EXTRA-CELLULAR HMGB1: ORIGINS AND MECHANISMS OF RELEASE FROM CELL
HMGB1 biological properties cannot be limited to its nuclear functions, as recently illustrated by a series of surprising observations. H. Rauvala and co-workers were the first ones to describe extracellular activities for HMGB1.
In a series of remarkable publications, they showed that a cellular, membrane-bound protein called amphoterin (another name for HMGB1) was highly expressed in embryonic rat neurons, and could promote neurite outgrowth even in a purified form. Later, this activity was proved to require the binding of amphoterin to RAGE, the receptor for advanced glycation end-products [7-9].The potential of HMGB1 as a soluble factor was eventually recognized in 1999 when Kevin Tracey’s group established that HMGB1 was a central pro-inflammatory cytokine involved in the most severe forms of sepsis [10]. Notably, the authors demonstrated that HMGB1 could be actively secreted by activated macrophages in response to bacterial lipopolysaccharide (LPS) and took part to the late phase of sepsis, at the contrary to TNF-alpha or IL-1β that are known to act in the earliest phases of the disease. This discovery immediately pointed out the therapeutic potential of strategies that would modulate HMGB1 pro-inflammatory activities in sepsis and in the numerous inflammatory conditions where it has been involved later.
Two different mechanisms may lead to the release of HMGB1 into the extracellular environment (Fig. 2). The first relies on an active secretion by immuno-competent cells, such as mononuclear phagocytes, dendritic cells or natural killer cells, following exposure to exogenous bacterial products such as LPS or classical pro- inflammatory cytokines such as TNF-α, IL-1β or IFN-γ [11-13]. Similarly to IL-1β, HMGB1 lacks leader sequence and its extracellular release may result from an unusual secretory pathway.
Notably, HMGB1 secretion would rely on the acetylation of several clusters of lysine that are distributed throughout HMGB1 sequence [2,13]. HMGB1 is highly mobile within the nucleus [14], but it can also shuffle between the nucleus and the cytoplasm. HMGB1 acetylation takes place in the nucleus upon cell stimulation, and prevents HMGB1 re-entry into the nucleus, which may be a prerequisite for addressing HMGB1 to the secretory pathway. The cytosolic HMGB1 would then travel to still un-characterized secretory vesicles that eventually deliver the protein out the cells. The mechanisms controlling HMGB1 acetylation are not yet fully characterized, but they likely involve the inhibition of nuclear deacetylases. Other post-translational modifications have been shown to promote intracytoplasmic sequestration into the cytoplasm and subsequent secretion, such as phosphorylation [15].Activated immuno-competent cells Dying cells
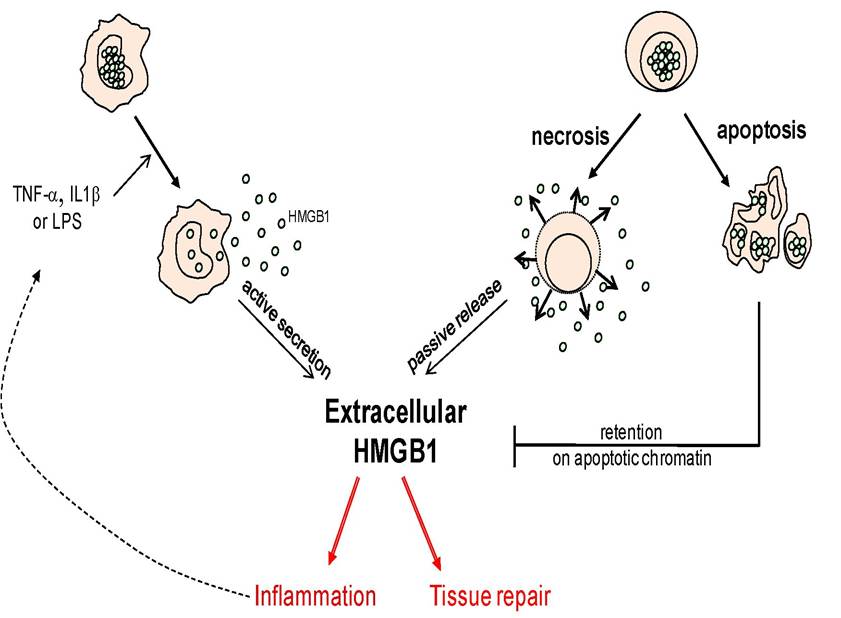
Figure 2: Release of HMGB1 into the extracellular environment. The two recognized sources of HMGB1 release are activated immuno-competent cells (left) and necrotic cells dying through a necrosis process (right), although some forms of apoptosis can also lead to HMGB1 release.
An important contribution to the understanding of the origin and possible functions of extracellular HMGB1 was provided by Bianchi’s laboratory, demonstrating a close link between HMGB1 release and cell death signalisation [16]. Indeed, it was established for the first time that HMGB1 can passively diffuse out of the cells when the plasma membranes are damaged, a process that is observed during primary necrosis. This extracellular form of HMGB1 could promote the secretion of TNF-α from monocytes as well, providing strong evidence that HMGB1 may initiate necrosis-induced inflammation. Conversely, HMGB1 did not diffuse out of apoptotic cells, even during secondary necrosis (also called post-apoptotic necrosis), which was in agreement with the general observation that apoptosis does not promote inflammation most of the time. In this situation HMGB1 was found in a close association with the chromatin, possibly in response to the general histone hypoacetylation that is observed during apoptosis. Importantly, HMGB1 itself does not appear hypoacetylated during apoptosis [16]. This model is widely recognized, but recent works have suggested that some forms of apoptosis could still be compatible with HMGB1 release [17]. The immune tolerance observed during apoptosis may rely in this case on the caspase-induced production of reactive oxygen species (ROS) by mitochondria. ROS indeed mediated the oxidation of cystein residues of HMGB1 that has been shown to inactivate its immunostimulatory activity [18].