Menstrual physiology
Menstrual cycle physiology may be considered at different levels: hypothalamic, pituitary, ovarian, and endometrial (Figure 40.1). Peptide and steroid hormones provide cross-talk and feedback between these levels to regulate menstruation.
The predominant hormones of the menstrual cycle are gonadotropin-releasing hormone (GnRH), follicle stimulating hormone (FSH), luteinizing hormone (LH), oestrogen (predominantly oestradiol), and progesterone (Table 40.1). Function of this endocrine system is affected by the stage of reproductive life.Hypothalamic-pituitary regulation of ovarian function
The classical model of the menstrual cycle starts with the hypothalamus and GnRH (Figure 40.2). GnRH is produced and released from neurons in the arcuate nucleus of the medial-basal hypothalamus and the preoptic area of the ventral hypothalamus. It is released by axonal transport into the capillaries of the hypophyseal portal system for delivery to the anterior pituitary gland, where GnRH stimulates the synthesis and secretion of LH and FSH from the gonadotrophs.
GnRH is secreted in a pulsatile manner, which is an absolute prerequisite for normal adult reproductive function, including pubertal maturation and fertility (9, 10). The frequency and amplitude of GnRH secretion vary depending on the stage of reproductive life and across the menstrual cycle. In the normal cycling woman,
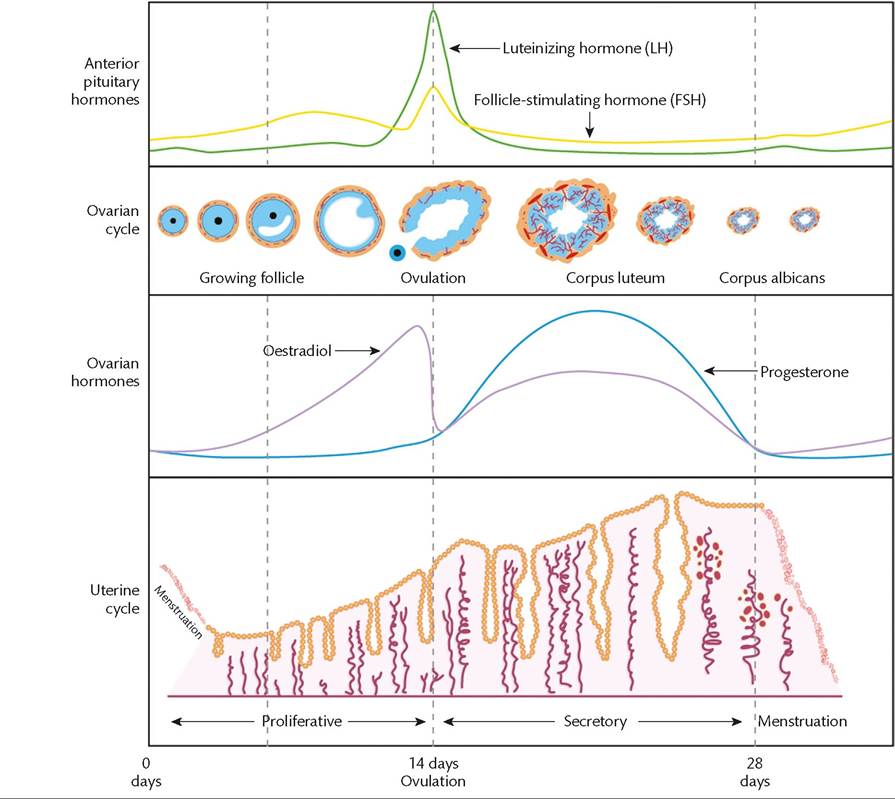
Figure 40.1 Schematic representation of pituitary and ovarian hormones across the menstrual cycle and the response of the ovary and endometrium.
GnRH pulses are of low amplitude, but increase in their frequency during the follicular phase, to a frequency of every 60 minutes during late follicular phase. High GnRH pulse frequency favours LH release, which is also secreted in pulses reflecting those of GnRH and is predominant over FSH in the late follicular phase necessary for ovulation.
