Description and Pathophysiology of the ART-Related Abnormal Fat Distribution
Description
Different forms of abnormal fat distribution or lipodystrophies are seen in HIV-infected patients undergoing HAART. Lipoatrophy mainly affects peripheral fat at the level of the limbs, buttocks, face, and abdomen and could be clinically evaluated by the reduced skin-fold due to the decrease in subcutaneous fat.
Results from the US FRAM study revealed that, in this population with high BMI in the control group, both peripheral and central lipoatrophy was associated with HIV infection [50]. Central lipoatrophy, inside the abdomen, can be diagnosed by imaging technologies such as MRI or CT scans. In addition, fat is increased in some regions located primarily in the visceral area but also in the chest and neck, giving an aspect of a buffalo hump. When lipoatro- phy and lipohypertrophy are associated, a phenotype of mixed lipodystrophy is present. This syndrome resembles the typical metabolic syndrome but is more striking since both the loss of the peripheral fat and the increase in visceral fat can be very marked in the same patient.A number of studies have now clearly shown that peripheral lipoatrophy is related to the duration of the treatment with stavu- dine or to a lesser extent with zidovudine and that switch strategies towards other NRTI have allowed a slow recovery [51]. While the prevalence of lipodystrophies was important when these NRTI and first generation PIs were widely used, their occurrence seems to be low with the new-generation ART. Atazanavir, appears to exert few adverse effects on the fat tissue. Healthy subjects [35] or HIV-infected patients [52] treated with atazanavir for 10 days or 48 weeks, respectively, do not develop metabolic disorders or fat tissue redistribution. Moreover, naive HIV-infected patients treated with atazanavir for 96 weeks do not suffer from late-emerging adverse effects [53], though change in fat tissue distribution has not been evaluated in this study.
Up to now, treatments with second-generation NRTIs (abacavir or tenofovir), as well as non- NRTIs (nevirapine, efavirenz) or the fusion inhibitor (enfuvirtide) are considered to have a low incidence on lipodystrophy [54-57].The pathophysiology of ART-related lipodystrophies remains not entirely understood due to their multifactorial origin. Among the numerous factors found to be concerned, drugs play the leading role, but other factors linked to the disease and to the patients themselves also have to be considered. Studies performed with cohorts of patients have outlined the importance of the severity of HIV infection, of the quality of immune restoration, as well as of age, sex, and BMI in the prevalence, type, and severity of lipodystrophy. Among the NRTIs, stavudine in particular as well as zidovudine were found to be linked to peripheral lipoatrophy and PIs to visceral fat accumulation. Their association increases their prevalence and severity [58, 59]. However, individual molecules of these two classes have different effects.
In Vitro Studies
To better understand the pathophysiology of fat alterations, studies were performed first in vitro, so as to decipher the precise role of each drug on adipocyte functions. Most of the studies concerned PIs, since lipodystrophies were diagnosed shortly after their introduction. Several studies reported that in the short term, PIs were able to inhibit the insulin-activated glucose transporter GLUT4, thereby rapidly inducing insulin resistance, which was reversible when the PI concentration was lowered [48]. In the long term (several days), most of the studies reported that some PIs, but not all, were able to greatly alter adipocyte functions by decreasing adipocyte differentiation and by inducing insulin resistance [60]. Our group and others have reported that the step of the adipogenic transcription factor SREBP-1 was specifically targeted by some but not all PIs, with a decreased level of the protein and a decreased activation of the downstream pathways.
We have shown that some PIs were able to impair the nuclear location of SREBP-1 and to alter the structure and stability of the nuclear lamina, by impairing the maturation of prelamin A to lamin A [61-63]. Lamin A and lamin C are encoded by the same gene and they can combine with lamin B to form a meshwork of filamentous proteins that lines the inner nuclear membrane called lamina. Lamina interacts with the nuclear membrane and with chromatin. In particular, the C-termi- nal globular domain of lamin A/C can bind DNA and also SREBP-1 [64, 65]. Interestingly, mutations in this domain are responsible for a genetic form of partial lipodystrophy, Dunnigan’s syndrome or FPLD, with peripheral lipoatrophy, accumulation of fat at the level of the face and neck, and major metabolic alterations with hyper-TG, diabetes, and insulin resistance. The mutations responsible for FPLD reduced the interactions of lamin with DNA and SREBP-1, which could explain the altered adipose tissue differentiation observed in these patients. The cells from these patients present nuclear alterations and altered lamina stability, similar to the alterations induced in cultured adipocytes by some PIs [66]. Thus, we can hypothesize that some PIs could alter lamina structure by comparing lamin A maturation and thereby altering the SREBP-1 normal location inside the nucleus [62]. This could impair adipocyte differentiation and induce insulin resistance. Importantly, in vitro studies have revealed that some PI were able to increase the production of reactive oxygen species (ROS) in mitochondria leading to an increased oxidative stress which alters adipocyte function. In addition, increased ROS production induced by some PIs alters the expression of adipokines in cultured adipocytes [67-69]; they increased the expression of TNF-α and/or IL-6, the proin- flammatory cytokines that could play a role in adipose tissue insulin resistance and apoptosis. They were also shown to decrease the expression of adiponectin, which could explain, at least in part, the resistance to insulin.As regards NRTIs, only a few in vitro studies have been presented. We have observed that the thymidine analogues, stavudine and zidovudine, but not the other NRTIs, were able to induce a mitochondrial dysfunction, increase ROS production and decrease lipid content in adipocytes [69, 70]. In addition, thymidine analogues also altered the expression of adipokines in vitro, in part as results of increased ROS production [68, 69].
