Amphibian and Reptilian Anatomy and Physiology
Ryan DeVoe
OUTLINE
INTRODUCTION, 542
TAXONOMY, 542
METABOLISM, 542
INTEGUMENT, 543
VISION, 545
Periocular Structures, 545
The Globe and Intraocular Structures, 546
CARDIOVASCULAR SYSTEM, 547
Blood, 548
RESPIRATORY SYSTEM, 549
EARS AND HEARING, 551
GASTROINTESTINAL TRACT, 551
CLOACA, 554
KIDNEYS, 555
REPRODUCTIVE SYSTEM, 556
Male Anatomy, 556
Female Anatomy, 557
Reproductive Cycle, 558
Oviposition, 558
Egg Incubation, 558
Sex Determination, 559
Secondary Sexual Characteristics, 559
Amphibian Reproduction, 559
ENDOCRINE SYSTEM, 560
NERVOUS SYSTEM, 561
MUSCULOSKELETAL SYSTEM, 561
Skull, 561
Axial Skeleton, 562
Appendicular Skeleton, 562
Muscles, 563
SUMMARY, 563
LEARNING OBJECTIVES
When you have completed this chapter you will be able to:
1.
List the taxonomic orders in the classes Reptilia and Amphibia.2. Define ectothermic and explain how ectothermic animals regulate their body temperature.
3. List the unique features of the reptilian and amphibian integumentary systems.
4. Describe the process of ecdysis.
5. List the unique features of reptilian and amphibian vision and hearing.
6. List the components of the reptilian and amphibian heart and describe the flow of blood through the heart.
7. Describe the unique features of the reptilian and amphibian respiratory and gastrointestinal systems.
8. Describe the structure of the kidneys of reptiles and amphibians.
9. Describe the factors that determine the sex of the offspring of reptiles and amphibians.
10. List the unique features of the musculoskeletal systems of reptiles and amphibians.
VOCABULARY FUNDAMENTALS
Acrodont teeth ahck-ruh-dohnt teth
Aglyphous ah-glι-fuhs
Anapsid skull ah-nahp-sihd skuhl Behavioral thermoregulation be-hav-yar-uhl thar-mδ-rehg-u-la-shuhn
Bidder’s organ bihd-darz ohr-gahn
Carapace kear-ah-pas
Chromatophore kro-maht-uh-fδr
Conus papillaris ko-nuhs pah-puh-lear-ihs Diaphragmaticus dι-ah-frahm-aht-uh-kuhs Diapsid skill dι-ahp-sihd skihl
Ecdysis ehck-dih-sihs
Ectothermic ehck-tδ-thar-mihck
Endolymphatic sac ehn-dδ-lihm-fah-tihck sahck Faveloli fah-ve-o-lι
Foramen of panizza fohr-a-mehn o/pah-nez-ah
Hemipenes hehm-ih-pe-nez
Heterophil heht-ar-δ-fihl
Lissencephalic lihs-ehn-seh-fahl-ihck Melanomacrophage mehl-ahn-δ-mah-krδ-faj Multicameral lung muhl-tι-kahm-ar-ahl luhng Nuptial pad nuhp-shuhl pahd
Opisthoglyphous ah-pihs-thuh-gli-fuhs
Oviparous δ-vihp-ear-uhs
Oviposition δ-vih-pδ-zihsh-uhn
Parietal eye pah-ri-eh-tahl ι
Paucicameral lung paw-se-kahm-ar-ahl luhng
Plastron plahs-truhn
Pleurodont teeth ploor-uh-dohnt teth
Preferred optimal temperature zone prih-fard ohp-tuh-muhl tehm-par-uh-char zδn
Proteroglyphous prδ-tar-uh-glι-fuhs
Septum sehp-tuhm
Sexual dimorphism sehck-shu-ahl dι-mawr-fihz-uhm Spectacle spehck-tah-kuhl
Solenoglyphous sohl-en-δ-glι-fuhs
Splenopancreas spleh-nδ-pahn-kre-ahs
Tail autonomy tal aw-tohn-uh-me
Thecodont teeth the-kuh-dohnt teth
Thrombocyte throhm-bδ-sιt
Unicameral lung u-nih-kahm-ar-uhl luhng
Vitellogenesis vι-tehl-lδ-jehn-eh-sihs
Viviparous vι-vihp-ear-uhs
Vomeronasal organ vohm-ar-δ-na-zahl ohr-gahn
INTRODUCTION
There are more than 4000 species of amphibian and 7700 species of reptile found on the earth today.
Amphibians and reptiles, collectively referred to as herptiles, comprise an extremely varied group of animals, and it is difficult to make many generalizations regarding their form and function. This chapter covers some of the more interesting and clinically applicable features of the anatomy and physiology of amphibians and reptiles.TAXONOMY
The class Reptilia includes four orders:
1. Crocodylia (alligators and crocodiles).
2. Squamata (snakes and lizards).
3. Chelonia (turtles and tortoises).
4. Rhyncocephalia (tuataras).
Only a relatively few species within a number of different orders are commonly seen in captivity (Table 22-1).
The class Amphibia is made up of three orders:
1. Gymnophiona (caecilians).
2. Anura (frogs and toads).
3. Caudata (salamanders and newts).
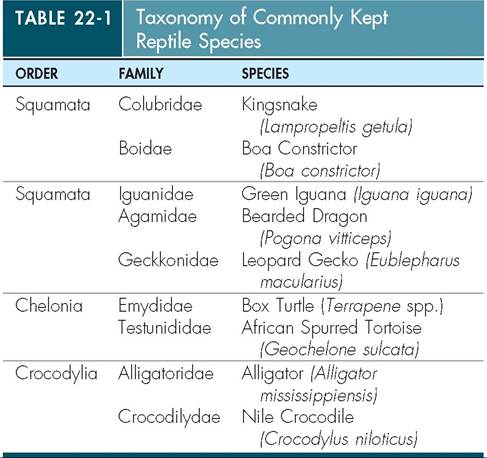
Most people are familiar with the general body structure of anurans and salamanders, but may have never seen a caecilian. Caecilians are legless and have elongated bodies much like a snake or eel. They are found in both terrestrial and aquatic habitats. As with reptiles, only a relatively few species are commonly kept in captivity (Table 22-2).
METABOLISM
Amphibians and reptiles are commonly referred to as ecto- thermic, or cold-blooded. Ectothermic animals are unable to generate body heat internally, therefore their body temperatures are dependent on environmental temperatures. Other ectothermic animals include fish and invertebrate species. Some reptiles are able to raise their body temperatures via metabolic processes. The two notable examples are
Leatherback Sea Turtles, which are able to generate body heat internally, and pythons, which elevate their body temperature via muscular contractions when incubating eggs.
Herptiles maintain their body temperatures in an appropriate range through a process called behavioral thermoregulation.
Through behavioral thermoregulation, amphibians and reptiles are able to regulate their body temperatures precisely according to metabolic need. Movements within the thermal gradient of their habitat, as well as postural changes, enable herptiles to adjust their body temperatures as needed. For instance, many herptiles bask in the sun to elevate body temperature, and will seek shade when they need to cool down. A snake trying to conserve body heat will coil tightly to decrease its surface area and consequently heat loss, whereas a hot snake will uncoil to accomplish the opposite. Some animals, such as chameleons, can actually adjust the color and pattern of their skin to increase or decrease the absorption of thermal energy from the sun. All of the aforementioned are examples of behavioral thermoregulation.Ectothermy is directly related to the energy conservation strategies employed by amphibians and reptiles. Compared to mammals, herptiles are incredibly energy efficient. They are able to manipulate their body temperatures behaviorally to match changing metabolic needs. For instance, a quiescent herptile will usually choose to maintain its body temperature in the low end of its preferred range, whereas an animal digesting a meal will likely choose higher temperatures to support the metabolic demands of digestion. In some pythons, the gastrointestinal tract and associated organs, such as the liver and pancreas, actually fluctuate in size drastically depending on food intake. This allows the animal to conserve energy by not supporting a large tissue mass that is not in use.
An ectotherm's ability to thermoregulate effectively is entirely dependent on access to temperatures within the animal's preferred optimal temperature zone (POTZ). The POTZ is a range of temperatures in which the animal can perform all necessary metabolic functions. This principle becomes very important when maintaining herptiles in captivity. If not provided with an appropriate thermal gradient, the herptile will not be able to thermoregulate efficiently.
When provided with inappropriately low temperatures, ectothermic animals can suffer from digestive problems, immunosuppression, and other disorders. If kept at temperatures that are too high, the animal is forced to maintain a high metabolic rate and may suffer from energy deficits.The efficiency of ectothermy allows many herptiles to survive on very small amounts of food. Some sit-and-wait predators, such as large pythons, may only consume a few large meals during the course of a year. Insectivorous herp- tiles can survive on minute fractions of what would be required to maintain a bird of equal size.
A disadvantage of ectothermy is that activity of herptiles is limited by the availability of appropriate environmental temperatures. For this reason, herptiles that reside in temperate areas go through periods of hibernation, or bruma- tion, when temperatures drop out of the range in which the animal can remain active. Herptiles that are hibernating do not feed and are minimally active until environmental temperatures return to the POTZ.
TEST YOURSELF 22-1
1. What are the four orders of living reptiles? What are the three orders of amphibians?
2. Define ectothermic.
3. What is a preferred optimal temperature zone?
4. How does an ectothermic animal maintain an appropriate body temperature?
CLINICAL APPLICATION
Drug Administration
It is important that ectothermic animals be kept within their preferred optimal temperature zones when receiving drug therapy. If a drug is administered to an ectothermic animal kept outside its POTZ, it may not be absorbed, distributed, metabolized, or eliminated as expected. This can result in ineffective treatment if the drug is not properly absorbed and distributed. Toxicity can occur when metabolism or elimination is affected. Toxicity is also possible when an animal receives a number of doses at suboptimal temperatures, and the drug pools at the injection sites or in the gastrointestinal tract instead of being absorbed systemically.
When this occurs, the animal may effectively receive a toxic dose when it is warmed and therefore capable of absorbing the medication.INTEGUMENT
The skin of reptiles is keratinized and varies in appearance according to species. The integument has three components: the subcutaneous space, dermis, and epidermis. The subcutaneous space is typically limited in most reptiles, and the skin is relatively inelastic, making administration of subcutaneous fluids difficult in comparison to most domestic mammals. The dermis is made up of dense connective tissue and contains blood and lymph vessels, nerves, and chro- matophores, which are pigment-containing cells. Chro- matophores allow some lizards, such as chameleons, to change their skin color and pattern drastically. Some lizards and crocodilians have bony plates within the dermis called osteoderms that provide protection. Most reptiles have distinct scales that are formed by a folding of the epidermis. Scales can vary greatly in size and shape. The epidermis can form unique structures, such as crests, tubercles, spines, and dewlaps. Instead of eyelids, snakes have modified scales that cover the eyes; these scales are called the spectacle, or brille (Figure 22-1).
Scale and scute nomenclature is important for species identification and medical recording (Figure 22-2). Different forms of keratin are found in the scales and the interscalar skin. Alpha keratin is softer and more flexible and is found
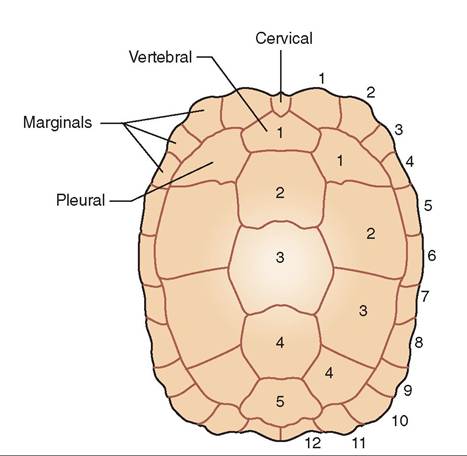
FIGURE 22-2 Chelonian carapace illustrating nomenclature of scutes. (Redrawn from O'Malley B: Clinical anatomy and physiology of exotic species, New York, 2005, Elsevier Saunders.)