On the other hand, luteal phase is characterized by high amplitude and low frequency (approximately every 216 minutes) of pulsatile GnRH secretion. Low GnRH pulsatility stimulates FSH secretion, which is released in a more constitutive manner and is dominant over LH in the luteal and early follicular phase necessary for follicular development. Conversely, continuous GnRH exposure (such as with administration of GnRH analogues) results in desensitization of the gonadotrophs and suppression of LH and FSH secretion. Measurement of GnRH in peripheral circulation is difficult due to its short half-life (to theca cells in the ovary; FSH promotes follicular growth, activates aromatase, and induces expression of LH receptors on the granulosa cell in preparation to respond to the preovulatory LH surge. The granulosa cells also produce inhibin B.During the menstrual cycle GnRH and gonadotropin activity is highly regulated by ovarian feedback loops. In the follicular phase, oestrogen exerts negative feedback at the level of hypothalamus to suppress LH and FSH secretion. Inhibin B more selectively suppresses FSH secretion. However, in the late follicular phase, by yet unclear mechanisms, negative oestrogen feedback switches to positive oestrogen feedback, culminating in the preovulatory LH surge (and to a lesser extent a rise in FSH) necessary for ovulation. Following ovulation, progesterone from the corpus luteum mediates negative feedback to slow down GnRH pulse frequency and subsequently LH secretion. During the luteal phase, inhibin A also suppresses FSH levels. With demise of the corpus luteum in the absence
Table 40.1 Principal hormones of the menstrual cycle
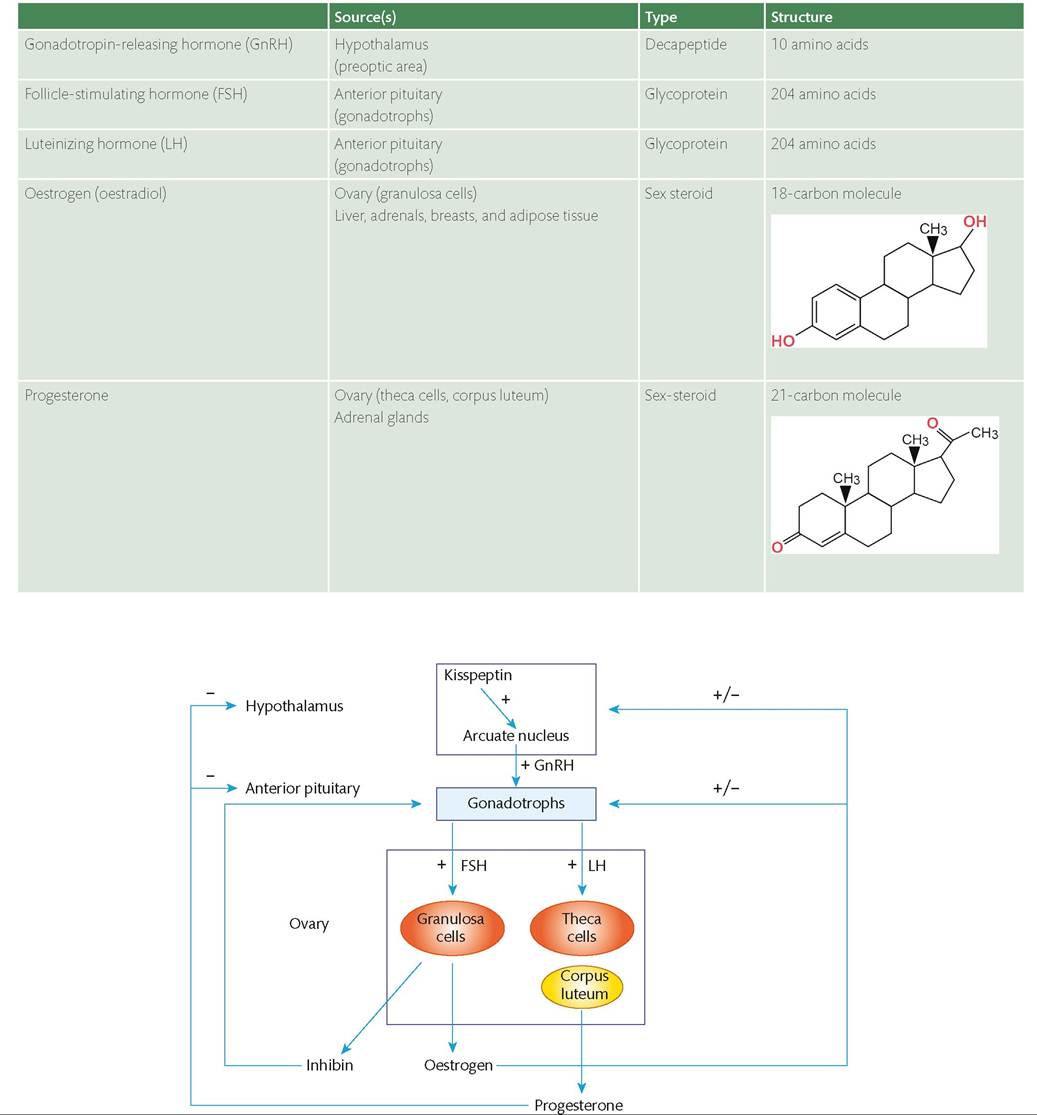
Figure 40.2 The hypothalamic-pituitary-ovarian axis. Hypothalamic gonadotrophin-releasing hormone (GnRH) signals to the anterior pituitary which in turn results in follicle-stimulating hormone (FSH) and luteinizing hormone (LH) secretion.
