1,25(OH)2D3-VDR Mediated Transcriptional Regulation of Immune Related Genes
As for 1,25(OH)2D3-VDR target genes, 1,25(OH)2D3-VDR mediated activation/repression of immune genes can be done in a VDRE dependent and independent manner (Table 1).
Increase of tumour necrosis factor-alpha (TNF)-α production by 1,25(OH)2D3 treated bone marrow cells is mediated by direct binding of the complex 1,25(OH)2D3-VDR-RXR∕VDRE in the TNF-α promoter [57]. Moreover, in peripheral blood mononuclear cells of normal and haemodialysis patients [58], in murine MAC cell line P388D1 [59] and in peritoneal MAC of patients treated with continuous ambulatory peritoneal dialysis [60], 1,25(OH)2D3 has a potent inhibitory effect on the production of TNF-α, independent of nuclear translocation of (NF-KB) [59] and 1,25(OH)2D3- mediated down-regulation of TLR2 and TLR4 [61]. VDRE has been recently found in the promoter region of both the human cathelicidin antimicrobial peptide (CAMP) gene and the defensin-β2 (defβ2) gene, mediating 1,25(OH)2D3 antimicrobial peptide induction in monocytes, MAC and keratinocytes [49, 62].Negative VDRE-mediated response has also been described for the 1,25(OH)2D3-mediated inhibition of IFN- γ [63] and granulocyte/monocyte colony-stimulating factor (GM-CSF) production [64]. In addition, 1,25(OH)2D3-VDR also mediates VDRE independent regulation of gene transcription, through a direct or indirect influence on signalling cascades.
By this way, 1,25(OH)2D3-VDR complex can interfere, in a dose-dependent manner, with the signalling of key transcription factors involved in the regulation of immune related genes. NF-Kb is a major transcription factor that regulates genes responsible for both the innate and adaptive immune response.
Down-regulation of interleukin (IL)-12 and IL-8 is mediated by ligand-VDR complex that impedes both the activation of NF-Kb transcription factor and the binding on the NF-Kb consensus motif in the promoter sequence of these two genes.
Moreover, 1,25(OH)2D3 also influences the activity of NF-Kb by interfering i) with the ubiquitination and subsequent degradation of the cytosolic inhibitor of NF-Kb (IKb-α) and ii) by inhibiting NF-Kb nuclear translocation and DNA binding. Thus, enhanced repression of RelB transcription by 1,25(OH)2D3-VDR contributes to downregulation of the NF-Kb pathway. Additional pathways are mediated by transcription factors, such as NFAT/AP-1 and MAPK, are involved in the 1,25(OH)2D3-VDR mediated inhibition of IL-2, IL-4 and Fas ligand (FasL) genes expression, whereas 1,25(OH)2D3-VDR∕SP-1 may mediate CD14 upregulation (Table 1). 1,25(OH)2D3 can also regulate transcription of NF-Kb genes such as RelB and c-Rel. A negative VDRE has been described in the promotion region of RelB gene that is constitutively linked to unbound VDR. This VDR-promoter association is enhanced by ligand-binding but reduced by LPS [65].Table 1: 1,25(OH)2D3-VDR mediated transcriptional regulation of immune related genes.
| Regulatory mechanism | Target gene | Mechanism and effect on gene transcription | |
| VDRE Mediated Regulation | |||
| 1. | TNF-α | ||
| Activating VDRE | 2. | Cathelicidin | Direct interaction of ligand-VDR-RXR with VDRE. Increase transcription. [49,57] |
| 3. | Defensin-β2 | ||
| 4. | GM-CSF | Direct binding of ligand-VDR monomers to VDRE. Increase transcription. [64] | |
| Repressing VDRE | 5. | IFN-γ | Direct binding of ligand-VDR-RXR with VDRE and inhibition of an upstream enhancer element. Inhibition of transcription. [63] |
| 6. | RelB | Constitutive binding with unliganded-VDR that is increased by ligand and decreased by LPS. Inhibition of transcription. [65] | |
| Non VDRE Mediated Regulation | |||
| 7. | IL12p40 | Indirect inhibition of NF-Kb heterodimer binding to promoter motif. Inhibition | |
| NF-kB pathway | 8. | IL-8 | of transcription. [66,67] |
| 9. | TNF-α | Reducing nuclear translocation of NF-Kβ. Inhibition of transcription. [59] | |
| 10 | IL-2 | ||
| NFATp/AP1 pathway | 11. | IL-4 | Interfering with NFATp/AP-1 complex assembly. Inhibition of transcription. [68] |
| SP-1 transcription factor | 12. | CD14 | VDR/SP-1 interaction at promoter SP-1 binding site. Increase transcription. [70] |
ROLE OF VITAMIN D ON THE IMMUNE RESPONSE
Expression and regulatory properties of key components of vitamin D endocrine system, CYP27A1, CYP27B1, CYP24A1 and VDR, on immune cells strongly supports the role of vitamin D on immune system modulation. Since the first description of 1,25(OH)2D3/VDR metabolism acting in peripheral mononuclear leukocytes and MAC of healthy individuals and sarcoidosis patients [25,71], vitamin D action on immune system has been mainly considered in the context of both immunosuppressive activity on APC and immunomodulatory effect on polarization of Th lymphocyte.