FIGURE 22-1 The spectacle, or brille, of a Ball Python (Python regius).

FIGURE 22-3 Milksnake (Lampropeltis triangulum) with recently shed skin.
Snake skin is usually shed in one piece.in the interscalar skin. Beta keratin is relatively rigid and is found in the scales.
One of the unique and most clinically important features of the reptilian integument is the process of ecdysis, or shedding of the skin. Ecdysis occurs as a reptile grows and in response to skin injury. The process of ecdysis is under the control of the thyroid gland. Some reptiles shed the skin completely in one large piece, such as snakes and some lizards (Figure 22-3), whereas others shed in a piecemeal fashion. Many species of lizard consume the dead skin, or exuvia. The shed cycle is initiated by cellular replication in the epidermis, followed by secretion of lymph containing enzymes between the old and new epidermal layers. This results in dulling of the skin color and opacification of the spectacle in snakes (Figure 22-4). The dullness and opacification resolve as the lymph is reabsorbed a few days prior to

FIGURE 22-4 Opacification of the spectacle in a Burmese Python (Python molurus) indicating impending ecdysis.
actual shedding of the old skin. Actual shedding of the skin is typically accomplished by mechanical rubbing on objects in the environment.
The epidermis of amphibian skin typically has a single or very few layers of keratinized cells, which makes it extremely permeable. Aquatic amphibians have no keratinized cells in their epidermis. The dermis contains chromato- phores and many glands that produce secretions that help protect the amphibian's skin. Some glands within the dermis and epidermis produce toxic secretions that are important defense mechanisms for animals that have them (Figure 22-5).
The dermis is firmly attached to the underlying musculature and bone in salamanders and caecilians, making subcutaneous space minimal to nonexistent. Anurans have more significant amounts of subcutaneous space, because the attachments of the dermis are much looser. Amphibians regularly shed the outer layers of the epidermis and usually consume the exuvia.

FIGURE 22-5 Parotid gland in a Colorado River Toad (Bufo sp.). The gland is the raised oval structure caudal to the eye.
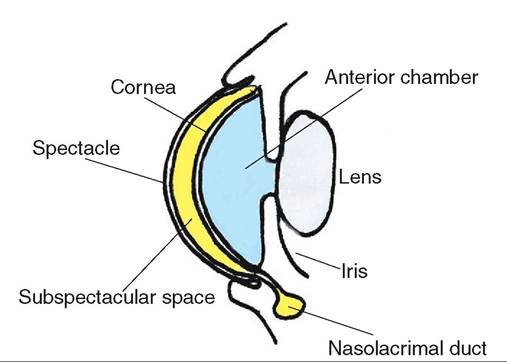
FIGURE 22-6 Cross section of the anterior portion of a snake eye.
The extremely permeable nature of amphibian skin allows them to absorb all the water they need from the environment, thus amphibians do not drink. Areas of increased permeability, referred to as drink patches, are usually present on the ventral surfaces that most frequently come in contact with water.

Dysecdysis
Dysecdysis, or abnormal shedding, can occur for a variety of reasons. Sometimes the cause is as simple as low relative humidity in the environment or a lack of appropriate cage features to rub on to begin the shed. Shedding difficulties can also be caused by dehydration due to systemic illness, dermatitis, skin wounds, and malnutrition. Dysecdysis is frequently caused by external parasites, such as the snake mite (Ophionyssus natricis).
Occurrence of dysecdysis should prompt a review of husbandry parameters and the correction of any problems. Most cases can be resolved simply by soaking the reptile in a container of warm water for an hour or two. Most of the adhered skin will typically fall off following the soak, or can be removed manually with ease. The removal of retained spectacles in snakes should be attempted with caution. Typically, retained spectacles can be removed with a cotton-tipped applicator or masking tape following soaking.
Failure to correct cases of dysecdysis can result in dermatitis, dyspnea caused by obstructed nares, and continued problems with future sheds. Annular rings of retained shed skin around the toes and tails of lizards can result in ischemic necrosis and sloughing of tissues.
VISION
There are many important differences between mammalian and herptile eyes. Even among the amphibians and reptiles, there are significant anatomic differences between families.
Reptiles are similar to birds in that the iris is made up of skeletal muscle and is under voluntary control. Therefore, the mydriatic agents used to dilate the pupil for ophthalmic examinations in mammals are not effective in reptiles. A direct pupillary light response is usually noted, but consensual reflexes are often not seen.
PERIOCULAR STRUCTURES
Some species of lizard, especially those inhabiting sandy, dry areas, have lower eyelids that are very thin. The relative transparency of these lids allows for some vision even when the eyelids are closed. In most species, the lower lid is more mobile than the upper. Crocodilians are the exception, because their upper lids are more mobile. Like birds, some reptiles have cartilaginous pads, or tarsal pads, in their eyelids.
The third eyelid, or nictitans, is very well developed and mobile in a variety of reptiles. Crocodilians have nictitans that they use to protect the eyes while swimming under water and procuring prey. Snakes and lizards that do not possess true eyelids do not have nictitans.
Snakes and some species of lizard do not possess true eyelids. Instead these species have a clear, fused scale called the spectacle, or brille, which covers and protects the eye. The tear film is retained between the cornea and the spectacle in the subspectacular space. The tears drain into the mouth through the nasolacrimal duct system (Figure 22-6).
Most reptiles have poorly developed extraocular muscles, therefore the globe tends to be relatively immobile. The chameleons are exceptions in that they have well-developed extraocular muscles. These extraocular muscles allow the eyes to move freely and independently of one another.
Chelonians do not possess nasolacrimal ducts. The tears spill over the lid margin, so some degree of epiphora is normal in healthy animals (Figure 22-7).

FIGURE 22-7 African t5∣^ι^rr^d Tortoise (Geochelone sulcata). A small amount of epiphora is normal in this tortoise owing to the lack of a nasolacrimal duct system.
called the conus papillaris. Tlts con us extends into the vitre- ouumsohr from the optic disc and is probably involved in putrroivtiodning n and removing waste. Crocodilians have
a tapetum, whereas other reptiles do not.
Tuataras and some lizards have a well-developed parietal eye Ioedyd dorsally on the head. This structure has a rudi- emtiennatary r and cornea but no iris, lids, or musculature. oAncnection exists between the parietal eye and sections of trhaienb, including the pineal gland. The exact function of itehtealpar eye is unknown, but it is thought to play a role in liaht-ckclc-mediated hormone function.
TEST YOURSELF 22-3
1. Which reptiles hαveα tapetum?
2. How dees the reptile iriseiffetfrom tSet ot mammals? Why is this clinically important?
3. The nesolαcrimαldhct systemis abssnt i nweiat order of reptiles?
Most reptiles possess both lacrimal glands and harderian glands. Typically the harderian gland lies medial to the eye in the rostral aspect of the orbit, whereas the lacrimal gland is Tcpiyally in a caudodorsal position. Both glands produce secretions that combine to form the tear film. Only the harderian gland is found in snakes.
The eyelids develop at metamorphosis in most salamanders, frogs, and toads. Eyelids are absent in some species of completely aquatic frogs and salamanders. Caecilians' eyes are covered with skin in a manner somewhat analogous to tehcteacslpe in snakes. The presence of periocular glands cvcaoriredsinag to species. Only the harderian gland is
present in anurans and caecilians, whereas some salamanders hwe both lacrimal and harderian glands.
fyTehse e o many amphibian species protrude ventrally irnalto the o cavity when the animal swallows. The move- fment o the eyes helps propel food from the oral cavity into the esophagus. Animals that have lost eyes may have diffi- lcouwltiynsgw. al
THE GLOBE AND INTRAOCULAR STRUCTURES
Most reptiles possess scleral bones, or ossicles, the exceptions ebseing snak and crocodilians. The ossicles help the eye maintain its shape and provide protection from trauma.
The Iots is more fluid in reptiles than in mammals, espe- ciallyin chelonians, in which accommodation is performed by the Iots being squeezed through the pupil. The lens is more rigid in snakes, and the ciliary bodies are very closely associated with the root of the iris. Accommodation in snakes is accomplished by the lens moving back and forth as a result of changes in pressure within the aqueous and vitre- uoumsohrs.
All reptiles have avascular retinas. All nutrition and waste irtehminoval w the eye is performed by choroidal vessels or modified vessels that protrude into the vitreous humor. Lizerds possess a structure analogous to the avian pectin

CLINICAL APPLICATION
Hypovitaminosis A
Diets deficient in vitamin A can result in abnormal cell growth within epithelial tissues. With hypovitaminosis A, the epithe- eial ceUs tome flattened and lose their ability to perform normal functions. This change is called squamous metaplasia, and et frequently affects epithelial tissues associated with the lnadcrimal a harderian glands. The metaplastic cells can no longer produce a normal tear film and sloughed cells build up abnstdruoct the glandular ducts. Damage to the cornea and conjunctival tissues can occur because of decreased tear pro- dnudction a bacterial or fungal infection. Treatment involves andydressing a secondary infections that may have occurred aonrrde ccting the vitamin A deficiency, but caution should
be used, because vitamin A toxicity commonly occurs in che- looiaoe treated with parenterally delivered vitamin A prepara- iteiotanrsy. D supplementation is usually a safer alternative. Also, ia it important to keep in mind that the functions of other organs may be disrupted, because other epithelial tissues fcefaenctbed,a including those in the kidneys and liver.

Red-Eared Slider Turtles (Chrysemys scriptae). The turtle on the left suffers from hypovitaminosis A. Notice the closed, swollen eyelids as compared to the normal turtle on the right.
CARDIOVASCULAR SYSTEM
The location of the heart within the body cavity varies according to species. In chelonians, the heart lies on the midline just caudal to the thoracic girdle and ventral to the lungs. The heart in most lizards lies within the thoracic girdle. The hearts of some lizards, such as monitors, tegus, and crocodilians, lie farther back in the coelomic cavity, and some are almost in the middle. Cardiac location varies in snakes according to species, but usually the heart is found at the junction of the first and second thirds of the animal's body length. Snakes' hearts are fairly mobile within the coelomic cavity, which facilitates the ingestion of large prey.
The cardiac structure of reptiles is significantly different from that of mammals. Most reptiles have three-chambered hearts, with two atria and one common ventricle (Figure 22-8). However, the single ventricle functions as a fourchambered heart, and therefore oxygenated and deoxygenated blood rarely mix. Three regions exist within the ventricle and are functionally separate: the cavum venosum, cavum arteriosum, and the cavum pulmonale. The cavum pulmonale receives blood from the right atrium and directs flow into the pulmonary circulation. The cavum arteriosum receives blood from the pulmonary veins and directs oxygenated blood to the cavum venosum. The paired aortic arches arise from the cavum venosum and lead to the systemic circulation.
