DIRECT EFFECT OF VIRAL PROTEINS
HIV-1 gp120 envelope protein was shown to be neurotoxic in serum-free primary neuronal cultures (serum-free neuronal cultures contain few nonneuronal cells), in neuroblastoma cell lines, and in mature neuronal differentiated human NT2 cells.99-102 Soluble HIV-1 gp120 can bind CXCR4, and perhaps CCR5, on neurons in the absence of CD4100 to induce neuronal signaling and apoptosis,103 and blocking this interaction was shown to prevent gp120-induced neuronal apoptosis.104 Recombinant gp120 from the macrophage tropic HIV-1 BaL isolate also induced apoptosis in cultured human neuronal NT2 cells,101 suggesting that HIV-1-induced neuronal cell death may be mediated through the direct interaction of gp120 with CCR5 expressed on neurons.
In addition to gp120, it was suggested that HIV-1 gp41, the transmembrane region of the envelope protein, is neurotoxic.105 Similarly, full-length envelope-derived proteins from FIV are directly cytotoxic to neurons.106,107 Recent work from our group indicates that both replication-competent and replication-incompetent virions are highly cytotoxic to neuronal cultures in an envelope-sequence-dependent manner.108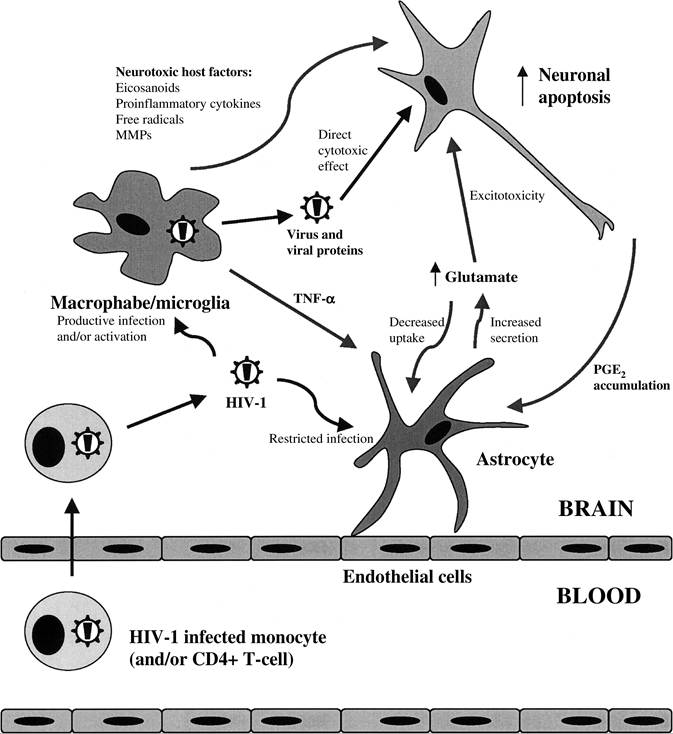
FIGURE 22.1 Neuronal apoptosis results from the direct cytotoxicity of HIV-1 proteins, the production of neurotoxic factors from HIV-1-infected or -activated macrophages/microglia, and the disruption of glutamate homeostasis, resulting in neuronal cell death through an excitotoxic pathway involving hyperstimulation of the NMDA receptor. MMPs, matrix metalloproteinases; TNF-α, tumor necrosis factor-α; PGE2, prostaglandin E2.
Although in vitro studies demonstrated that CXCR4-dependent HIV-1 isolates induced the highest level of neuronal death,109 the majority of brain-derived viruses are CCR5 dependent.110,111 Some CCR5/CXCR4 dualtropic viruses were identified in the brains of HIV-1-infected individuals,111 but detection of virus that is solely CXCR4 dependent is rare.
Experiments using neuroblastoma cell lines have supported the hypothesis that several other HIV-1 viral proteins can directly injure neurons without the intermediary role of nonneuronal cells.
HIV-1 Tat can be taken up into rat PC 12 neuronal cells by a receptor-mediated mechanism.112 Compared with BSA-treated control cells, HIV-1 Tat (at a concentration as low as 1 nM) induced significantly greater apoptosis of PC12 cells, 72 h after stimulation.113 In addition, HIV-1 Tat was shown to induce apoptosis in human neuroblastoma SK-N-MC cells in a dose-dependent manner.114 In this study, the specificity for Tat-induced apoptosis was confirmed using Tat-neutralizing antiserum. These findings are underscored by the early observation that the Tat protein from the animal Ientivirus maedi-visna virus is also neurotoxic.115 Experiments using human NT2 cells (stably differentiated postmitotic neurons) demonstrated that extracellular HIV-1 Vpr induced apoptosis in a dose-dependent fashion.116 In addition, this latter group showed that when delivered intracellularly, HIV-1 Vpr was capable of inducing apoptosis of mature neuronal differentiated human NT2 cells.117 Trillo-Pazos et al. demonstrated that 100 ng/ml of recombinant HIV-1 Nef caused a 30% decrease in cell number in the human SK-N-SH neuroblastoma cell line after a single exposure over 3 days.118 At this time, fragmented nuclei were detected in the cell line, suggesting that the Nef-induced toxicity might also be mediated via apoptosis. However, these studies are complicated by the observation that HIV-1-infected cells minimally release the Nef protein.