The involvement of vitamin D endocrine system on the induction of innate immune response offers a new and promising picture from which the role of vitamin D on immune system regulation emerges with contrasting and pleiotropic effects.Regulation of Vitamin D Endocrine System on Immune Cells
VDR is expressed by many cell types of the immune system [71,72], in particular APCs such as MAC, DCs [73] and LCs [74,75], as well as in CD4+ and CD8+ T cells [72]. Constitutive expression of VDR has been observed in MAC and DCs whereas their expression is induced by immune stimulus in B and T lymphocytes. Regulation of VDR expression in APC (monocyte/MAC and DC) depends on maturation status.
Immature APC expresses basal levels of VDR, allowing their responsiveness to agonist stimulus. When infectious agents interact with DC and MAC, these cells mature to capture and present the antigens. Maturation process is characterised by the upregulation of CYP27B1 expression and the downregulation of VDR expression (Fig. 4). As a consequence, mature APC produces large amounts of 1,25(OH)2D3 while lose its capability to respond to the hormone. As a result, a paracrine circuit is generated in which neighbouring immature APCs respond to the hormone produced by mature APCs (Fig. 4). Afterwards, 1,25(OH)2D3 produced by mature APC inhibits maturation switch of neighbouring immature cells contributing to suppress immune activation [76,77].
Action of Vitamin D on Innate Immune Response
The main features of vitamin D effect on immune system regulation are depicted in Fig. 5 and Table 2. This interaction is performed by the recognition of pathogen products, named pathogen-associated molecular patterns (PAMPs), a subclass of pattern recognition receptors (PRRs) located in the plasma membrane of these cells. Among membrane-bound PRRs are the transmembrane proteins known as Toll-like receptors
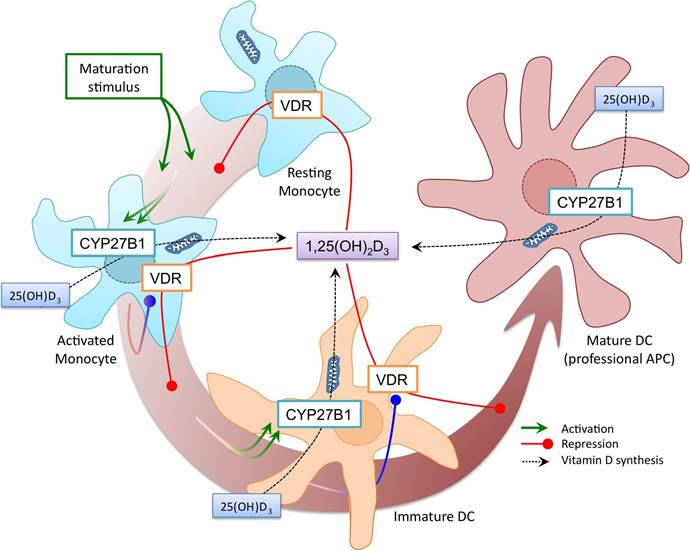
Figure 4: Regulation of vitamin D receptor (VDR) and 25-hydroxy vitamin D3 1-α-hydroxylase (CYP27B1) synthesis during maturation of antigen presenting cells (APC).
APC, such as DC and MAC, express basal levels of VDR that allow their responsiveness to ligand stimulus. Activated monocyte (DC and MAC) become induced to maturate (green lines). Maturation process is characterised by the upregulation of CYP27B1 expression (green degraded lines) and the downregulation of VDR expression (blue degraded lines). As a result, APC produce large amounts of 1,25(OH)2D3 while lost their capability to respond to the hormone. Neighbouring immature APC can responds to 1,25(OH)2D3 secreted by mature APC, by inhibiting star of maturation through ligand-VDR interaction (red lines). By this paracrine circuit 1,25(OH)2D3 can lead to the suppression of further DC development.(TLRs) that have the capability to recognize a broad type of molecules, including peptides, lipids and nucleic acids of viral and bacterial origin [78]. These interactions result in the trigger of downstream signalling cascades, many of which terminate in the activation of the transcription factor NF-Kb. The final result is the induction of the innate immunity responses as well as the instruction of the adaptive immune response against pathogen. Mainly, innate immune response is conducted in the form of antigen phagocytosis and pathogen destruction as well as promotion of microbial killing by host antimicrobial peptides (AMPs). Following this, adaptive immune response is instructed by the initiation of cell (cytotoxic activity by T lymphocytes) and/or humoral (antibody production by B lymphocytes)-directed immune response.