Differential blood flow and separation of oxygenated and deoxygenated blood are maintained by pressure differences in the outflow tracts and a muscular ridge that partially separates the cavum venosum and cavum pulmonale (Figure 22-9). In times of oxygen deprivation, such as when some reptiles dive or when snakes consume large prey, reptiles can shunt blood away from the lungs. Right-to-left cardiac shunting is facilitated by an increase in pulmonary vascular
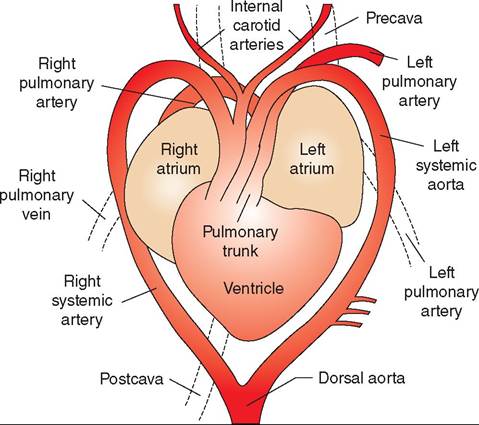
FIGURE 22-8 Ventral view of a lizard heart. (Redrawn from Mader D: Reptile medicine and surgery, ed 2, St Louis, 2006, Saunders.)
resistance. Resumption of breathing results in a decrease in pressures within the pulmonary vasculature and a restoration of pulmonary blood flow.
Crocodilians are the only reptiles that possess fourchambered hearts comparable to those of mammals and birds. Even so, crocodilian cardiac anatomy is quite different from what is seen in birds and mammals. Crocodilians possess two aortas, the right arising from the left ventricle and the left from the right ventricle. Both aortas route blood to the systemic circulation. The right and left aortas are connected near the base of the heart by the foramen of Panizza. The foramen allows blood from the right ventricle to bypass the pulmonary circulation when necessary. A valve exists at the opening of the pulmonary artery that has interdigitating muscular projections, hence the commonly used name cog-wheel valve. When the animal holds its breath, the cog-wheel valve closes, and blood that would have normally entered the pulmonary circulation is diverted into the left aorta and the systemic circulation.
The amphibian heart is three-chambered and similar in anatomy and function to that of snakes, lizards, and chelo- nians. The interatrial septum is fenestrated in caecilians and most salamanders; in anurans, it is not.
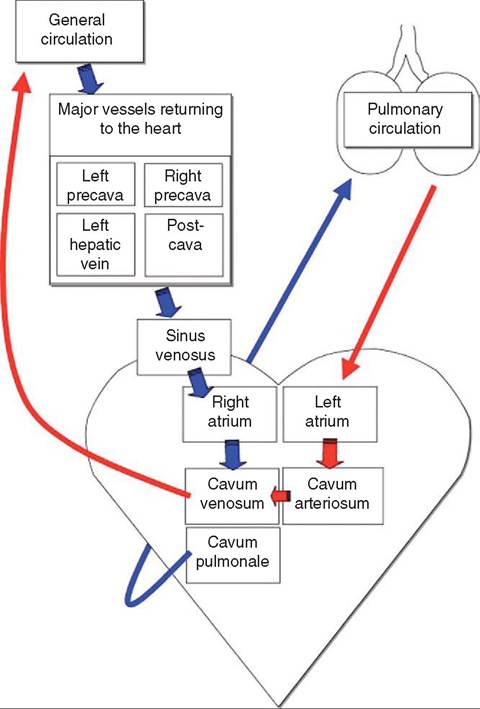
FIGURE 22-9 Diagram of blood circulation in the noncrocodilian heart. (From Mader D: Reptile medicine and surgery, ed 2, St Louis, 2006, Saunders.)
The heart rate of reptiles depends on species, size, temperature, activity level, and metabolic function. An equation employing metabolic scaling for determination of the “appropriate” heart rate in reptiles has been proposed:
Heart rate = 33.4 ? (weight in kilograms-0'25)
This equation assumes that the reptile is within its preferred optimal temperature zone.
Lizards possess a vasovagal reflex that can be induced by applying gentle pressure to both eyeballs through closed lids. Pressure on the eyeballs causes a drop in heart rate and blood pressure, and induces a catatonic state from which the lizard easily recovers upon cessation of the pressure and mild stimulation.
Cardiac monitoring can be difficult in reptiles, but direct cardiac movement is easily observed in most snakes and lizards with caudally positioned hearts. When the animal is placed on its back, the motion of the ventral scutes or scales is usually visible. Auscultation is possible but difficult owing to the low-amplitude sounds produced by the reptilian heart. Placement of a thin, moist towel between the bell of the stethoscope and the animal can help cut down on incidental noise as the scales scrape against the bell. Electrocardiography and cardiac ultrasound can also be employed, but the lack of normal values and methods for standardized examination makes the interpretation of results difficult.
The vascular anatomy of herptiles is similar to that of mammals and birds. Reptiles and amphibians are unique in that they have very well developed lymphatic systems with large lymph vessels found near arteries and veins. Large dilations of the lymph vessels occur roughly where lymph nodes would be found in mammals, though reptiles have no lymph nodes. Instead, the walls of some lymph vessels contain smooth muscle that actively pumps lymph through the vasculature; these structures are known as lymph hearts. The presence of the large lymph vessels in association with blood vessels results in relatively frequent lymph contamination of a sample during venipuncture.
BLOOD
Reptilian red blood cells are oval in shape, nucleated, and larger than those of mammals (Figure 22-10). Erythropoiesis takes place in the bone marrow, and the erythrocytes contain hemoglobin, which allows them to carry oxygen to tissues. The reptilian erythrocyte is relatively long lived, with a life span in some species of 600 to 800 days. Immature erythrocytes are occasionally seen in the circulation of normal animals, especially in juveniles or in animals undergoing ecdysis.
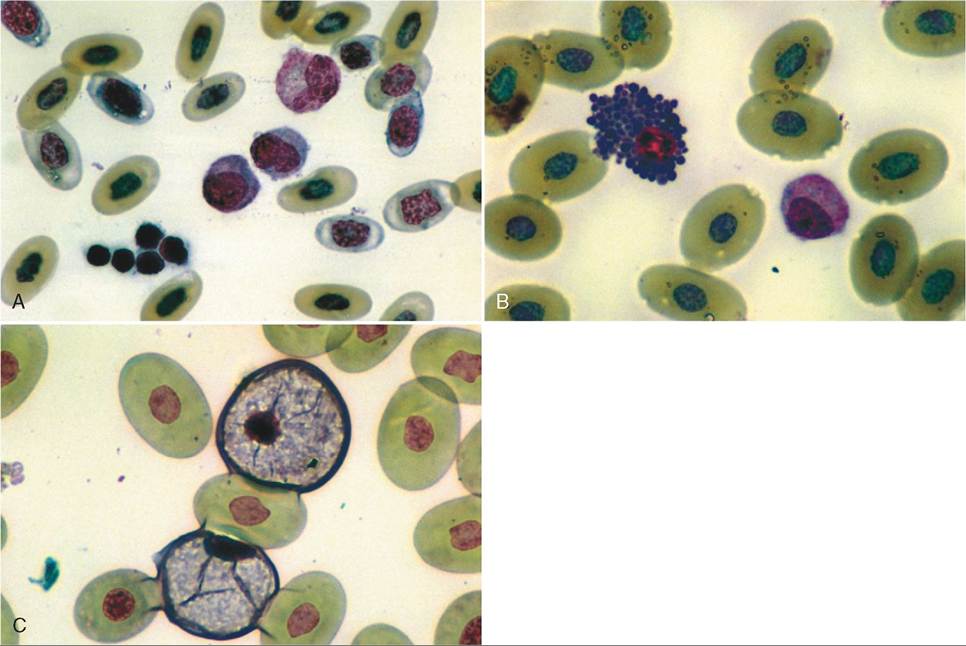
FIGURE 22-10 Reptilian blood smears. A, Nucleated polychromatic erythrocytes, lymphocytes, thrombocytes, and an azurophilic monocyte. B, Basophil and azurophilic monocyte. C, Heterophils.
Heterophils are analogous to mammalian neutrophils. They differ from neutrophils in that they do not possess peroxidase and acid phosphatase, which are enzymes involved in breaking down necrotic material. Given that reptiles lack these enzymes, they produce caseous pus instead of liquid material. Heterophils are involved in the phagocytosis of foreign materials. Reptilian heterophils are very similar in appearance to avian heterophils; the cells are typically round with eosinophilic rod-shaped granules in the cytoplasm, and the nuclei are round to oval in shape.
Eosinophils are easily confused with heterophils. They are similar in appearance but differ in that the cytoplasmic granules are round instead of rod shaped. In some chelonians, eosinophils may account for over 20% of circulating white blood cells.
Basophils are small, round cells with deep blue cytoplasmic granules. These granules can often obscure the nucleus. Like eosinophils, basophils can normally make up a large percentage of circulating leukocytes in normal chelonians.
Lymphocytes can vary in size, and large and small lymphocytes are found together. Lymphocytes are usually round and have large nuclei. Lymphocytes typically have a large nucleus-to-cytoplasm ratio and no cytoplasmic granules. In most reptiles, the lymphocyte is the predominant leukocyte in circulation.
Monocytes are the largest leukocytes in the reptile circulation. The nuclei may be oval or lobed. The cytoplasm in monocytes is typically blue-gray and may contain small vacuoles or fine granules. The monocytes of some reptiles, namely snakes, may have small azurophilic granules in the cytoplasm and therefore are sometimes referred to as azurophils. Because these cells are simply a monocyte variant, they are more appropriately labeled azurophilic monocytes.
Thrombocytes perform the same function as mammalian platelets. Thrombocytes are small oval nucleated cells with clear cytoplasm. The cytoplasm is usually colorless, but may contain small granules.
Clinical application
Blood Collection
Blood collection can be very difficult in amphibians and reptiles, because there are usually no externally visible vessels, therefore it is important to know vascular anatomy to be successful in obtaining usable samples. The vessel most commonly used for blood collection in reptiles is the ventral coccygeal vein, which courses on the midline just ventral to the vertebrae of the tail. This vessel is accessible in snakes, lizards, crocodilians, and salamanders.
Obtaining a blood sample from chelonians presents additional challenges because of the presence of the shell. A variety of options exist, and which one to use depends on the size and species of the turtle or tortoise. The jugular vein, dorsal coccygeal vein, and brachial vein are all used with success, as is the subcarapacial sinus formed by the confluence of the common intercostal veins and the caudal branch of the external jugular vein.
Cardiac puncture can be performed safely on most reptiles and amphibians as long as they are securely restrained or anesthetized. Other options for blood collection include the ventral abdominal vein in amphibians and lizards and the postoccipital sinus in crocodilians and chelonians.
Respiration in reptiles is driven by oxygen levels (PO2) in the blood. This is in contrast to mammals, in which CO2 levels are the primary determinants of respiratory rate and tidal volume.
The glottis of most amphibians and reptiles is easily located in the rostral portion of the oral cavity (Figure 22-11); it is found behind the tongue in all species, and in snakes it is very mobile. The mobility of the snake glottis makes it capable of protruding from the mouth, allowing respiration during ingestion of prey. The glottal opening, which is only open during respiration, is bordered by paired arytenoid cartilages. Amphibians and reptiles do not have vocal cords and are therefore only capable of limited
TEST YOURSELF 22-4
1. How many cardiac chambers do noncrocodilian reptiles and amphibians have?
2. Right-to-left cardiac shunting serves what function in reptiles?
3. What is the main difference between a heterophil and a neutrophil?
4. What is the function of a lymph heart?
RESPIRATORY SYSTEM
The anatomy and physiology of the reptilian respiratory system differ significantly from those of mammals. Reptiles are capable of functioning with very low oxygen levels, accounting for many unique features. Owing to their relatively large pulmonary volume, efficient anaerobic metabolism, and cardiac shunting capabilities, reptiles are capable of surviving for long periods of time without breathing.
