APOPTOSIS IN HIV-1 INFECTED ORGAN CULTURE
The thymus is the primary lymphoid organ where T cells are generated during fetal and neonatal development. In adults, however, peripheral proliferation of T cells is sufficient to maintain normal immune homeostasis.
Nevertheless, the thymus functions throughout life, although at reduced levels, and responds to T cell depletion induced by HIV-1.1’2 Clinical studies of HIV-1-infected children and adults suggest that HIV-1 infection causes thymic dysfunction, increased involution, and thymocyte depletion.3’4 HIV-1 infection of the thymus, particularly of infants, is documented and is frequently associated with rapid progression to AIDS.5-8 Reduced thymic volume, impaired delayed-type hypersensitive responses, and spontaneous apoptosis of thymocytes are associated with HIV-1 infection.9’10 Alteration of thymocyte maturation by HIV-1 could have an impact on the composition and function of peripheral T cell populations and may interfere with immune responses.11 Recent data on thymic function during highly active antiretroviral therapy (HAART) suggest that thymic recovery may be achieved in some patients on HAART.5’12’13 Extensive thymic structural damage, however, may hamper immune reconstitution, especially in pediatric patients for whom the thymus is crucial for generating naive T cells. The mechanisms by which HIV-1 causes depletion of thymocytes and destruction of the thymus are not well understood. Thymocytes develop in a specialized microenvironment that is required for their normal maturation and selection. As the thymus is difficult to study in humans, several model systems of human thymopoiesis were developed, including fetal thymic organ culture (FTOC) and the severe combined immune deficient (SCID) mouse implanted with human fetal thymus and liver tissue (SCID-hu Thy/Liv mouse). HIV-1 can infect, replicate, and cause depletion of thymocytes in these experimental models.14-18Fetal Thymic Organ Culture (FTOC)
Dissecting human thymus into small pieces that contain several intact lobules creates FTOC. Tissue pieces are infected with HIV-1 and cultured on sterile membrane filters with or without collagen rafts in enriched media. Several groups have used human FTOC as a model to study HIV-1 infection and pathogenesis.15’19-26 FTOC can be infected with both CCR5-tropic HIV-1 (R5 HIV-1) and CXCR4-tropic HIV-1 (X4 HIV-1) and exhibits pathology similar to that observed in the SCID-hu Thy/Liv model.15’21’25 Both R5 HIV-1 and X4 HIV-1 deplete thymocytes in a time-dependent manner; however, X4 HIV-1 exhibits greater cytopathic effect (CPE) than R5 HIV-1 in FTOC (Figure 18.1A).15’25 Many cytokines that are induced in HIV-1 infected individuals27-30 are also induced by HIV-1 infection of FTOC. These include interleukin (IL)-10, TGF-β, IFN-α, CXCL10, and Oncostatin m.23’25’31’32 FTOC was shown to be responsive to several antiviral agents15 and, therefore, is a useful thymus model for the study of HIV-1 replication and its inhibition.
The mechanisms by which HIV-1 infection causes thymic damage are not well understood. Nevertheless, apoptosis is clearly the ultimate cause of CD4+ thymocyte depletion during HIV-1 infection of FTOC.15 The nature of the signals that initiate apoptosis in the HIV-1-infected thymus, however, is not known. R5 HIV-1 clones preferentially deplete the CCR5+ subset of CD4+ thymocytes, whereas X4 HIV-1 clones spare this thymocyte subset, indicating that direct killing of infected cells is likely the predominant mechanism of thymocyte depletion.25 HIV-1 can infect thymocytes at several different stages of maturation, which may contribute to direct CPE.33,34 Alternatively, changes in the thymic milieu due to HIV-1 infection may induce apoptosis of uninfected thymocytes.14’34 HIV-1 infection of thymocytes impedes thymocyte maturation at early stages, including CD3-, CD4-, CD8-, CD25-, and CD44+.11 HIV-1 nef gene expression in thymic precursor cells and their colonization in FTOC and SCID-hu also lead to impaired T cell develop- ment.32’35 Progenitor cells derived from infants born to HIV-positive mothers have decreased cloning efficiency and generate fewer T cells in FTOC.