Table 2: Action of Vitamin D on target immune cells.
| Inhibits | Induces/Favours |
| T cells | |
| T cell proliferation [80] | Hyporesponsiveness to self antigens [81, 82] |
| Th1 polarization [83, 84] | Th2 polarization [83] |
| NF-kB activation [85] | T-regulatory cells [86] |
| IL-2 and IFN-γ [63, 66] | CD95 (FasR) [69] |
| GM-CSF [87] | CD 152 (CTLA4) [81] GATA-3 [88] IL-4, IL-5, IL-10 [81] |
Table 2: cont...
| Dendritic cells | |
| Differentiation, maturation [89] IL-12 [66] TNF-α (mature DC) [61] MHC II mediated antigen presentation [90, 91] Costimulatory molecules (CD40, CD40L, CD80, CD80L) [94] DC migration [95] | Apoptosis [81] IL-10 [81] TNF-α (immature DC) [57] Tolerogenic status of myeloid DC [92, 93] CD14 [91] |
| MACMonocyte | |
| Effector function of IFN-γ-activated MAC: Listericidal activity and Chemotactic and phagocytic capacities [92] phagocyte oxidase-mediated burst [96] Costimulatory molecules | |
| (CD40, CD40L, CD80, CD80L) [97] | Cathelicidin [49, 98] |
| TLR2/4 expression [61] | Defensin-β2 [49] |
| TNF-α (in mature cells) [58, 99] | TNF-α (in immature cells) [57] |
| NF-kB activation [59] B cells | CD14 [57] |
| Proliferation of activated B cells [100] Ig production [100] Cell cycle progression [100] Memory B cell differentiation [100] | Apoptosis [100] |
Finely tuned mechanisms are engaged to avoid exacerbate innate and adaptive immunity in which vitamin D plays a key role. 1,25(OH)2D3 synthesis is upregulated in response to TLRs activation, acting in an autocrine/paracrine fashion to promote APMs synthesis, such as cathelicidin and defensin-β2, while at the same time, checking on the vigour of the adaptive immune response to the pathogen [79]. In this way, the capability of 1,25(OH)2D3 to inhibit NF-Kb pathway and to suppress MAC TLR expression supports a key role of the hormone as an autocrine feedback regulator of MAC responses (Fig. 5). 1,25(OH)2D3 mediates innate immune response against Mycobacterium tuberculosis in MAC by the induction of antimicrobial peptides such as cathelicidin [15]. CYP27B1 and VDR genes are induced in MAC as a response to an infection by M. tuberculosis through the AP-1/NF-Kb pathway, induced after TLR2/4 activation. Following this, hydroxylation of 25(OH)D3 to 1,25(OH)2D3 produces large amounts of active vitamin D hormone that, after interaction with VDR, modulates the expression of vitamin D dependent genes. Among these genes, the defensin-like cathelicidin gene is upregulated through the interaction with the VDRE enhancer located on its promoter [98]. There is no feedback control for the induced 1,25(OH)2D3 production in MAC, whereas in other cells the negative feedback is mediated by the 1,25(OH)2D3-catabolic enzyme CYP24A1. While CYP24A1 expression is also induced in MAC together with the expression of CYP27B1 and VDR genes, there is no concomitant increase in 24-hydroxylase-enzyme activity.
It has also been shown that CYP24A1 gene in MAC expresses, in addition to the normal CYP24A1 mRNA, an mRNA splice variant that codes for CYP24A1-SV that lacks the mitochondrial targeting domain [101]. It has been proposed that CYP24A1-SV interferes with normal CYP24A1 enzyme attenuating 24-hydroxylase- enzyme activity, which strongly reduces the conversion of active hormone to catabolic products.
Action of vitamin D on T-lymphocyte activation of TLR initiates protective immune responses with both innate and adaptive profile. Thus, the above mentioned induction of 1,25(OH)2D3/VDR-mediated AMPs production by MAC goes together with the TLR-mediated activation of NF-Kb in APCs, resulting in the release of proinflammatory cytokines and chemokines as well as the upregulation of co-stimulatory molecules that are essential for T-cell activation (Fig. 5). Activation of TLR signalling pathway is a critical element for the induction of the innate immune response as well as for the instruction of the adaptive immune response
against pathogens [102], although an extensive release of TLR-triggered pro-inflammatory mediators can be capable of harming the host, as in the case of sepsis or autoimmune disorders [61].