Taken as a whole, in the two classes of ART, some drugs modified adipocyte function. PIs altered differentiation, mitochondrial function and insulin sensitivity possibly through their effect on lamin A/C and SREBP-1 and on ROS production, while NRTIs altered lipid content and adipokine production through their effect on mitochondria and ROS. Therefore, when present together, a synergistic effect could be hypothesized (Fig. 8).
Ex Vivo and In Vivo Studies
To go further, different groups performed studies on subcutaneous adipose tissue from lipodystrophic patients, which were compared with fat from control subjects or from non-lipodystrophic patients undergo-
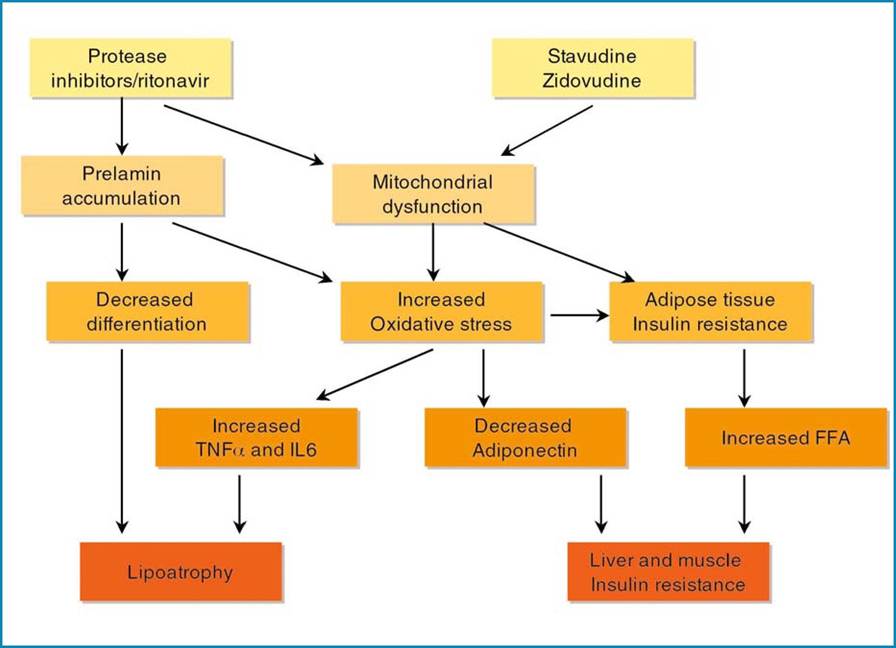
Fig. 8 Hypothetical scheme of the mechanisms responsible for Iipoatrophy and insulin resistance in HAART-related lipodystrophy
ing HAART. The level of mitochondrial DNA was evaluated by several groups and consistently found to be decreased in fat from HAART-treated patients: this decrease was seen in patients undergoing stavudine treatment and to a lesser extent with zidovudine, in accordance with the hypothesis that these NRTIs can alter mitochondrial function [58]. Morphological studies revealed important morphological alterations in patients’ lipoatrophic fat with adipocytes of reduced size, increased apoptosis, increased fibrosis, and mitochondria numbers [70-75]. An increased number of macrophages invading lipoatrophic adipose tissue was reported by these groups.
The expression of the transcription factors SREBP-1, PPARγ, and C∕EBPα was found to be decreased in fat from lipodys- trophic as compared to non-lipodystrophic and control subjects, indicating a state of altered differentiation and insulin resistance [71, 75, 76]. In addition, the expression of the adipokines was deeply altered in fat from lipodystrophic patients as compared to control or non-lipodystrophic patients; TNF- α and IL-6 expression was increased, while that of leptin and adiponectin was decreased [71, 75, 76]. Accordingly, the circulating levels of these cytokines were altered [42, 43].