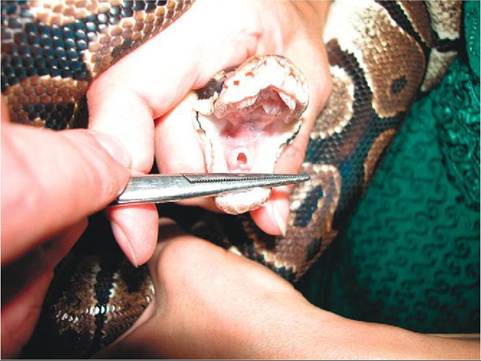
FIGURE 22-1 1 Open glottis in a Ball Python (Python regius).
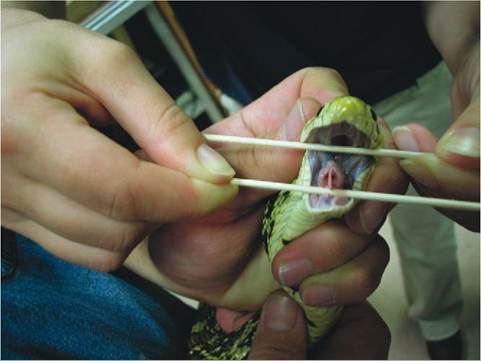
FIGURE 22-1 2 Bullsnake (Pituophis melanoleuca) glottis showing the preglottal keel.
vocalization: all species hiss; some grunt, such as chelonians and crocodilians; and some, like crocodilians, bellow. The calling sounds produced by frogs and toads are produced by vocal sacs that arise from the trachea. Movement of air in and out of the vocal sacs produces the sound. Some species of snake possess a glottal keel (Figure 22-12), which increases the volume of vocalizations.
Chelonians posses complete cartilaginous tracheal rings, whereas the rings of all other reptiles are incomplete. The tracheal bifurcation occurs in the cervical area of chelonians, but it is lower in other reptiles and is usually found around the base of the heart.
Three distinct lung structures are found in reptiles (Figure 22-13). The most primitive is the unicameral lung, which is found in snakes and some lizards. The unicameral lung is a scilmikeple, sa structure. The cranial portion of the unicameral lung typically contains the tissues involved in gas exchange, and the relatively avascular caudal portion is comparable to the avian air sac. The multicameral lung is found in chelonians, some lizards, and crocodilians. The multicam- eral lung is divided into many compartments and possesses intrapulmonary bronchi. An intermediate lung structure, the paucicameral lung, shares characteristics of both the unicameral and multicameral lungs. Paucicameral lungs are found most notably in iguanas and chameleons.
The lungs of amphibians are typically simple, saclike structures with little if any partitioning. Some species of salamander do not have lungs at all and rely entirely on cutaneous respiration.
The pulmonary tissue of amphibians and reptiles grossly has a honeycomb appearance (Figure 22-14). The openings of the honeycomb terminate in gas-exchange structures called faveoli. Unlike mammalian alveoli, the faveoli are fixed structures that do not expand or contract. The faveoli are surrounded by capillary beds, where the blood takes up oxygen and releases carbon dioxide.
Reptiles do not have true diaphragms, therefore they must rely on different methods of respiration. Most
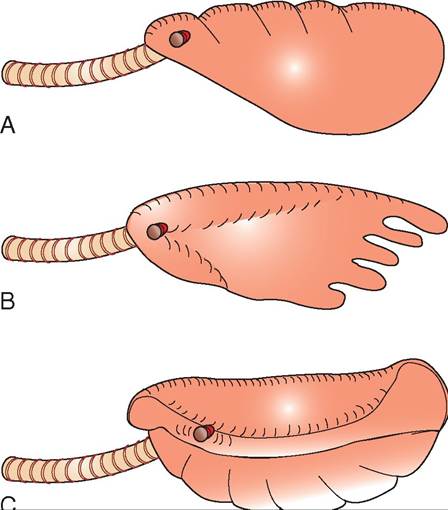
FIGURE 22-13 Different lung structures found in reptiles. A, Unicameral. B, Paucicameral. C, Multicameral. (Redrawn from O'Malley B: Clinical anatomy and physiology of exotic species, New York, 2005, Saunders.)
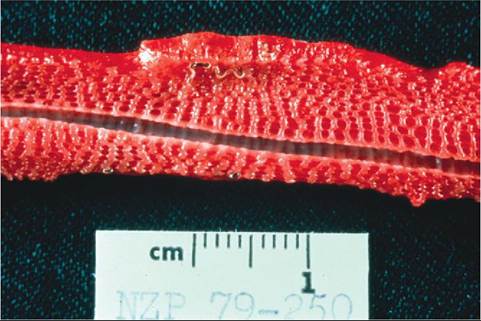
FIGURE 22-14 Inner surface of a snake lung showing the honeycomb structure. The intrapulmonary trachea is seen running the length of the section. A lungworm is also present.
chelonians and some lizards possess membranous separations between the lungs and the other coelomic viscera that are somewhat analogous to the mammalian diaphragm, although these membranes are not directly involved in the respiratory cycle. Reptiles rely on the action of the intercostal muscles and other parts of the axial musculature to perform the active phases of respiration.
A unique variation in respiration is utilized by crocodilians, which respire through the function of a hepatic piston. Crocodilians have a muscular septum caudal to the lungs that is somewhat analogous to the mammalian diaphragm, and the cranial aspect of the liver is attached to this septum. The diaphragmaticus rsιu^le attaches the caudal aspect of the liver to the pubis. Contraction of the diaphragmaticus moves the postpulmonary septum caudally through the liver, resulting in inflation of the lungs.
Pumping of the buccal cavity and pharynx provides the primary means of pulmonary ventilation in amphibians. Gas exchange can also occur across the mucous membranes of the buccal cavity and pharynx. Many aquatic chelonians are also capable of exchanging gases across the mucous mem- fbranes o the pharynx and cloaca.
OM World chameleons have an air sac that arises from tehnetrval aspect of the trachea in the cervical region. This structure is probably involved in behavioral displays and does not have gas-exchange capabilities. Snakes are unique in that most only have a single right lung. The left lung is present in some species, such as boas and pythons, but is reduced in size. A tracheal lung is present in some snake species and is presumed to allow for some degree of gas euxrcinhgange d ingestion of prey.
TEST YOURSELF 22-5
1. Which reptiIeshave complete tracheal rings?
2. Where doesgaa exchangetakeplaae inthe reptilian lung?
3. Describe the three different lung structures found in reptiles.
EARS AND HEARING
seThe ear ar found on both sides of the head and caudal to tyhees e in most amphibians and reptiles. The tympanum is ebasseirlvyeod in most species, but lies in a depression and may be covered by folds of skin in some lizards and crocodil- ians. Life birds, reptiles have a single bone in the middle ear called the columella. Tlie columella connects to the tympanum and the quadrate bone, and it transmits vibrations to itvnhadel oow w of the cochlea, which is part of the inner ear. Vibrations transmitted from the columella to the inner ear are; converted to nerve impulses that travel to the brain evsiatibthuelovcochlear nerve.
Reptiles possess semicircular canals comparable to those found in mammals and birds; these fluid-filled canals control balance and equilibrium.
Sonakes d not have external ears; their columella articu- lytes distally with the quadrate bone. Articulation of the ictohlumella w the quadrate bone allows snakes to be sensi- troiovuentd g vibrations that are transmitted through the mandibles. Snakes are also able to hear aerial sounds, especially those of lower frequency. Salamanders and caecilians amrewsohat like snakes in that they do not have tympanic
membranes and their columella is sometimes degenerate.
Snakes, fr^gs, toads, and some lizards possess endolymphatic sacs associated with their middle ears; these sacs function as calcium-secreting glands. Some, for example thooe of geckos in the genus Phelsuma, are so large they pass
through the skull and bilaterally into the cervical region. lTahnedsse g are involved in maintaining calcium homeo- tsitcausilsa,rlpyar during egg formation and metamorpho-
shiisb, iiannas.mp

TEST YOURSELF 22-6
1. W hat Ss the he tsegf tbeninglebone hontth within the middle eer of e reptile?
2. Which Ieotilhsdo noi neveextemαleαr openings?

CLINICAL APPLICATION
Aural Abscesses
Middle ear infections are Irequently encountered in both wild and captive chelonians. These infections result in the accumulation of caseous purulent material within the middle ear that results in outward bulging of the tympanum (see figure). The abscess can be unilateral or bilateral and is sometimes larger than the animal’s head.
Aural abscesses are likely to result from the ascension of bacteria frail the oral cavity into the middle ear through the auubde.itory t A variety of bacteria are commonly cultured from these abscesses. Predisposing factors include unsanitary captive conditions and nutritional deficiencies, namely hypovitaminosis A. Hypovitaminosis A results in squamous metaplasia of epithelial surfaces and can affect the middle ear, making it more susceptible to infection.
eTqreuaitrmesent r incising the tympanum and removing the caseous material, followed by thorough flushing. After surgery has been performed, antibiotic therapy, continued flushing, and elimination of predisposing factors complete trhape yt.he
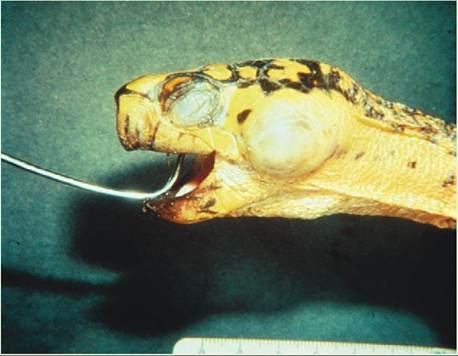
Aural abscess in a Box Turtle (Terrapene sp.).
GASTROINTESTINAL TRACT
Reptiles are remarkable in the wide variety of feeding strate- gmiepsloeyed across the class. Carnivorous, omnivorous, rabnidvohreous reptiles all exist. Snakes and crocodilians are sntirvioctresc,ar whereas all three feeding strategies are
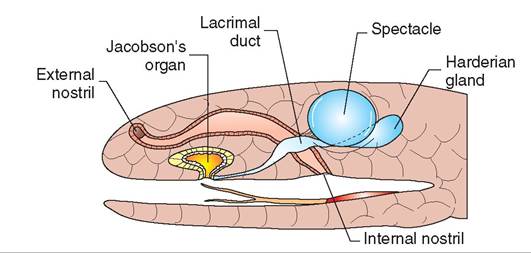
FIGURE 22-15 Diagram of a snake head illustrating the structure of the vomeronasal organ. (Redrawn from O'Malley B: Clinical anatomy and physiology of exotic species, New York, 2005, Saunders.)
represented among the lizards and chelonians. All adult amphibians are carnivores as well. Some animals have highly specialized diets and only consume specific foods or prey. Examples include snakes such as the King Cobra (Ophiopha- gus hannah), which preys exclusively on other snakes; the Caiman Lizard (Dracaena paraguinesis) feeds only on snails. It is important to know these specialized dietary requirements when attempting to maintain a species in captivity. If incorrect food is offered, acceptance may be poor and nutritional deficiencies can occur.