These stimulate the ovary to produce oestradiol and progesterone to regulate the endometrial function. GnRH and gonadotropin signalling is tightly regulated by negative and positive gonadal-steroid loops and is further regulated by other hypothalamic neuropeptides (such as kisspeptin).
of pregnancy, gonadal steroid secretion declines, resulting in menstruation and stimulation of FSH secretion in response to a loss of negative sex-steroid feedback.
Prepubertal development of the HPO axis and menarche
The ovarian germ cells (oogonia) appear at around day 25, and then migrate to the developing gonad at around day 30 of gestation. They undergo meiosis and differentiate into primary oocytes, having arrested during the first meiotic division before entering prophase I.
Pulsatile GnRH secretion is observed during fetal life from around the 16th week of gestation with sustained gonadotropin and sex-steroid secretion. With advancing gestation, LH and FSH secretion is suppressed due to the negative sex-steroid feedback from the developing ovary. Withdrawal of placental steroids following delivery results in rebound GnRH and gonadotropin activity and persists into the early neonatal period. To distinguish this phenomenon from the true puberty, this phase is referred to as ‘mini-puberty’, where despite the secretion of gonadotropins and sex steroids, ovulation is not present. After the postnatal rise, pulsatile GnRH and gonadotropin secretion become quiescent throughout childhood, with some irregular and low-amplitude pulses, suppressing any activity of the now developed and responsive gonads.
With the onset of puberty there is a nocturnal increase in GnRH secretion and enhanced responsiveness of the pituitary. Slow GnRH pulsatility initially stimulates FSH secretion, resulting in ovarian follicular development and a multicystic appearance of the ovaries, and with the acceleration of GnRH pulsatility LH is secreted.
This precedes any external manifestations of puberty by several years. The factors triggering this process are not yet fully elucidated but appear to be centrally regulated either by an activation of stimulatory signals or a suppression of inhibitory signals. Kisspeptin, a hypothalamic neuropeptide, is now recognized as key regulator of pulsatile GnRH secretion (Figure 40.2) (11). Its obligate role in human puberty was demonstrated by hypogonadotropic pubertal delay seen in patients with an inactivating mutation in kisspeptin (12) and its receptor (13, 14), and precocious puberty in those with activating mutations in the kisspeptin receptor (15).The irregular nocturnal pattern of LH secretion gradually increases until regular LH pulses are established, occurring every 90 minutes irrespective of time of day. This results in the production of gonadal steroids and contribution to the development of external sexual characteristics (thelarche—onset of breast development). As circulating concentrations of oestrogens rise, positive feedback develops to induce a preovulatory LH surge with resultant ovulation and the first experience of menstruation (menarche). Of note, adrenarche (axillary and pubic hair growth) reflects the maturation of the hypothalamic-pituitary-adrenal axis. Many of the initial cycles may be anovulatory, which can contribute to the heavy and irregular menstrual bleeding that can affect some girls in adolescence. After 4-5 years the number of anovulatory cycles reduces from 90% to 20% with the consequent experience of more regular menstrual bleeds.
Menopause
Menopause marks the end of the female natural reproductive life and is a retrospective diagnosis based on the absence of periods for 1 year. It occurs when the supply of primordial follicles available for recruitment is exhausted. This process of follicle loss commences while the fetus and her developing gonads are still in utero (16-18). At the time of menopause there is a corresponding decline in circulating oestrogen levels and gonadotropin levels (FSH and LH) are elevated.
