VITAMIN D SOURCES AND ENDOGENOUS SYNTHESIS
Vitamin D is a secosteroid hormone that has pleiotropic effects on the regulation of mineral metabolism and the modulation of the immune response [1, 2]. Sources of vitamin D come from dietary intake of foods that are rich in vitamin D, such as fish liver oils and egg yolks, and from endogenous synthesis by photolysis reaction in the skin (Fig.
1).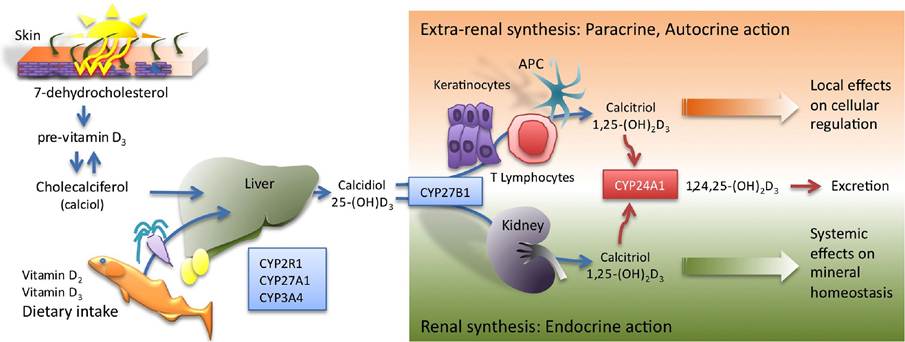
Figure 1: Vitamin D synthesis. Ultraviolet radiation acts on 7-dehydrocholesterol in human skin to form the pre-vitamin D3, that after thermogenic isomerisation forms cholecalciferol (calciol). In addition, vitamin D2 and vitamin D3 can be supplied via food intake. Active vitamin D3 is formed by two sequential hydroxylations in the liver and kidney, to produce 25(OH)D3 (calcidiol) and 1,25(OH)2D3 (calcitriol), respectively. In addition to renal vitamin D synthesis, other tissues and cell types, such as keratinocytes, APC (MAC and DC) and other immune system cells have the capability to produce the active hormone. Vitamin D produced in extra-renal tissues acts locally in a paracrine/autocrine manner and usually does not contribute to the hormone levels in circulation.
There are two main forms of vitamin D used by the human body, vitamin D2 and vitamin D3, which have a chemical structure closely related to the cholesterol molecule. Ultraviolet radiation (UVB), at the 290-315 nm wavelength, alters the cholesterol-based precursor 7-dehydrocholesterol in human skin by breaking C-9 and C10 of the B ring to form the pre-vitamin D3. Following this, naturally occurring thermogenic isomerisation give rise to the unhydroxylated vitamin D3 form cholecalciferol (calciol) that is biologically inactive. In a similar way, vitamin D2 is formed from the irradiation of ergosterol, a plant sterol that can be ingested within the diet.
Endogenously produced vitamin D3 can be selectively transported in the bloodstream by vitamin D binding protein (DBP) to target cells of the vitamin D endocrine system for metabolism. In addition, vitamin D3 supplied by the diet can be absorbed through the duodenum and transported into the lymph via chylomicrons. To reach its active form vitamin D3 undergoes two sequential hydroxylations. First, hydroxylated onto the C25 group by the monooxygenases, mainly P450 cytochrome sterol 27-hydroxylase (CYP27A1), to produce the major circulating vitamin D form, 25-hydroxycholecalciferol (25(OH)D3) also known as calcidiol. This 25(OH) D3 is then transported to the kidney by DBP where it is further hydroxylated onto C-1.This key step is performed in the proximal tubular cells of the nephron by P450 cytochrome 25- hydroxyvitamin D3 1-α-hydroxylase (CYP27B1) enzyme to produce (1,25(OH)2D3), also known as calcitriol, that corresponds to the “active” or “hormonal” form of vitamin D. Calcitriol plays an essential role in stimulating an intestinal absorption of calcium and phosphorus ions, in the mobilization of calcium from bone, and in renal resorption of calcium [23]. While the liver conversion of vitamin D3 to 25(OH)D3 is not actively regulated, the conversion of 25(OH)D3 to the active 1,25(OH)2D3 form is tightly regulated in the kidney by parathyroid hormone (PTH), calcium and phosphorus levels. In response to low calcium and elevated phosphorous, PTH is released by the parathyroid gland to stimulate calcium resorption and to activate CYP27B1 to synthesize more 1,25(OH)2D3. Over-production of 1,25(OH)2D3 is regulated by a negative feedback loop, in that 1,25(OH)2D3 inhibits PTH synthesis and induces 1,25-dihydroxyvitamin D3 24-hydroxylase (CYP24A1) activity in a very sensitive manner [24]. CYP24A1 participates in the 1,25(OH)2D3 catabolism by incorporating a hydroxyl group on C-24 allowing its secretion in the bile.