Snakes and lizards, such as monitors and tegus, have tongues that are deeply forked and function as a particle delivery system for the vomeronasal organ. The vomeronasal organ is an accessory olfactory organ (Figure 22-15). The forking of the tongue allows detection of particle gradients across the sampled area, enabling animals to follow scent trails efficiently. Most lizards have fairly mobile tongues, which they use frequently to taste items they encounter. Chameleons have highly specialized projectile tongues designed for capturing prey at long distances. The tongue is long and has a sticky end that sticks to the prey. Muscular connections between the tongue, a specialized hyoid apparatus, and the sternum allow the tongue to be extended to lengths longer than the body in most chameleons. Turtles and tortoises typically have thick, fleshy tongues that are relatively immobile. The Alligator Snapping Turtle has a uniquely adapted tongue, which resembles a small pink worm. The turtle wiggles this tongue with the mouth open to attract prey. Crocodilians have immobile tongues that are attached along their entire length to the intermandibular space. Croc- odilians also have muscular flaps arising from the base of the tongue and dorsal pharynx that allow them to open their mouths while submerged without ingesting or inhaling water.
Tongue structure varies according to species in amphibians. All species, except aquatic anurans, salamanders, and caecilians, use the tongue for capturing prey. Anurans and most terrestrial salamanders grasp food through a process called lingual flipping, in which the caudodorsal aspect of the fleshy tongue is flipped forward into a cranioventral position (Figure 22-16). Sticky secretions on the surface of the tongue cause the prey to adhere, allowing it to be pulled into the
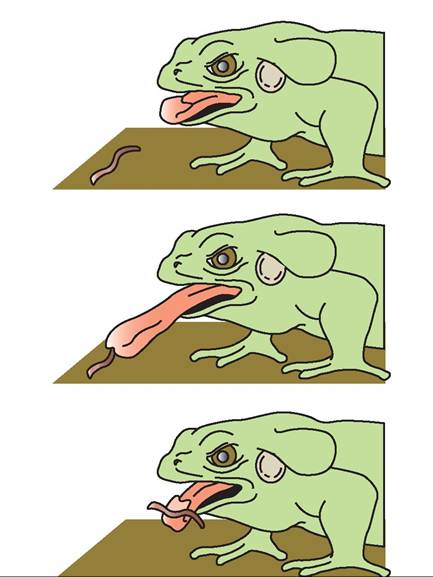
FIGURE 22-16 Lingual flipping mechanism for prey capture in an anuran. The sequence shows how the tongue unfolds and the rostroventral surface adheres to the prey.
mouth. Lungless salamanders are able to project their tongues much like chameleons when feeding.
There are numerous salivary glands in the oral cavities of most reptiles. The salivary secretions facilitate the ingestion of large prey by providing lubrication. As in mammals, the saliva also has enzymatic properties that aid digestion. The venom glands of some snakes are modified salivary glands and are found in the upper jaw below the eyes. In venomous lizards, members of the genus Heloderma, the venom glands are found along the lateral aspects of the lower jaws.
The dentition of reptiles varies significantly according to family. Turtles and tortoises have no teeth but possess keratinized beaks similar to those of birds, called tomia. Among the other reptiles, three types of dentition exist: thecodont, pleurodont, and acrodont (Figure 22-17). Thecodont teeth, in which the teeth arise from sockets in the skull bones, are found only in crocodilians. Acrodont teeth, in which the teeth are fused to the biting edges of the mandible and maxillae, are found in some species of lizards, such as those in the family Agamidae. Pleurodont teeth are attached to the periosteum on the medial aspects of both the mandibles and maxillae. Snakes and iguanid lizards have pleurodont dentition. Thecodont and pleurodont teeth are replaced periodically during the life of the animal, whereas acrodont teeth do not grow back if lost or broken.
Snakes are unique in that they possess six rows of teeth: two on the mandibles, two on the maxillae, and two on the palatine/pterygoid bones (Figure 22-18). Venomous snakes
FIGURE 22-18 Vontrel e sneko skull illustretinn tto nexillere and aeletino/atorenoia tooth. (From O'Melloe B: Clinical enetome and atesiolone oI oxotic saocios, Now York, 2005, Seunaorsi)
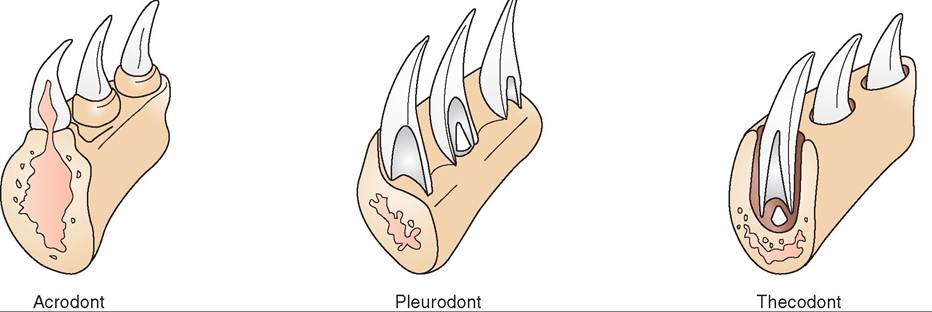
FIGURE 22-1 7 Illustration of thedifferent typesof reptil e dentition.(FromO'Malley EnCIinicaI anatomy and physiology of exotic saocios, Now York, 2005, Seunaors.)
have specialized dentition for venom delivery and are divided into three categories: Solenoglyphous snakes, or vipers; proter- oglyphous snakes, or elapids; and opisthoglyphous snakes, or rear-fanged colubrids. Proteroglyphous and opisthoglyphous snakes hwe fangs that are fixed in an upright position. The fangs of Solenoglyphous snakes Md against the roof of the mouth when the mouth is closed and are moved into an erect position when the snake bites. Nonvenomous snakes are rederred to as aglyphous, meaning they are without fangs. The venom-delivering teeth are comparable to
hypodermic needles in that they are hollow with an opening near the end through which the venom is expelled. The fang reencoemives v through a duct from the venom gland at its base. When delivering a bite, muscles surrounding the lvaendom g contract to force venom out through the
fang.Venomous snakes are capable of controlling the amount of vendm delivered with each bite. The primary function eonf osmnak v is for procurement of food; its use for defense is secondary. It should also be noted that enzymes found in snake venom perform an important role in the fdigestion o prey.
With the notable exception of bufonid toads, most amphibians have teeth. Caecilians and salamanders have lbaortyh maxil and mandibular teeth, and some species have palatal tee th. Maxillary dentition occurs sporadically in eacniuers,an sp and only one species is known to have mandibular teeth. Some species of frog have well- developed cutting plates on their rostral mandibles called the odontoid process.
eTphteiliran esophagus is relatively thin and distensible
ienciems,ost sp especially in those adapted for consuming
large prey. Some reptiles have unique esophageal structures that reflect the type of prey they consume. Sea turtles have conical projections that line the esophagus, and these aid in swallowing slippery food items.
oTmheascth varies in size and shape according to species and is involved in both chemical and mechanical digestion onfoakdfe. s S have stomachs that are very distensible but rooftsesnly not g distinct from the esophagus and duode-
oncuomd.ilIiancsr, the body of the stomach is made up of thick mnscle comparable to that of the avian gizzard. Like that of mammals, the pH of the reptilian stomach is usually quite acidic, ranging between 2 and 3 in healthy animals. Hydrochloric acid, pepsinogen, and other enzymes are involved in the chemical digestion of food held in the stomach.
hTihbeiaanmp gastrointestinal tract is relatively short and simple. The esophagus is very short and wide, especially in anurans. Anurans are capable of prolapsing their stomach through the mouth and cleaning it off after they have ingested an undesirable item. Gastric prolapse can also be seen with some methods of anesthesia or as a terminal event in dying animals. Unlike the intestinal tract in mammals, the amphibian intestinal tract is not easily divided into distinct regions. The liver and gallbladder are usually found close together, and the livers of amphibians often contain aggregations of melanin-containing macrophages called melano- macrophages. These cells are involved in immune function and can give the liver a mottled black appearance. The amphibian liver performs the same functions as in other vertebrates. The pancreas is usually found between the stomach and the proximal segments of intestine.
The structure of the reptilian intestinal tract is extremely variable, according to species and, consequently, diet. Herbivores tend to have longer intestinal tracts than carnivores, and omnivores fall somewhere between the two. Snakes have relatively straight intestinal tracts with only minor winding found in the small intestine. The colon can be large and complex in herbivorous species that rely heavily on hindgut fermentation for digestion. In some species, such as the Green Iguana, the colon is partitioned as an adaptation for more efficient hindgut fermentation. Cecae are found in herbivorous lizards and chelonians and also serve as a site for hindgut fermentation. The cecum is absent or rudimentary in carnivorous chelonians, lizards, snakes, and crocodilians.
The liver in reptiles is usually quite large and consists of at least two lobes in most species. The exceptions are snakes, which have a large, elongated, single-lobed liver. Typically the liver makes up approximately 3% to 4% of the body weight in lizards and snakes, although the size can vary with season, nutritional status, and other factors. Most reptiles have a gallbladder, which is variable in size and position; the gallbladder in snakes is found caudal to the liver, near the spleen and pancreas. Blood is supplied to the liver by the hepatic artery and portal vein.
The reptilian liver performs the same functions as the mammalian liver; it excretes bile for digestion, performs metabolic degradation, and produces various substances. Normally, the liver can be dark brown to black in color. Certain metabolic states, such as vitellogenesis, can cause a normal change in color of the liver. For example, during vitellogenesis the liver can take on a yellow color due to fat accumulation. This should not be misinterpreted as pathologic hepatic lipidosis. As in amphibians, reptilian livers also often contain large numbers of melanomacrophage centers, which can result in a mottled appearance.
The pancreas is typically found within the mesentery in close proximity to the stomach or duodenum. In snakes, the pancreas is found near the pylorus of the stomach near the gallbladder and spleen. In some snake species and in chelo- nians, the spleen and pancreas are combined to form a single organ, called the splenopancreas. The pancreatic ducts enter the duodenum in most species. In chelonians, the pancreatic ducts and the bile ducts enter the pyloric region of the stomach.
The reptilian pancreas performs both exocrine and endocrine functions. As in mammals, the pancreas produces a variety of digestive enzymes, including but not limited to amylase, chymotrypsin, trypsin, and chitinase. The exact makeup of pancreatic secretions is dependent on species and diet, and the function of enzymes depends on appropriate temperature. Pancreatic islets are found throughout the pancreas in reptiles and contain glucagon-secreting alpha cells, insulin-secreting beta cells, and other cells capable of secreting somatostatin and pancreatic polypeptide. Glucagon and insulin are involved in the maintenance of blood glucose levels in mammals, but it is important to note that blood glucose levels in reptiles are much more variable because of the fluctuations in metabolic rate related to ectothermy.
Figures 22-19 through 22-22 illustrate the gastrointestinal tract and other aspects of internal anatomy of a chelonian, lizard, snake, and frog, respectively.
TEST YOURSELF 22-7
1. What is the function of the vomeronasal organ?
2. Name the different types of dentition found in reptiles.
3. Which amphibians do not have teeth?
4. What herptiles are capable of gastric prolapse as a method of emptying their stomachs?
CLOACA
All amphibians and reptiles possess a common outflow tract for the gastrointestinal and urogenital tracts called the cloaca. The cloaca is made up of three chambers: the copro- deum, urodeum, and proctodeum (Figure 22-23). In some species, the cloacal chambers are distinctly separated by muscular sphincters; in other species, they are less discrete. The coprodeum is the most cranial chamber and receives the rectum. The urodeum is the middle chamber, where the urogenital openings, urethra, and male reproductive organs
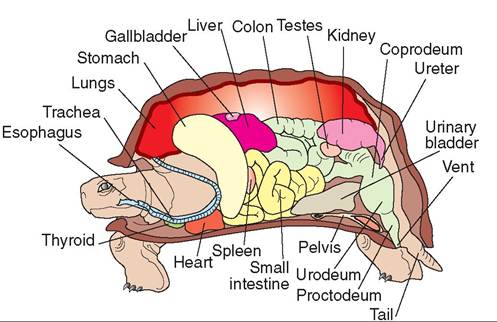
FIGURE 22-19 Cross section showing the internal anatomy of a chelonian. (Redrawn from O'Malley B: Clinical anatomy and physiology of exotic species, New York, 2005, Saunders.)