In the perimenopausal transition, fluctuation in oestradiol levels may occur, giving rise to menstruation. While the majority of such cycles are anovulatory, spontaneous ovulation and occasionally pregnancy may occur.Ovarian function
Ovarian function within the menstrual cycle can be broadly divided into the follicular phase, ovulation, luteal phase, and finally the luteal-follicular transition.
Follicular phase
Throughout ovarian life the oocytes reside within follicles. At any time point in the reproductive years there are follicles at different developmental stages within the ovary.
Initially, they are found in primordial follicles, consisting of a primary oocyte surrounded by a single flattened layer of granulosa cells. At the beginning of each menstrual cycle, a cohort of primordial follicles is activated to initiate growth, regulated by both stimulatory and inhibitory molecular signals. The follicles transition into primary follicles, with the granulosa cells becoming cuboidal in shape, before developing into larger preantral follicles. At this stage, the zona pellucida develops between the oocyte and granulosa cells and surrounding stromal cells differentiate to form the theca. Subsequently, the follicles enlarge, developing a fluid-filled cavity termed the antrum. Preantral and early antral follicles produce anti-Mullerian hormone, which appears to have an inhibitory effect on the growth of nearby primordial follicles, thus preventing their activation. This prevents the ovarian reserve being exhausted too quickly. Anti-Mullerian hormone is also therefore an indirect measure of ovarian reserve as it reflects how many growing follicles there are within the ovary—t he higher the serum concentration of anti-Mullerian hormone, the greater the remaining number of follicles (19).
As the follicles grow they acquire FSH and LH receptors and become increasingly gonadotropin dependent, with antral follicles completely dependent on FSH for granulosa cell proliferation and LH for theca cell sex steroidogenesis.
The largest follicle in the cohort is termed the dominant follicle, with all other growing follicles undergoing atresia. The dominant follicle produces oestradiol, resulting in a rapid rise in serum oestradiol concentration and consequent reduction in FSH and LH levels by negative feedback at the hypothalamus. This dominant follicle matures into a preovulatory follicle and expresses LH receptors on both granulosa and theca cells. There is a short period of positive feedback in the late follicular phase whereby increasing oestradiol levels result in a surge of LH. The preovulatory follicle responds to this surge by undergoing ovulation.Ovulation
Following the LH surge and just prior to ovulation, the oocyte responds to its hormonal environment and re-enters meiosis, arresting at metaphase II resulting in a secondary oocyte and a polar body. The granulosa cells decrease oestradiol secretion as a result of decreased sensitivity to FSH. Progesterone production is initiated as a result of LH-driven increased cholesterol side-chain cleavage (P450scc) levels. This enzyme is critical to steroidogenesis, particularly the conversion of cholesterol to pregnenolone. The rise in LH and FSH causes an increase in antral blood flow. This increased vascularity and local secretion of prostaglandins causes an increase in size of the follicle and it distends the surface of the ovary. Proteolytic enzymes are synthesized in the theca and activated by prostaglandins, causing degradation of the distended follicular wall followed by rupture of the follicle capsule. The oocyte, surrounded by the zona pellucida and attached granulosa cells, is ejected from the ruptured follicle. This occurs on approximately day 14 of the menstrual cycle. Under the influence of oestrogens prior to ovulation, increased tubal motility and elevated activity of the densely ciliated fimbriae allows approximation of the aperture of the distal fallopian tube to the ovary. This promotes passage of the expelled oocyte into the tube to be met by the ascending spermatozoa. If fertilization occurs, meiosis is completed.
Luteal phase
Following ovulation, the walls of the ovarian follicle collapse. Under the influence of LH both the theca and the granulosa cells proliferate and the latter develop into luteal cells. There is an influx of lipid droplets and lutein, which gives the corpus luteum its characteristic yellow appearance. Overall, the corpus luteum enlarges to around 15 mm. The oestrogen and progesterone secreted by the luteal cells negatively feed back to the anterior pituitary and levels of FSH and LH decrease. The corpus luteum is reliant on LH for progesterone production and luteolysis occurs in the absence of pregnancy as the mature corpus luteum becomes less sensitive to the remaining LH. This process remains incompletely understood, particularly with regard to the initiating trigger. In addition to reduced gonadotropins, luteolysis is likely a product of decreased LH sensitivity, altered progesterone receptor isoform ratios, and oestrogen receptor (ERα) levels (20, 21). Demise of the corpus luteum begins at around day 24 and the corpus luteum is replaced by whitish scar tissue, the corpus albicans. Over subsequent cycles it is replaced by connective tissue, then absorbed.