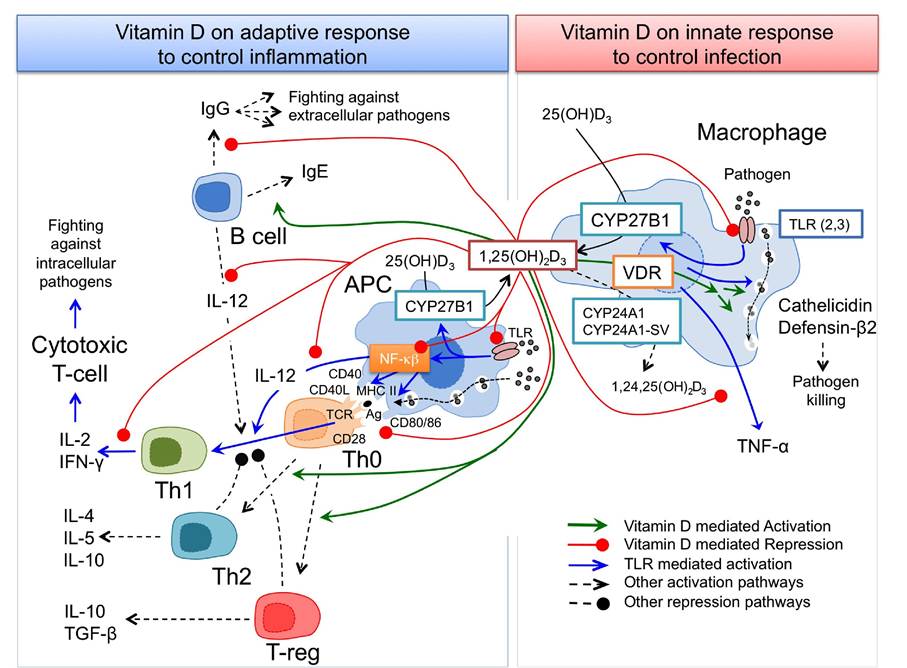
Figure 5: Action of vitamin D on the innate and acquired immune responses. Vitamin D influences innate (right) and adaptive (left) immune responses in a coordinated fashion. Pathogen activated MAC TLR stimulates CYP27B1 synthesis (blue lines) to produce 1,25(OH)2D3 from the 25(OH)D3 precursor (black lines). 1,25(OH)2D3, trough interaction with VDR, promotes synthesis of soluble factors killing pathogens, cathelicidin and defensin-β2 (green lines). Afterward, 1,25(OH)2D3 acts in a paracrine mode to control exacerbated response on APC and T cells. In response to pathogens TLR is also activated in APC that, in addition to activate CYP27B1 synthesis and therefore 1,25(OH)2D3, acts through activation of NF-Kb to promote antigen presentation and Th1 polarization (blue lines). 1,25(OH)2D3 produced by MAC and APCs inhibits NF-Kb pathway (red line) modulating APC response. In addition, 1,25(OH)2D3 inhibited the polarization of Th0 cells to Th1 profile by inhibiting the production of Th1-promoting IL-12 from APC and B cells and down-regulating the expression of Th1 specific cytokines (IL-2 and IFN-γ) (red lines). Finally, locally produced 1,25(OH)2D3 acts on Th0 cells by promoting Th2 and Treg polarization, on B cells by inhibiting IgG and activating IgE production and on MAC by inhibiting TNF-α production. A control on the innate immune response is also assumed by 1,25(OH)2D3 by inhibiting TLR synthesis that contributes to shut off the immune response circuit.
It has been proposed that 1,25(OH)2D3 produced by TLR stimulated MAC or endogenously produced 1,25(OH)2D3 by APC protects from exacerbated TLR signalling by down-regulating the expression of pro- inflammatory cytokines (IFN-γ and TNF-α), antigen-presenting and costimulatory CD40, CD80/86 molecules, and TLR in MAC and APC [61] polarization. The contribution of 1,25(OH)2D3/VDR on the modulation of immune response is also highlighted by its role on T-lymphocyte polarization (Fig. 5). Although 1,25(OH)2D3 can act directly on T cell functions, the main immunomodulatory effects on T cell differentiation are monitored indirectly through APC.
Naive CD4+ T helper (Th) cells, upon stimulation by specific antigen, can differentiate into pluripotent ThO cells that produce a broad pattern of cytokines.