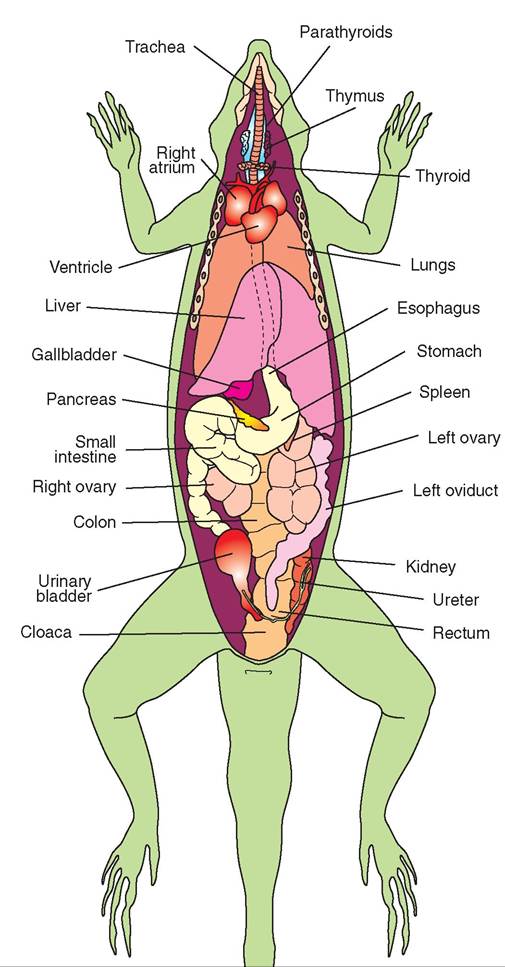
FIGURE 22-20 Ventral view of a female lizard showing internal anatomy. (Redrawn from O'Malley B: Clinical anatomy and physiology of exotic species, New York, 2005, Saunders.)
of crocodilians and chelonians are found. The proctodeum is the last chamber before the vent and often contains glandular tissue.
KIDNEYS
The kidneys of reptiles differ significantly in structure and function from those of mammals. In crocodilians, turtles, and most lizards, the kidneys are oblong, smooth-surfaced structures found in the caudal coelomic cavity. Snakes possess kidneys that are lobulated and grossly resemble a stack of melted coins (Figure 22-24). The ureters empty into the dorsolateral aspect of the urodeum instead of the urinary bladder. The urinary bladder, if present, is connected to the urodeum by a short, broad urethra. The urine enters the bladder for storage through the urodeum. It is important to note that crocodilians, snakes, and some species of lizard do not possess urinary bladders. When the bladder is absent, any storage of urine occurs in the cloacal chambers or the rectum.
Reptile kidneys do not have a distinct renal pelvis. Instead, the distal tubules of the nephrons join into collecting ducts, which eventually come together and form the ureter. Unlike those of mammals, the reptile nephron does not have a loop of Henle, so reptiles are incapable of producing urine more concentrated than their blood. When water conservation is necessary, reptiles are able to absorb additional water from the urine through the wall of the urinary bladder, rectum, or cloaca, which results in voided urine that is somewhat concentrated.
Following the distal tubule of the nephron, a sexual segment is found, the cells of which enlarge when high levels of reproductive hormones are in circulation. These sexual segments are believed to add secretions to the seminal fluid in male reptiles, but their exact function is unknown.
Other unique physiologic characteristics allow reptiles to conserve water very efficiently, even though they are unable to produce concentrated urine. The first is the production of uric acid as a protein waste product in terrestrial reptiles. Uric acid is relatively insoluble in blood and is voided via secretion from the renal tubules. Because uric acid is secreted rather than filtered, very little urine is needed to excrete it from the body. (Production of uric acid is also functionally important for embryos in the egg, because it is relatively insoluble and nontoxic, therefore relatively large amounts can be stored in the egg without adverse effects.) Aquatic or semiaquatic reptiles produce ammonia, urea, or both as a waste product; these are much more soluble but also more toxic than uric acid. The aquatic environment allows these reptiles to pass large amounts of urine to eliminate waste products without risk of dehydration.
The second characteristic that allows water conservation is the presence of a renal portal system (see Chapter 18). The renal portal veins arise from the veins of the tail and hindlimbs, if such are present. The renal portal vessels lead to capillary beds; these perfuse the renal tubules but bypass the glomeruli. This anatomic configuration allows the reptile to discontinue blood flow to the glomeruli and decrease urine production during times of dehydration, while maintaining blood flow to the renal tubules. By maintaining blood flow to the tubules, ischemic necrosis is avoided and uric acid secretion continues.
Following excretion by the kidneys, uric acid combines with minerals to form potassium or calcium salts. These uric acid salts, or urates, are passed with the feces and urine and appear as a white, chalky substance (Figure 22-25).
Amphibian kidneys are somewhat more primitive than reptilian kidneys. In addition to filtering the blood, amphibian kidneys also filter coelomic fluid via openings called nephrostomes that connect the coelomic cavity to the renal
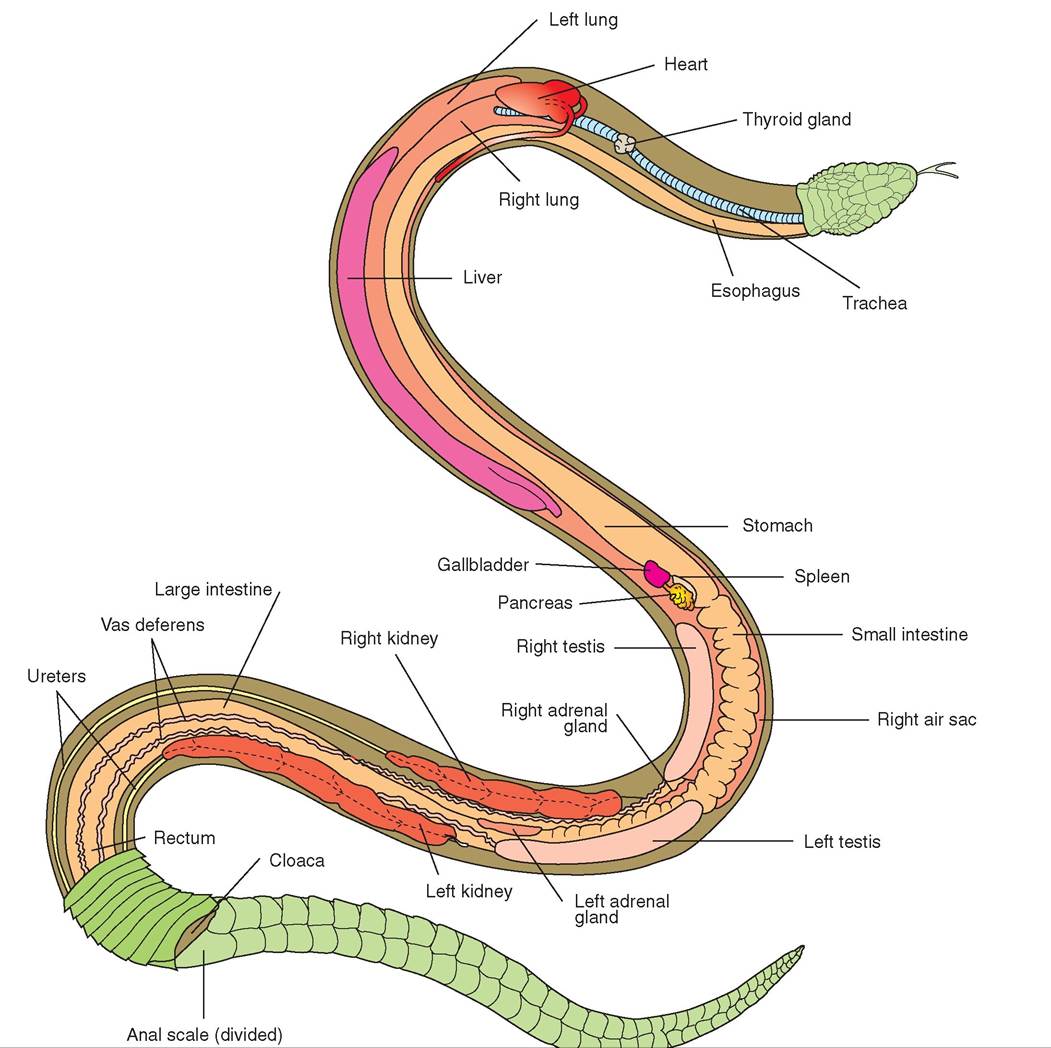
FIGURE 22-21 Ventral v law ef a ma lesnal22-23 Illustration of the distal colon, cloaca, bladder, and kidney of a lizard. (From O'Malley B: Clinical anatomy and physiology of exotic species, New York, 2005, Saunders.)
TEST YOURSELF 22-8
1. Given that reptiles are unable to produce concentrated urine, how do they conserve water?
2. What nitrogenous waste products are produced most frequently by aquatic amphibians? What nitrogenous waste products are produced by terrestrial amphibians?
3. What is the purpose of the renal portal system?
4. Where do the ureters terminate in reptiles and amphibians?
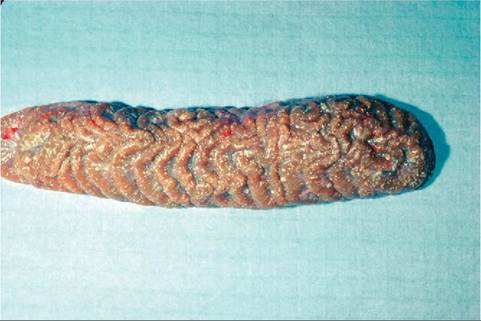
FIGURE 22-24 Snake kidney. The white mottling is due to uric acid deposits; this snake suffered from gout.

FIGURE 22-25 Urates from a snake.
cava, aorta, and adrenal glands. The ductus deferens leads from the testes to the dorsal wall of the urodeum. Most male reptiles also have a sexual portion to the kidney tubules, which develops in response to high levels of circulating sex hormones. These areas of the renal tubules probably provide secretions that contribute to the seminal fluid.
Male reptiles all possess copulatory organs, and fertilization is internal. The copulatory organs of reptiles vary in structure according to family. Crocodilians and chelonians have a phallus that originates in the floor of the cloaca. This phallus is composed of erectile tissue and forms a trough, through which semen is transmitted to the female's cloaca. The phallus has no function in urination. Male snakes and lizards possess paired structures, called hemipenes, that can be everted from the tail base through the vent (Figure 22-26). These hemipenes also form a trough through which semen is carried and serve no purpose in urination.