Luteal-follicular transition
As the corpus luteum degenerates, circulating concentrations of oestradiol and progesterone rapidly decrease. As a result, FSH and LH plasma concentrations rise and a fresh group of follicles are recruited. The withdrawal of progesterone initiates menses.
Maintenance of the corpus luteum in pregnancy
In the event of conception, the developing embryo secretes human chorionic gonadotropin (hCG) from the syncytial trophoblast. This is detectable in maternal serum from around 8 days following conception. hCG rescues the corpus luteum, preventing menstruation and supporting early embryonic growth. Between 7 and 12 weeks' gestation, oestrogen and progesterone production shifts to the placenta and once the second trimester is entered, the corpus luteum slowly involutes and hCG levels decline.
If the corpus luteum is removed prior to 7 weeks' gestation then miscarriage almost inevitably occurs as a result of progesterone and oestrogen withdrawal. A similar effect can be induced by the use of progesterone antagonists such as mifepristone which underpins its use in the medical management of unwanted pregnancy.
Endometrial response to ovarian function
The endometrium consists of two functionally separate layers overlying the myometrium: functional and basal layers. The functional layer is shed during menses and the basal layer is preserved throughout. Both layers comprise glandular epithelial and stromal cells and the functional layer is covered by a surface epithelial layer. The constituent cells may express receptors for progesterone, oestrogen, and androgens as well as receptors for the glucocorticoid hormone, cortisol (22, 23). The expression of the receptors varies depending on the stage of cycle. Occasional lymphoid follicles may be observed as well as a variety of leucocytes. The endometrium has a complex blo o d supply arising from the radial branches of the uterine artery and the ovarian vessels. There are both short, straight arteries which supply the basal layer and longer, spiral vessels supplying the whole of the endometrium. The spiral vessels connect with the venous system though capillary networks and direct arteriovenous communications.
The endometrial aspect of the menstrual cycle is divided into proliferative, secretory, and menstrual phases, broadly corresponding to the follicular, luteal, and luteal-follicular transition phases of the ovarian cycle. The classical histological description of the cycling endometrium dates from the 1950s (24) but for research purposes dating may be more robustly determined utilizing histological features combined with the date of the reported last menstrual period and measurement of serum oestradiol and progesterone. More recently, advances in molecular phenotyping have added a greater depth of understanding to the molecular and cellular events that underpin menstrual cycle stage and function (25).
Proliferative phase
Following menstruation, the exposed basal layer of the endometrium proliferates rapidly under the influence of rising oestradiol levels. Concurrently, the glandular cells within the endometrium expand: in the early proliferative phase the glandular cells are initially cuboidal and the glands themselves are small but by the late proliferative phase the glands are tortuous and the individual epithelial cells appear columnar. Proliferation is brisk and mitotic figures are observed (Figure 40.3). The stroma is compact throughout the proliferative phase and angiogenesis commences with elongation of the spiral arteries.
Secretory phase
Following ovulation, secretion of progesterone from the corpus luteum inhibits proliferation. The morphological changes associated with progesterone exposure develop between ovulation and approximately 48 hours after progesterone withdrawal (due to demise of the corpus luteum). The endometrial glands become more tortuous and acquire increased secretion of glycoproteins, evident as subnuclear vacuolation (a feature of the early secretory phase). Glandular nuclei move to the centre of cells and mitosis is supressed. The endometrial spiral arterioles undergo remodelling to become increasingly coiled.
The spiral arterioles have a pivotal role at this time because if implantation is successful, the interaction with invading trophoblasts will play a crucial role in the onward pregnancy outcome.
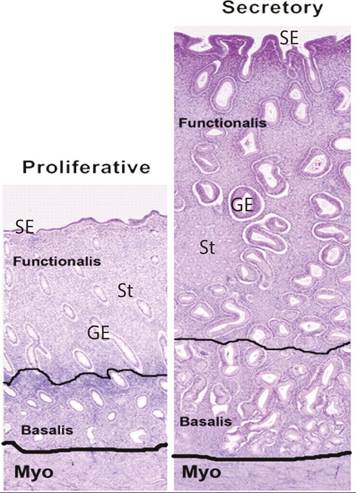
Figure 40.3 Endometrial morphology across the menstrual cycle. Proliferative and secretory endometrium. Surface epithelium (SE), glandular epithelium (GE), and stromal compartment (St) are marked. Note the increased tortuosity of glands in secretory phase and marked thickening of the functional layer.
