GENOMIC AND RAPID RESPONSE PATHWAY TO VITAMIN D
Genomic and rapid response pathways to vitamin D have been described on target cells. While slow response normally takes hours to days to reach full manifestation, being dependent on new protein synthesis, rapid response to 1,25(OH)2D3 only needs seconds to minutes to become manifested, being unaffected by inhibitors of transcription and translation.
Genomic responses are initiated upon the interaction between vitamin D compound and nuclear VDR, whereas the rapid response pathway has been related to a putative 1,25(OH)2D3 membrane-associated rapid response steroid-binding protein (MARRS).Evidence for a Vitamin D membrane Receptor
The rapid-acting response has been observed in several contexts as fast intestinal absorption of calcium, secretion of insulin by pancreatic cells, rapid migration of endothelial cells and opening of Ca2+ and Cl- voltage-gated channels in osteoclasts. This rapid and non-genomic related response seems to be mediated by the activation of signal transduction pathways, including protein kinase C, cAMP, intracellular calcium and MAP kinase [36] in target cells through a putative cell membrane-associated receptor for 1,25(OH)2D3. The nature of the membrane receptor for vitamin D has been extensively debated with some authors arguing that the “classical” nuclear VDR could act as the membrane receptor being responsible for the vitamin D rapid response [37]. Recent reports also proposed that an alternate pocket conformation in the ligand-binding domain of the nuclear VDR is involved in the rapid response [38]. A candidate for the rapid response-related membrane binding protein/receptor for 1,25(OH)2D3 was isolated from chicken intestinal basolateral membrane [39] and also detected in mammalian cell membranes [40, 41], termed MARRS for membrane- associated rapid-response to steroid.
The 1,25(OH)2D3-MARRS protein has no sequence similarity with the nuclear VDR. Characterisation of chicken MARRS cDNA sequence reveals it to be identical to the multifunctional protein ERp57, also known as Protein disulfide-isomerase A3 precursor (PDIA3) [40], a protein conserved among several species. ERp57∕PDIA3∕1,25D3-MARRS receptor is a member of the protein disulphide-isomerase (PDI) family of oxidoreductases that contains two redox domains. ERp57 works in a variety of cellular processes and functions. PDIs are involved in native disulphide bond formation of glycoproteins in the endoplasmic reticulum (ER), acting as chaperone proteins. PDIs participate in the antigen presentation process facilitating the formation of disulphide bonds in the nascent MHC class I heavy chains [42,43] and the mentioned 1,25D3-MARRS receptor function [44]. Besides ER, PDIs are present in plasma membrane rafts, cytosol and nucleus [45]. The reducing activity on disulfide bonds mediated by the cell surface PDIs seems to be related with the infective capacity of certain viruses, such as the Sindbis virus [46] and HIV [47]. Thiol-disulfide exchange reactions have been described to be crucial during cell penetration by virus disrupting the rigid protein-protein associations of the envelope and facilitating membrane fusion and release of the viral genome into the cell [46].Antibodies against PDI and inhibitors of disulphide isomerase activity have been described to interfere with virus-host membrane fusion. It has been proved that thiol-disulphide interchange mediates HIV-target cell fusion. Reduction of critical disulphides in viral envelope glycoproteins may be the initial event that triggers conformational changes required for HIV entry [47]. Although, it has yet to be determined whether, and to what extent, these PDI mediated phenomena are affected by the interaction with vitamin D endocrine system.
Nuclear Vitamin D Receptor
Genomic actions of the vitamin D on target cells are mediated by the nuclear VDR, which belongs to the family of steroid-hormone receptors [48].
Full length VDR protein has a molecular weight of 48 kDa that shows nuclear localization motif and several functional domains such as ligand-binding domain to the hormone, DNA-binding domain that binds to the vitamin D response elements (VDREs) located in the promoter region of target genes, dimerization domain which allows its binding with partner retinoic X receptor and transactivation domain that mediates transcriptional activation (Fig. 2).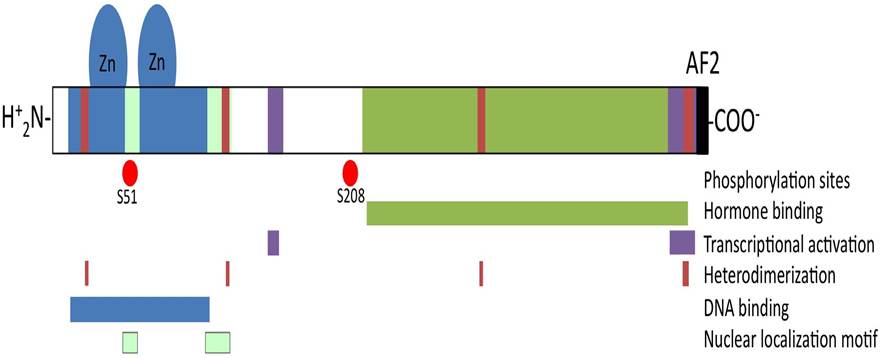
Figure 2: Schematic structure of the vitamin D receptor. VDR protein holds several functional domains. Nuclear localization motifs and DNA binding domain with two zinc fingers motifs are located at the N-terminal region. Hormone- binding domain is located along C-terminal half of the molecule including the activation function (AF)-2 domain on the C-terminal extreme. Scattered regions define the heterodimerization domain that allows binding with partner retinoic X receptor.
