopes et al. 1993, Pfaller and Diekema 2005). Diagnosis is based on finding the characteristic yeast cells in lesion exudates or tissue sections (Burns et al. 2000); a vinyl adhesive tape method has proved simple and useful for diagnosis (Miranda and Silva 2005). Western blotting analysis is used to detect dolphin and human antibodies against L. loboi, and may be useful for further epizootiological studies on the parasite (Mendoza et al. 2008).
Although lobomycosis generally has been reported as isolated observations among dolphins, there is evidence for a developing epizootic among bottle-nosed dolphins in a Florida lagoon. Environmental stress caused by freshwater intrusions are speculated to have contributed to the high (30%) prevalence of infections observed (Reif et al. 2006).
Several antifungal agents have been used in humans and other animals (Brun 1999). Generally, treatment of free-ranging animals is not likely or warranted (Burek 2001).