In addition to renal vitamin D synthesis, other tissues and cell types have the capability to convert 25(OH)D3 to the active 1,25(OH)2D3 form. This conversion is due to the activity of CYP27B1, the same enzyme that is present in the kidney. Extra-renal synthesis was first reported in studies of granulomatous diseases such as sarcoidosis, characterised by an ectopic over production of 1,25(OH)2D3. Primary cultures of pulmonary alveolar MAC of patients with sarcoidosis showed a significant level of conversion of 25(OH)D3 to 1,25(OH)2D3, but, in contrast to renal conversion, macrophage CYP27B1 activity did not respond to the 1,25(OH)2D3 dependent negative regulation, being sensitive to the up-regulation by IFN-γ [25]. This explains that the observed overproduction of 1,25(OH)2D3 and the associated hypocalcaemia that characterises the severe forms of this disease, besides indicate a distinct regulation for the extra-renal enzyme. Extra-renal sites for CYP27B1 expression include skin, endothelium, lymphoid organs, decidua, parathyroid, pancreas, adrenal medulla, colon and cerebellum [26]. Expression of CYP27B1 activity has also been described in normal MAC [27] and other antigen presenting cells such as DC [28] and Langerhan cell (LC) [29]. In addition, T lymphocytes obtained by bronchoalveolar lavage of patients suffering from granulomatous disease such as tuberculosis [30] and T Cell Lymphotrophic Virus-I-transformed Lymphocytes [31] have been shown to produce 1,25(OH)2D3. The functional hormone produced in extra-renal target tissues acts locally in a paracrine/autocrine manner and usually does not contribute to the hormone levels in circulation.
Autonomous synthesis of vitamin D, from 7-dehydrocholesterol to 1,25(OH)2D3, can be achieved in the skin keratinocytes as they can perform the sequential hydroxylation at both C-25 on previtamin D3 to be converted to 25(OH)D3 and at C-1 to transform it to the 1,25(OH)2D3 active form [32].
This autonomous vitamin D3 pathway has also been reported in intestinal CaCo-2 and myeloid THP-1 cell lines [33]. Recently, it has been described that also human primary MAC and DC are also able to perform both hydroxylation steps on vitamin D3 precursor, allowing these cells to develop an autonomous production of active vitamin D [34].This autonomous production of the hormone could be of vital importance for tissues with limited availability of 25(OH)D3 and 1,25(OH)2D3. To reach keratinocytes in the deeper layers of the epidermis 25(OH)D3-carrier complexes must cross from low vascularised stratums. Under these circumstances the low concentration of 25(OH)D3 could impair enough synthesis of the functional hormone. Total serum concentrations of 1,25(OH)2D3 are in the range of 10-11 to 10-10 M, being too low to induce hormonal response on target cells, such as skin keratinocytes in peripheral tissues [35]. Even more if we consider that nearly 99% of serum calcitriol is bound to carrier, which means that less than 1% of serum calcitriol, corresponding to a “functional” concentration of 10-13 M, remains free and is able to induce hormonal response.
The autonomous capacity of keratinocytes to produce functional hormone from vitamin D3 precursor can overcome this limitation. High levels of vitamin D precursors are present in the skin (from UVB photochemical activation) that could be incorporated in the autonomous production of functional 1,25(OH)2D3 in keratinocytes, dermal MAC and DC. In addition, ingested vitamin D precursors can serve as substrate for the autonomous production of functional 1,25(OH)2D3 in MAC and DC from the gastrointestinal tract. Therefore, the direct local conversion of vitamin D precursors to functional hormone in the skin and gut could overcome the limited access to the systemic hormone. This could be essential for these two “barrier” systems to enable an appropriate vitamin D dependent innate immune response.