Clinical data on the effects of individual molecules on lipodystrophy are now becoming available. A role for thymidine analogues, stavudine in particular, in peripheral lipoatrophy has been revealed by different studies, which found a strong correlation between the presence of peripheral Iipoatrophy and the use of these molecules [49]. In addition, in the MITOX study, when Iipoatrophic patients were switched from thymidine analogs to abacavir, peripheral fat slowly increased [51], arguing again for the implication of thymidine analogs in lipoatrophy. Other switch studies, TARHEEL or RAVE found similar results [55, 56].
Increased apoptosis was reported in fat from patients undergoing treatment with PIs and NRTIs. Domingo [77] found that when the patients were switched from PIs to nevirapine, fat-cell apoptosis persisted. Otherwise, when patients were switched from stavudine to ABC, they recovered about 20% of peripheral fat after 1 year. However, increased mDNA level remained two times lower than that of control fat and even if apoptosis was reduced, it remained higher than that observed in control fat [55]. This adipose tissue toxicity could result from the ability of these molecules to induce mitochondrial dysfunction and alter adipokine secretion. In addition, the fact that apoptosis was only partly reverted argues in favor of a role for other ART, including PIs, in lipoatrophy (Fig.
8).We have performed a study to evaluate the specific role of the different drugs in adipose tissue alterations. To that end, we examined the reversibility of adipose tissue alterations in HIV-infected patients after a 6-month (M6) interruption of ART; A 6month ART interruption markedly improved adipose tissue functions, although fat distribution did not visibly change. Stavudine and zidovudine were associated with marked inflammation, which improved when these drugs were withdrawn; they also had a negative impact on differentiation and mitochondrial status. PI were also associated with altered adipocyte differentiation and mitochondrial status. These data clearly show the detrimental impact of antiretroviral drugs, and particularly thymidine analogues, on adipose tissue and argue for switch strategies sparing these drugs [78].
Pathophysiology of Increased Visceral Fat in HAART-Related Lipodystrophy
While the mechanisms responsible for peripheral lipoatrophy begin to be understood, as fat biopsies can be performed to analyze adipose tissue alterations, only speculative mechanisms can be proposed for visceral fat hypertrophy, since to date no results have been presented on fat from this location. One working hypothesis could refer to the presence of 11β-HSD1 in greater amounts in visceral than in subcutaneous fat, which will result in an increased synthesis of cortisol from cortisone and an endogenous activation of fat hypertrophy. Such a mechanism has been postulated in the classic metabolic syndrome.
In addition, TNF-α has been shown to activate this enzyme. If we hypothesize that TNF-α production is increased in HAART- treated patients due to the effect of therapeutic molecules, this could lead to an even greater activation of the enzyme. Moreover, Ledru et al. has shown that CD4 and CD8 lymphocytes in HAART-treated patients produced TNF-α and were resistant to TNF- α-induced apoptosis [79], which would further increase TNF-α levels. Otherwise, in subcutaneous fat, we observed the presence of activated macrophages that release TNF- α [75, 78]. This could also be the case in visceral fat as found in animal models of obesity [26, 27]. Therefore, it could be proposed that increased TNF-α could hyperactivate 11β-HSD 1 and result in an increased synthesis of cortisol inside the adipocytes, thereby favoring their hypertrophy. In addition, other hormones could modulate this enzyme: growth hormone (GH) and testosterone were found to inactivate the enzyme. It is interesting to note that HIV-infected patients with lipodystrophy often present a relative decrease in GH [80] and testosterone levels. Moreover, when patients with visceral fat hypertrophy were treated with GH, a reduction in the amount of visceral
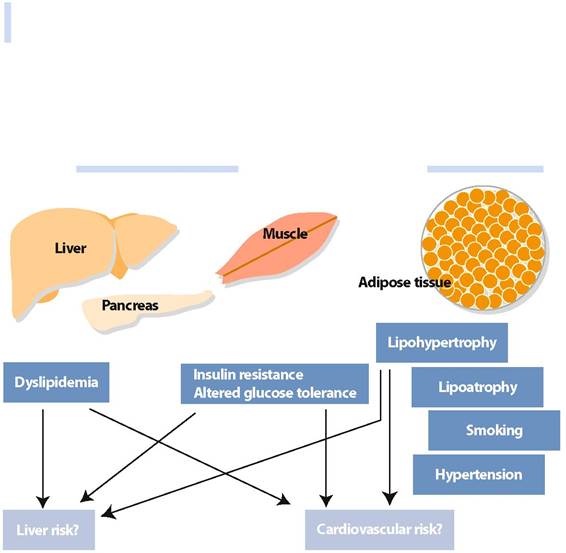
Fig. 9 Pathophysiology of HAART-related metabolic syndrome
fat was consistently reported [81] and reduced truncal adiposity has also been reported in another study with lipodys- trophic HIV-infected patients [82].