Adapted from FM Horne and DL Blithe. Progesterone receptor modulators and the endometrium: changes and consequences, Hum Reprod Update 2007;13:567-580.
Menstruation
In the absence of pregnancy the corpus luteum regresses, resulting in a rapid decrease in circulating progesterone and oestradiol. It is progesterone withdrawal that initiates menstruation (Figure 40.4). A local inflammatory response within the endometrium is characterized by cytokine release and infiltration of leucocytes with resultant oedema, activation of matrix metalloproteinases (MMPs), and lysis of the extracellular matrix. This culminates in the shedding of the upper two-thirds of the endometrium (the functional layer). The lower third of the endometrium (basal layer) remains in situ but has an exposed, raw mucosal surface that requires efficient repair.
Inflammatory mediators generated within the endometrium upon withdrawal of progesterone include prostaglandins, cyclooxygenase- 2, MMPs, and interleukin-8 (Figure 40.4). Leucocyte traffic is initially neutrophil dominated although macrophage numbers also increase. Neutrophils contain high levels of MMPs and can activate tissue MMPs and play a critical role in the induction of endometrial shedding. Tight regulation of local endometrial events at the time and appropriate apoptosis is important to secure satisfactory onward endometrial repair. Macrophages are involved in both cytokine production and local remodelling of the endometrium and removal of debris and thus participate in both the breakdown and repair of the endometrium. Tight regulation of localized ‘physiological' inflammation is critical to prevent excessive bleeding at menses (4, 26).
An intact endometrial coagulation system is also necessary for the efficient cessation of menstruation. Endometrial blood vessel injury initiates immediate activation and aggregation of platelets to form a ‘plug'. Subsequently, fibrin is formed via the intrinsic and extrinsic coagulation cascade. Each pathway culminates in the conversion of factor X to Xa, which drives production of thrombin, ultimately leading to the formation of a stable fibrin clot to seal previously bleeding vessels (Figure 40.5) (4). Disorders that interfere with this systemic haemostasis have an impact on menstrual blood loss (see ‘Overview of menstrual pathology') (Figure 40.5).
Conversely, fibrinolysis is the process of conversion of plasminogen to active plasmin, promoting the degradation of fibrin deposits. Tissue plasminogen activator (tPA) and urokinase plasminogen activator (uPA) drive the production of plasmin. In contrast, plasminogen activator inhibitor (PAI) inhibits fibrinolytic activity. The human endometrium contains tPA and uPA, as well as PAI and the uPA receptor (27, 28). Tight regulation to balance coagulation and fibrinolysis is necessary for normal menstruation.
The pivotal role of the coagulation system in menstruation is evidenced by the association between perturbation of this system and heavy menstrual bleeding (HMB). There is evidence that an overactive fibrinolytic system interferes with haemostasis and contributes to HMB. Women with HMB had raised levels of tPA activity on the second day of bleeding compared to those with normal loss (27). The efficacy of tranexamic acid as a treatment for HMB provides further evidence for overactivation of the fibrinolytic system in the endometrium of these women. The antifibrinolytic, tranexamic acid, reduces tPA and PAI levels in women with HMB and results in a 58% reduction in blood loss (29). Furthermore, up to 13% of women with a complaint of HMB will have von Willebrand disease (30).
During menstruation, the shed surface of the endometrium leads to bleeding from damaged vasculature. Factors influencing blood flow include length and radius of a blood vessel and the viscosity of blood. Of these, vessel radius is the dominant contributory factor (31). Therefore, the endometrium has evolved specialized spiral arterioles that have the ability to undergo intense vasoconstriction during the late secretory and menstrual phases to limit menstrual blood loss. Experiments examining the endometrial vasculature in the non-human primate (rhesus macaque) suggest that this vasoconstriction is so intense that the luminal portion of the endometrium becomes hypoxic during menses (32). There is mounting evidence that this hypoxia may trigger a cellular protective response to increase local repair factors and drive blood vessel and tissue regeneration (33, 34).