FEMALE ANATOMY
All female reptiles have paired ovaries, which are found in the dorsal coelomic cavity in close proximity to the caudal vena cava, aorta, and adrenal glands. The paired oviducts
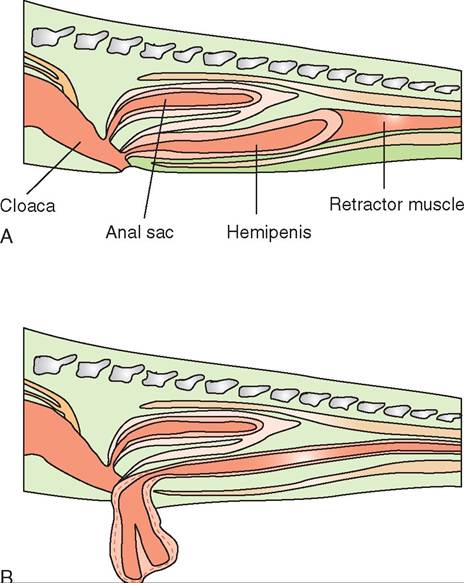
FIGURE 22-26 Cross section of a snake tail showing the hemipenes in both the resting (A) and everted (B) positions. (From O'Malley B: Clinical anatomy and physiology of exotic species, New York, 2005, Saunders.)
lead to the cloaca and terminate at the genital papillae in the dorsal wall of the urodeum (Figure 22-27). The oviducts are divided into five regions: the infundibulum, magnum, isthmus, uterus, and vagina. Ova from the ovaries are released to the oviducts during ovulation. The albumen and shell are added in the oviduct in oviparous species prior to egg laying. The fetuses are retained in the uterine portion of the oviduct for development in viviparous species.
REPRODUCTIVE CYCLE
The quiescent ovary is covered with small, inactive follicles. When certain environmental cues are encountered, estrogen and follicle-stimulating hormone (FSH) are released; this causes development and maturation of the follicles via a process called vitellogenesis, which is the process of adding vitellogenin, a lipid substance, to the yolk within the developing follicle. Vitellogenin is formed by the liver from mobilized fat, and during vitellogenesis the liver often takes on a yellow color. Large amounts of calcium are also added to the yolk during this process. Circulating calcium levels tend to be very high in reptiles during vitellogenesis, because calcium is mobilized by bone to supply the developing follicles.
Following maturation of the follicles, a surge of luteinizing hormone (LH) causes ovulation, and the ova are released from the follicles and taken up into the oviducts. In the oviducts, the albumen, shell membranes, and shell are laid down prior to the egg being laid. In all viviparous
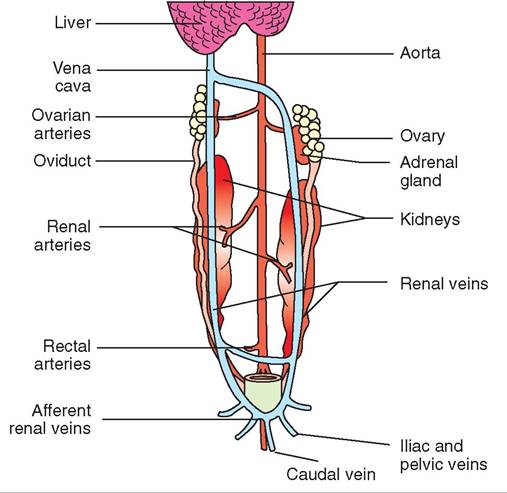
FIGURE 22-27 Diagram illustrating the reproductive anatomy of a female lizard. (From O'Malley B: Clinical anatomy and physiology of exotic species, New York, 2005, Saunders.)
reptiles, some degree of support of the fetus is continued by the female, and some lizards possess true placentas.
Oviposition
Many reptiles dig nests and therefore need suitable substrate to lay eggs. If not provided with suitable nesting material, some female reptiles will be reluctant to lay and may experience dystocia. Most snakes readily make use of enclosed egg-laying chambers lined with moist substrate. Chelonians and lizard species, such as iguanas and chameleons, will often excavate deep holes to deposit their eggs in; they should be provided with suitable nesting material when oviposition is imminent.
Few reptiles show any interest in their eggs or offspring after oviposition or parturition. Exceptions include crocodil- ians, which protect their nests and provide protection for the young for a period of time following hatching. Pythons and cobras protect their nests until hatching.
EGG INCUBATION
The anatomy of the reptilian egg is comparable to that of the avian egg. Refer to Chapter 21 for a description of the avian egg.
Successful hatching of eggs requires appropriate environmental conditions in the nest. Temperature, humidity, and gas composition of the environment must be within acceptable ranges for healthy embryos to develop. The substrate on which eggs are incubated must retain an appropriate amount of moisture without encouraging growth of bacteria and fungi (Figure 22-28). Incubation time and temperature vary according to species; incubation time can range from a few weeks to well over a year. Typically, eggs will hatch faster if

FIGURE 22-28 Incubating lizard eggs in vermiculite substrate.
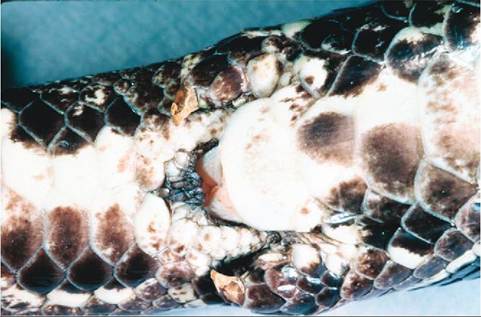
FIGURE 22-29 Pelvic spurs of a male African Rock Python (Python sebae).
incubated at temperatures at the high end of the acceptable range. Some species of chelonian require a diapause during incubation for the embryos to develop normally and proceed to hatching. A diapause occurs when incubation temperatures drop and development stops for a period of time; development resumes when temperatures rise again.
Reptile eggs do not need to be rotated during incubation, unlike bird eggs. The embryonic membranes and shell membranes form attachments within a few days of oviposition that solidly anchor the developing embryo within the shell. Repositioning eggs following development of these attachments can destroy the embryo.
SEX DETERMINATION
Sex in reptiles is determined by either the genotype or the temperature at which the eggs are incubated. Sex chromosomes are not present in species such as crocodilians, chelo- nians, tuataras, and some lizards, therefore egg incubation temperature determines the sex of the hatchling. In general, higher incubation temperatures produce males and lower temperatures produce females in crocodilians and lizards, but the opposite is true for chelonians. In nature, there is typically a temperature range within the nest that allows the production of a mixed clutch of male and female hatchlings. Reptiles that possess sex chromosomes differ from mammals in that the females are the heterozygous sex (ZW) and males are homozygous (ZZ).
SECONDARY SEXUAL CHARACTERISTICS
Snakes do not tend to show significant sexual dimorphism. Many times, the tail is longer and thicker in males owing to the presence of the hemipenes. In boids, pelvic spurs can be larger in males; pelvic spurs are remnants of the pelvic limbs, and these are used by males to provide tactile stimulation to females during courtship (Figure 22-29).
Male chelonians often have a concave plastron, which allows closer apposition of the cloacas when the male mounts the female. The vent is also located in a more distal position in males for the same reason. Other, less reliable, keys to sexing chelonians are eye color, carapace shape, and the length of claws on the forelimbs.
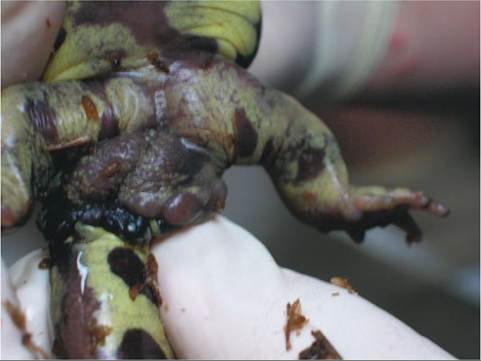
FIGURE 22-30 Cloacal glands in a male Tiger Salamander (Ambys- toma tigrinum). The glands are the bulging tissue behind the legs at the base of the tail.
Some lizards show significant dimorphism that is readily apparent, even to the untrained eye. A good example is the Jackson's Chameleon (Chameleo jacksonii); the males have three well-developed horns on their faces, and the females have none. In Green Iguanas, the males are larger with more pronounced dewlaps and crests. Hemipenal bulges can also be detected in the tail base of some male lizards. Iguanids and geckonids have femoral or precloacal pores that are frequently more developed in males.
Crocodilians sometimes display significant differences in size as adults, and the males are clearly larger than the females.
AMPHIBIAN REPRODUCTION
Sexual dimorphism is present in many amphibians. In some species of poison dart frogs, the males have enlarged toe pads. The tympanic membranes are larger in some male anurans. Male salamanders possess prominent cloacal glands (Figure 22-30) that are visible around the vent. Some male anurans and newts develop rough nuptial pads on their limbs in response to testicular hormones. These nuptial pads help the male grasp the female during fertilization. Male anurans will also call when conditions are appropriate.
Amphibians have paired gonads, which are found in the dorsocaudal coelomic cavity. Male toads possess ovarian remnants associated with the testes called the Bidder’s
organs.
The majority of caecilians are viviparous and most anurans and salamanders are oviparous. Internal fertilization occurs in caecilians and most salamanders. Caecilians evert a portion of their cloaca, the phallodeum, to deposit semen into the female's cloaca. Male salamanders deposit packets of sperm called Spermatophores onto substrate; these are subsequently picked up by the female salamander's cloaca. Female salamanders can store sperm for extended periods of time in pockets within the cloaca called Spermato- theca. In anurans, fertilization is external; male frogs and toads fertilize the eggs as they are laid, while grasping the female in an embrace called amplexus.
Amphibian eggs are usually deposited in or near water, because larvae, amphibian young, are typically water dependent. Some anurans and most salamanders display some form of parental care of eggs or young. Many amphibians that display parental care will carry the eggs or young on their backs, on their legs, or in pouches found on the dorsal or ventral surfaces.
Larval anurans, or tadpoles, lead completely aquatic lives prior to metamorphosis. Tadpoles have round bodies and laterally compressed tails. During metamorphosis, tadpoles lose their gills and tails and develop limbs. Metamorphosis is usually complete within 90 days in most species. Some species, such as the Bullfrog (Rana catesbiana), are capable of delaying metamorphosis for up to 2 years. Metamorphosis is stimulated by thyroid hormones.
Some larval salamanders have forms similar to those of tadpoles, but they do not lose their tails when they change to their adult form. Many species of salamander bypass the larval stage and hatch in their adult form. Some species of salamander, such as the Axolotl (Ambystoma mexicanum) are neotenic, which means they retain their larval form into adulthood.
TEST YOURSELF 22-9
1. How is sex determined in reptiles that do not possess sex chromosomes?
2. What are the names of the male copulatory organs in snakes and lizards?
3. Describe the process of fertilization in salamanders.
4. What is vitellogenesis?
ENDOCRINE SYSTEM
With the exception of lizards, all reptiles have a single thyroid gland. It is tan in color and found ventral to the trachea and cranial to the great vessels of the heart. In crocodilians, the thyroid is bilobed with a thin isthmus in between. Lizards have variable thyroid gland structure among the various
CLINICAL APPLICATION
Preovulatory Follicular Stasis and Egg Binding
Many factors are involved in stimulating and maintaining normal reproductive function in reptiles. These factors include nutrition, temperature, humidity, light cycle, and suitable egg-laying environments. Disruption of normal function can occur when inappropriate conditions occur in captive settings.
Preovulatory follicular stasis occurs when large, mature follicles develop on the ovaries but ovulation fails to occur. These mature follicles take up space within the coelomic cavity and can cause decreased feed intake owing to impingement on the stomach. Also, retained follicles are prone to infection, which can lead to sepsis. Preovulatory follicular stasis is usually corrected by surgical removal of the ovaries (ovariectomy).