This impaired progenitor cell function may be responsible, at least in part, for lower naive CD4 counts and reduced thymic output in HIV-negative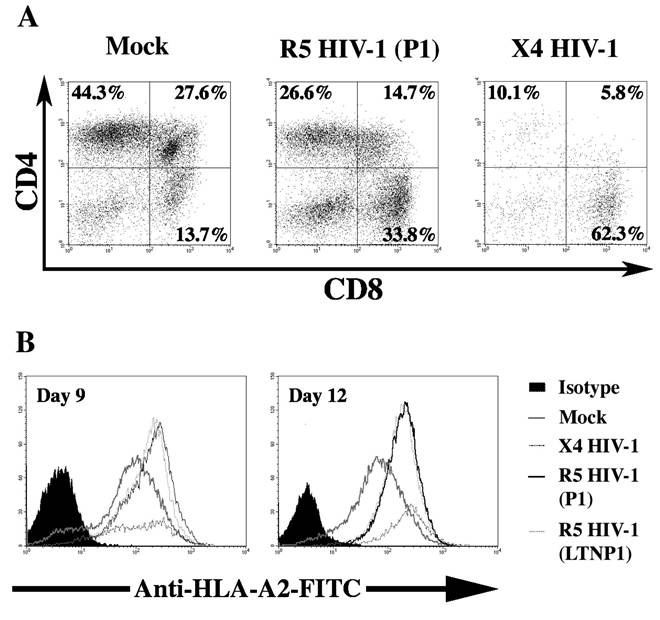
FIGURE 18.1 Pathogenesis of HIV-1 in thymocytes. FTOC was infected with the X4 HIV-1 molecular clone NL4-3, R5 HIV-1 progressor clone P1 (ACH142 *E11), R5 HIV-1 long-term nonprogressor clone LTNP 1 (ACH441 39.2g10) or was mock infected. FTOC was carried in the presence of IL-2 (20 IU/ml), IL-4 (0.5 ng/ml) and IL-7 (1 ng/ml). FTOC was maintained for 12 days with daily media changes. (A) R5 and X4 HIV-1 deplete CD4+ thymocytes in FTOC. Thymocytes were recovered 12 days after infection and stained for surface expression of CD4 and CD8. Representative dot plots for one of three independent experiments are shown. (B) Up-regulation of cell, surface MHC-1 protein in FTOC infected with HIV-1. Thymocytes were recovered 9 and 12 days after infection and stained with monoclonal antibody BB7-2 fluorescein isothiocyanate (FITC) directed at HLA-A2.
infants of HIV-positive mothers.36 Berkowitz et al. showed that CXCR4 is expressed at various stages of thymocyte development and is highly expressed on immature (CD3-, CD4+, CD8-) intrathymic T progenitor (ITTP) cells.37 X4 HIV-1 can infect and deplete ITTP, thereby interfering with thymocyte development.11’14 In HIV-1-infected FTOC, an increased expression of MHC class I (MHC I) was observed on ITTP and double-positive (DP) thymocytes due to increased IFN-α and IL-10 production (Figure 18.1B).23,31 This increased expression of MHC I interferes with thymocyte development and was shown to be responsible for generation of dysfunctional CD8+ T cells.22 ITTP cells, however, express undetectable levels of CCR5 and are, therefore, not targets for R5 HIV-1 infection or direct killing.37 IL-7 exhibits a strong antiapoptotic effect on early lymphoid progenitor cells,38-40 in part by increasing intracellular Bcl-2 levels.41’42 Moreover, IL-7 treatment of FTOC increases the proliferation of immature CD3-∕CD3low thymocytes and stimulates T cell receptor (TCR)-αβ gene rearrangements.42
Secondary Lymphoid Tissue Organ Culture (LTOC)
Secondary lymphoid organs are the sites where the majority of lymphocytes reside.43 They are also the sites of antigen presentation and lymphocyte activation and are, therefore, crucial for both viral replication and pathogenesis.43 Mucosal lymphoid tissue is the dominant site of HIV-1 infection during the initial, acute phase of infection.44-45 Mucosal CD4+ T cells constitute 50% of all CD4+ T cells and predominantly express CCR5. Mucosal T cells are, therefore, targets of R5 HIV-1, which predominates early in infection.44-46 The infection of macaques with simian immunodeficiency virus (SIV) is reported to cause severe mucosal CD4+ T cell depletion within a few weeks.47 It is likely that HIV-1 infection of humans also causes this devastating depletion of mucosal CD4+ thymocytes within a few weeks of infection, thereby undermining peripheral CD4+ T cell counts.48 HIV-1 also invades and replicates in other lymphoid organs during the acute and chronic phases of infection.