Under the influence of specific stimulatory soluble factors and costimulatory molecules expressed by APCs, pluripotent Th0 cells further differentiate into Th1, Th2 and T regulatory (Treg) cells that express a limited and specific set of cytokines.
Th1 subset is characterised by the expression of the pro-inflammatory cytokines IL-2 and IFN-γ.
The secretion of IL-4, IL-5 and IL-10 cytokines distinguishes Th2 subset.
Finally, a third subset of T cells may be induced, such as Treg cells, characterised by the secretion of IL-10 and TGF-β [103]. The decision, by which Th0 cells undergoes differentiation to one of these T cell subsets, depends on multiple factors from which APC driven stimulus is crucial. Th1 pro-inflammatory response is promoted in the presence of APC secreting IL-12 and presenting MHC-II coupled antigens and co-stimulatory molecules. Antigen presentation by APC in the absence of IL-12 stimulus promotes Th2 differentiation. Finally, APC lacking co-stimulatory molecules become tolerogenic and, in the presence of IL-10, give rise to regulatory T cells or even induce T cell energy [104]. Both Th2 and Treg cells inhibit Th1 differentiation, reason why Th1 profile is prevented if Th2 or Treg has been induced. Th1/Th2 balance hypothesis has been proposed by which each T-helper subset directs different immune response pathways. Type 1 pathway is directed by Th1 cells and is mainly involved in intracellular microbe control and cellular mediated immunity. Th2 cells drive type 2 pathway, being involved in the control of extracellular parasites and in humoral mediated immunity. Although criticised by some authors, Th1/Th2 balance hypothesis has been considered as a paradigm on co-ordinating immune response. Unbalanced Th1/Th2 response predisposes to immune diseases.
Dominance of Th1 drive response has been associated with autoimmunity, whereas Th2 over-dominance has been related with allergy [103]. 1,25(OH)2D3/VDR action on APC is characterised by the down-regulation of the expression of the co-stimulatory molecules CD40, CD80 and CD86, reduction of IL-12 and augmentation of IL-10 production. This resulted in a decreased T-cell activation that creates an environment that favours Th2 and, mostly, Treg differentiation [84, 86, 94, 105]. Inhibition of Th1 response and promotion of Th2 and Treg differentiation has been related with 1,25(OH)2D3-mediated protection against autoimmune diseases [84]. In addition, 1,25(OH)2D3-promoted tolerogenic status, substantiating its therapeutic use to prevent allograph rejection [92].
Effect of Vitamin D on host Thl Mediated Immunity
In the context of infection by intracellular pathogens, the balance between i) positive effects of the 1,25(OH)2D3∕VDR pathway by inducing anti-microbial response, ii) the detrimental effect of Th1 inhibition, and iii) induction of a tolerogenic status has to be considered for each disease model. The suppressive effect of 1,25(OH)2D3/VDR pathway on MAC functions has been revealed detrimental in certain infections. Infection by the intracellular protozoan parasite Leishmania major, host defence is focused around activated MAC acting as IFN-γ-effector cells.
Given the suppressive effect of 1,25(OH)2D3/VDR on Th1-mediated IFN-γ production a detrimental role for 1,25(OH)2D3 can be expected. In line with this, MAC of VDR-knock-out (VDR-KO) mice has stronger leishmanicidal activity that is reduced when exposed to 1,25(OH)2D3, supporting a negative role of the hormone [16].
Analogous results were observed in the LP-BM5 mouse model of AIDS; treatment with 1,25-(OH)2D3 enhanced the severity of the disease and increased the mortality rate [106].
Evidences indicate that HIV-1-specific cellular immune responses may play a critical role in antiviral control [107]. Th1 mediated immune response seems to be poorly developed in HIV-1 infected patients at all stages of the disease. In addition, T-cell proliferative responses against virus are inversely correlated with plasma viremia [107].
Importantly, in many seronegative exposed individuals and long-term non-progressors a vigorous HIV-1 specific Th1 and cytotoxic T-lymphocytes (CTL) responses have been observed [108]. Several authors have proposed a shift from Thl to Th2 immune response that correlates with HIV-1 disease progression [109, 110]. Whatever this Th1>Th2 shift constitutes a viral immune evasion strategy [111], either favoured by opportunistic infections [112] or derived from a selection process against more permissive Th cells [113], the main consequence is that this Th1/Th2 imbalance could preclude to mount humoral and cellular HIV-1 specific immune responses able to control virus infection. According to this, the 1,25(OH)2D3-mediated Th1- >Th2 shift could have detrimental effects on both HIV-1 infection and disease progression to AIDS.