Interaction of 1,25(OH)2D3 with VDR induces translocation from the cytosol to the nucleus, where it heterodimerizes with its partner retinoic X receptor (RXR), to form a protein complex that acts as a transcription factor involved in the expression of target genes. Promoter region of vitamin D responsive genes shows sequence motifs acting as vitamin D responsive elements (VDRE) composed of direct repeats of consensus PuG(GfT)TCA motifs [49] that are recognised by the 1,25(OH)2D3-VDR-RXR complex. 1,25(OH)2D3-VDR-RXR complex recognition a VDRE activating motif facilitates the assembly of the transcription initiation complex by the release of corepressors and the recruitment of basal transcription factors and co-regulator molecules, including members of the steroid receptor coactivator (SRC) family and the VDR activating proteins (DRIP) that modulates chromatin remodelling to increase RNA-polymerase II gene transcription rate. Alternatively, when the ligand-VDR-RXR complex is engaged to the inhibitory VDRE, corepressors are recruited to inhibit gene transcription.
In addition, VDRE independent regulation of gene transcription by 1,25(OH)2D3-VDR complex has been described. In this way, 1,25(OH)2D3 exerts regulatory role by interfering with the signal of key immune related transcription factors such as activating protein-1 (AP-1), nuclear factor of activated T-cell (NFAT), specific protein-1 (Spl) and nuclear factor kappa-light-chain-enhancer of activated B cells (NF-KB). The biological actions resulting from 1,25(OH)2D3-VDR transcriptional regulation have pleiotropic effects affecting genes involved on the vitamin D hormone synthesis/catabolism and calcium homeostasis [50], cell differentiation and suppression of cell proliferation [51], neuroprotection in the central nervous system [52] and modulation of innate and adaptative immune response [53].Human VDR gene expands over 100 kb of the chromosome 12q13.11 and consists of 14 exons distributed among 5'UTR region (upstream exons 1a to 1e), protein coding region (exons 2 to 9) and 3'UTR region (exon 9) (Fig. 3). VDR gene expression is under complex transcriptional control by multiple promoters. Distal promoter (1f) and proximal promoters (1a and 1d) generate-multiple variant VDR transcripts showing tissue and cell type specificity [54]. Naturally occurring mutations of VDR gene are responsible for the hereditary 1,25(OH)2D3 dependent rickets (vitamin D dependent rickets type II; VDDR II), that are characterised by defective bone mineralization, low intestinal calcium absorption, hypocalcaemia and elevated serum levels of 1,25(OH)2D3. All VDDR II related VDR mutations are inactivating mutations disturbing VDR DNA binding ability as well as transcriptional transactivation. VDR knockout mice have been obtained, and represent an animal model for the study of VDDR II [55,56].
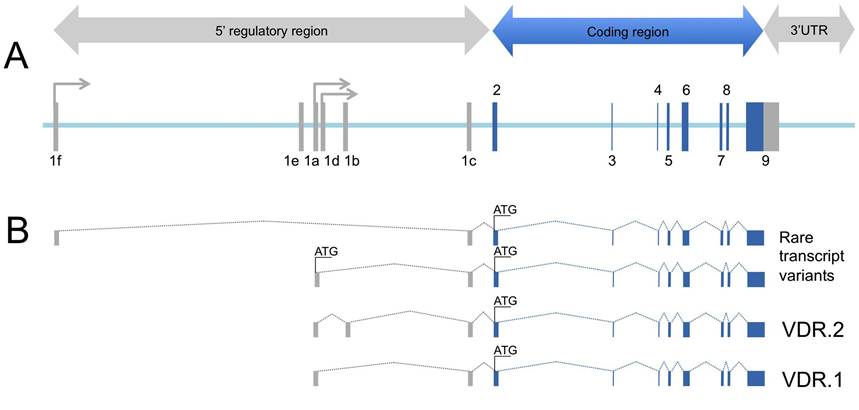
Figure 3: Genomic architecture of the human VDR gene. A, Exon-intron structure of VDR gene with non-coding (grey coloured) and coding (blue coloured) exons numbered according [54]. B, VDR gene expression is under complex transcriptional control by multiple promoters. Distal promoter (1f) and proximal promoters (1a and 1d) generates multiple variant VDR transcripts showing tissue and cell type specificity [54]. Transcript variant 1 (VDR.1) lacks an alternate exon in the 5' UTR (1b) compared to variant 2 (VDR.2). Variants 1 and 2 encode the same protein. Additional rare variants have been described using alternate promoters, at 1f and 1d, and alternative splicing rendering VDR isoforms [54].