An accumulation of HIV-1 has been observed in or on the follicular dendritic cells (FDCs) of lymph node germinal centers, thereby infecting lymphocytes and macrophages, which reside in or pass through the lymph node.49 FDCs may constitute a major reservoir of HIV-1 infection.50-54 HIV-1 infection of lymphoid tissue ultimately results in destruction of lymphoid tissue architecture that is important for the peripheral expansion of T cells.55 Suppression of HIV-1 replication by HAART may restrict lymphoid tissue damage and facilitate subsequent immune reconstitution.48Apoptosis plays a major role in HIV-1-mediated destruction of lymphocytes and lymphoid tissue (LT) architecture. Enhanced apoptosis can be seen in lymphoid tissue of HIV-1-infected individuals.56,57 In situ labeling of lymph nodes from HIV-1-infected children and SIV-1-infected macaques has suggested that apoptosis occurs predominately in bystander cells, not in productively infected cells.56,58 A significant correlation was observed between the degree of cellular activation and the intensity of apoptosis in lymphoid tissue associated with HIV-1 infection, which was independent of the rate of progression to disease or viral load.56 The induced expression of the peripheral lymph node homing receptor (CD62L) by abortive HIV-1 infection of resting memory T cells was also implicated as a cause of apoptosis of these populations in the lymph node.59-61 More systematic studies of mechanisms of HIV-1-induced apoptosis in LT, however, require an animal or organ culture model. Primary cell culture lacks the complexity of LT and may not represent the true nature of signals that initiate cell death pathways.
Margolis and colleagues first developed human tonsil histoculture as an ex vivo model of HIV-1 infection and pathogenesis.62,63 Spleen and lymph node histocultures were used later.62-64 We will collectively refer to them as lymphoid tissue organ culture (LTOC).
Secondary lymphoid organs were dissected into 2- to 3-mm-diameter tissue blocks and cultured on collagen gel supports at the liquid/air interface. LTOC was infected ex vivo with HIV, and cultures were maintained up to 4 weeks. Immunohistochemical analysis of tonsil histoculture revealed well-defined tissue architecture typical of normal lymphoid organs.63 LTOC can be productively infected with both X4 and R5 HIV-1 and does not require stimulation by phytohemagglutinin (PHA) or IL-2 for infection.62,63 Langerhans cells that play a key role in transmission of HIV-1 in mucosal and lymph nodes can transmit R5 and X4 HIV-1 infection to LTOC ex vivo.65 Eckstein and colleagues observed that HIV-1 infection, replication, and cell depletion in tonsil histoculture are not dependent on target cell maturation or proliferation status.66 Moreover, the system accurately reflects viral phenotype, HIV-1 Nef and Vpr mutants replicated to lower levels than the wild-type virus.67,68 HIV-1 Nef variants, which exhibited greater CD4 downmodulating activity, caused severe depletion of CD4+ T cells in lymphoid tissues.69 Furthermore, X4 HIV-1 exhibited greater CPEs than R5 HIV-1.62-64,68,70-72 The difference in CPE, however, could not be correlated with viral replication, as both R5 and X4 HIV-1 replicated to similar levels.62,64 The replication of R5 HIV-1 in macrophages may have contributed to its high viral load.67 Grivel and colleagues observed that both R5 and X4 HIV-1 are equally cytopathic for their targets in lymphoid tissue.73 The CD4+ CCR5+ population, however, constitutes a minor fraction of CD4+ cells and, therefore, their depletion by R5 HIV-1 does not have a major impact on the total CD4+ T cell population. R5 HIV-1 does cause AIDS in humans, but the mechanisms by which it depletes CD4+ T cells remain largely unclear.Both direct and indirect cell killing were reported in HIV-1-infected LTOC. The productive infection of lymphoid tissue ex vivo with X4 HIV-1 induced apoptosis in CD4+ T cells but spared the uninfected CD8+ T cells.73,74 Similarly, R5 HIV-1 infection induced apoptosis in CD4+ cells; however, it was restricted to the CD4+ CCR5+ subset, sparing the CD4+ CXCR4+ CCR5 subset.74 Moreover, no significant CD8+ T cell apoptosis was observed during HIV-1 infection.74 These results suggest that apoptosis cannot be attributed to destruction of general tissue architecture or creation of conditions detrimental to the viability of all cells within the same tissue block.73,74 Nevirapine, a nonnucleoside analogue reverse transcriptase inhibitor (NNRTI), protected uninfected cells residing near productively infected cells, further suggesting that HIV-1 causes cell death primarily by direct killing of infected cells.75 This is in contrast to observations made by Jekle and colleagues, who found that apoptosis of uninfected bystander CD4+ T cells is a major mechanism of lymphocyte depletion in LTOC by X4 HIV-1 but not R5 HIV-1.76 Moreover, X4 HIV-1-induced bystander apoptosis was linked to viral envelope glycoprotein gp120. Both AMD 3100 and the anti-CXCR4 monoclonal antibody 12G5, which specifically blocks the binding of X4 HIV-1 to CXCR4, prevented bystander killing. This is consistent with the recent observation that increased co-receptor affinity leads to increased apoptosis for both R5 and X4 HIV-1.77 NF-κB and p53 are the major apoptosis-inducing factors that are activated by the HIV-1 envelope. Perfettini and colleagues observed enhanced IkB phosphorylation, p53 activation, and higher PUMA levels in lymph nodes of HIV-1 infected people than in those of uninfected controls. The increase in PUMA level was found to be p53 dependent and necessary for syncytial apoptosis.78