Egg binding can be caused by all of the aforementioned factors, but most frequently it seems to be the result of inadequate calcium balance due to poor nutrition or conditions that result in mechanical obstruction of the oviducts. If no obstructions are present, oviposition can be encouraged by providing appropriate environmental conditions and nesting substrate, as well as correction of any nutritional deficits. If all conditions are considered normal, and the animal is healthy, oxytocin can be used effectively to stimulate oviduct contractions and oviposition in most chelonians. The effectiveness of oxytocin is variable, but is generally poor, in other reptile species. Surgical removal of the eggs or the reproductive tract should be pursued when oviduct or cloacal obstruction occurs, or in animals unresponsive to conservative therapy.
species, and the gland can be seen as diffuse tissue in the cervical region. The size of the thyroid gland varies according to season and metabolic state. Thyroid hormone is involved in the process of ecdysis and growth.
Also found in the cervical region are the parathyroid glands and ultimobrachial bodies. These glands are separate from the thyroid and can be difficult to find grossly. The parathyroid glands secrete parathormone and the ultimo- brachial bodies produce calcitonin. Both hormones are involved in calcium and phosphorus homeostasis (see Chapter 11).
The adrenal glands are usually found in the ligaments that suspend the gonads (mesorchium or mesovarium) and are therefore in close proximity to the gonads. The exceptions are chelonians and crocodilians, in which the adrenals lie against the kidneys. The adrenal glands are usually red or yellow and can become enlarged with stress. Reptilian adrenal glands produce the same hormones as avian glands: epinephrine, norepinephrine, aldosterone, and corticosterone. There is no distinct separation between the adrenal cortex and the medulla in reptilian adrenal glands.
The exact location of the various endocrine organs in amphibians varies greatly according to species. In general, the form and function of these organs are comparable to what is observed in reptiles. Thyroid hormone is responsible for stimulating metamorphosis and ecdysis in amphibians.
CLINICAL APPLICATION
Nutritional Secondary Hyperparathyroidism
Nutritional secondary hyperparathyroidism occurs frequently in herbivorous or omnivorous reptiles that receive diets deficient in calcium or vitamin D, or in animals not provided with access to adequate ultraviolet radiation. In response to low calcium availability, the parathyroid glands secrete high levels of parathormone, which results in resorption of calcium from the skeletal system, retention of calcium by the kidneys, and increased intestinal absorption of calcium. These effects maintain normal blood calcium levels, which are necessary for life. If the nutritional or environmental deficits are not corrected, bones become depleted of minerals, are weakened, and are prone to pathologic fractures. In an effort to provide support to the limbs, fibrous connective tissue develops in place of bone, in a condition called fibrous osteodystrophy.
Hypocalcemia can develop if the bones become completely depleted of calcium. Clinical signs associated with hypocalcemia include constipation, anorexia, muscle weakness, muscle tremors, and seizures. Many animals can recover if provided with supportive care, appropriate environmental conditions, and an appropriate diet.
NERVOUS SYSTEM
The neuroanatomy of reptiles follows the basic vertebrate design with a central nervous system consisting of a brain and a spinal cord. There are three major divisions within the reptilian brain: the forebrain, which comprises the olfactory lobes, cerebral hemispheres, and a diencephalon; the midbrain, made up of the optic lobes, cerebral peduncles, and nerve fibers connecting the hindbrain to the forebrain; and the hindbrain, which is the cerebellum and medulla oblongata. Peripheral nerves provide motor and sensory innervation.
The reptilian brain is Iissencephalic, meaning there are no gyri or sulci present on the surface. In terms of evolution, reptiles were the first group to have a cerebral cortex separated into hemispheres and to have a brain offset from the spinal cord and extending back over the diencephalon.
The two meninges in reptiles are the pia-arachnoid layer and the dura mater. The pia-arachnoid layer is vascular and lies directly on the surface of the brain and spinal cord. The dura mater lies over the pia-arachnoid layer and is relatively avascular.
Unlike mammals, reptiles do not have a cauda equina, and the spinal cord extends to the tip of the spine.
Reptiles are the lowest vertebrates to have 12 cranial nerves; fish and amphibians have only 10. The spinal nerves of reptiles are also advanced beyond those of fishes and amphibians, and only sensory fibers are found in the dorsal roots and motor fibers in the ventral roots.
Reptiles depend more on spinal segmental reflexes and locomotion centers for the control of movement than they do cerebral stimulation. As a result, body movements are more autonomous in reptiles than in mammals and birds. Because of their spinal autonomy, reptiles with spinal cord injuries usually have a much better prognosis than do mammals with comparable lesions.
The amphibian nervous system is more primitive than that of reptiles. Amphibian brains are well adapted to support basic functions such as sight, olfaction, and movement, but they have very limited integrative capabilities. The spinal cord extends to the tip of the tail in amphibians and caecilians. In frogs and toads, the spinal cord ends in the lumbar region and a cauda equina is present. Much like reptiles, amphibians are heavily dependent on spinal segmental reflexes to control movement.
MUSCULOSKELETAL SYSTEM
SKULL
There is extreme variation in the skulls of the different reptile orders. The different types are divided into two general categories: anapsid and diapsid (Figure 22-31). Che- lonians possess anapsid skulls, which do not have temporal openings. All other reptiles possess diapsid skulls, which means they do have temporal openings in the skull. Unique skulls are found in the lizards and snakes, which have skulls that are designed to have significant flexibility. This flexibility allows for ingestion of large prey. Snake skulls in particular are extremely mobile, because the mandibular symphysis is not fused in snakes, rather it is connected by ligaments. This flexible connection of the rostral mandibles allows the
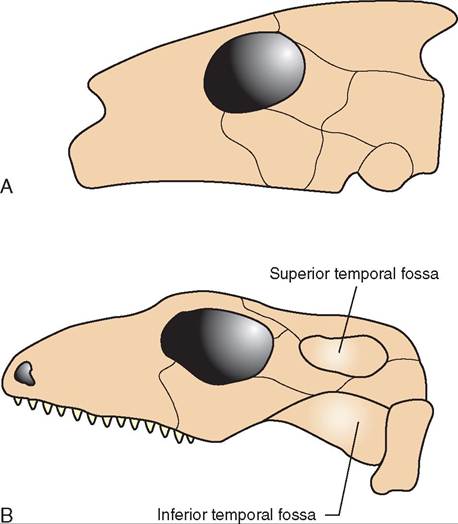
FIGURE 22-31 Illustration of anapsid and diapsid skull types. A, Anapsid skulls are found in chelonians. B, Diapsid skulls are found in crocodilians, tuataras, snakes, and lizards.
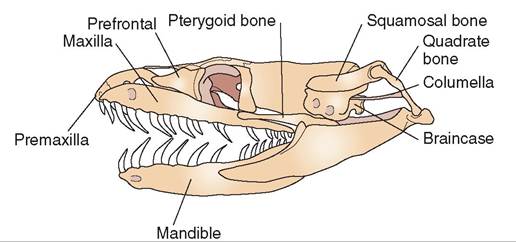
FIGURE 22-32 Skull of a nonvenomous snake. (Redrawn from O'Malley B: Clinical anatomy and physiology of exotic species, New York, 2005, Saunders.)
jaws to move independently of one another and literally “walk” forward when prey is ingested. Other adaptations include the presence of quadrate bones, which allow the mouth to open very wide, and movable bones that make up the maxillae and palate (Figure 22-32).
The skulls of anurans are typically broad and fenestrated, with fewer bones than those of other amphibians. The parts of the skull involved with olfaction and hearing are better developed than they are in salamanders and caecilians, but the palate is poorly developed, and the dentition is reduced.
Caecilians have compact, well-ossified skulls, and the dentition is well developed. Salamander skulls generally represent an intermediate between caecilian and anuran skulls.
AXIAL SKELETON
With the exception of chelonians, the spines of reptiles are extremely flexible in comparison to those of mammals. This is possible because reptilian spines are not required to support significant amounts of weight, as most reptiles spend the majority of their time with their bellies in contact with the ground. Unlike mammals, it is not possible to divide the vertebral regions of reptiles into cervical, thoracic, lumbar, sacral, and caudal regions. Instead, the terms presa- cral, sacral, and caudal are used in referring to reptilian vertebral regions.
Another unique feature of the reptilian axial skeleton is the presence of a single occipital condyle that forms the articulation between the skull and spine. This allows for increased mobility of the head on the spine but also makes this connection relatively fragile, necessitating care when restraining reptiles. Most reptiles have well-developed ribs that help support the body wall and assist in respiration. The exceptions are chelonians, in which the spine and ribs are fused to the bony shell.
The bony shell of chelonians is made up of the dorsal carapace and the ventral plastron. The bone is covered externally by keratinized scutes. Some chelonians, such as Box Turtles, have hinged plastrons that allow them to withdraw into their shells and close up completely for protection.
Many lizard and salamander species are capable of losing their tails via a defensive mechanism called tail autonomy. Tail autonomy allows lizards to drop their tails when being pursued or attacked by a predator. The discarded tail usually continues to wiggle about, which is intended to draw the attention of the attacker while the lizard makes its escape. The tails of lizards capable of autonomy have fracture planes along which they break. When tails are lost this way, there is usually little or no blood loss, because the muscles of the tail contract strongly, and the blood vessels are occluded. The lost tail can regenerate with a stiff, cartilaginous rod that replaces the spine in the tail. In lizards, the regenerated tail is usually smaller and stiffer than the original. The regenerated tails of salamanders usually closely resemble the original.
The number of vertebrae found in amphibians varies greatly. Caecilians can have up to 250, salamanders from 30 to 100, and anurans from six to nine. With the exception of the first cervical vertebra, the atlas, the vertebrae of caecilians are indistinguishable. In salamanders and anurans, the vertebrae are somewhat more differentiated, because these animals also possess sacral vertebrae. In anurans, the vertebrae are fused, and the last caudal vertebra is called the urostyle. Ribs are well developed in caecilians, poorly developed in salamanders, and absent in anurans. Caecilians lack a sternum, whereas it exists as a small cartilaginous plate in salamanders. Anurans have a well-developed, ossified sternum.
APPENDICULAR SKELETON
The skeletal structures of reptilian limbs are comparable to those of mammals, with a few exceptions. In reptiles, the pectoral girdle consists basically of a scapula and coracoid bone, which have muscular attachments to the body. The bones of the pectoral limbs articulate with the scapula and coracoid bones and consist of a humerus, radius, ulna, carpal bones, metacarpal bones, and phalanges. The pelvis of a reptile articulates with the sacral vertebrae and forms the connection between the spine and pelvic limb. The pelvic limb consists of the femur, tibia, tarsal bones, metatarsal bones, and phalanges. Reptiles typically have five digits on both front and rear feet. Exceptions occur, such as in crocodilians, which have only four digits on the hind feet. In most reptiles, the hindlimbs are longer than the forelimbs.
Snakes, obviously, are devoid of limbs, though some species, such as pythons and boas, have vestigial pelvic limbs. These vestigial limbs are referred to as spurs and are found on either side of the vent. The spurs are used in courtship behavior and can be larger in males. Some species of lizards also have rudimentary limbs that move in a manner similar to those of snakes. These legless species of lizards all retain vestigial thoracic and pelvic limb structures.
The pectoral girdle of salamanders consists of the precoracoid, coracoid, and scapula. All of these structures are cartilaginous, and they are ossified only at the point where they articulate with the humerus. The pectoral girdle of anurans is more complex and completely ossified. It is made up of the scapula, clavicle, and coracoid. The skeletal structure of the pectoral limb is comparable to that of reptiles, with the exception of the fused radius and ulna, called the radioulna, which is found in anurans. Anurans and salamanders typically have four toes on the pectoral limbs.
FIGURE 22-33 Different methods of locomotion employed by snakes. A, Concertina—the back half anchors while the front half moves forward, and vice versa. B, Lateral undulation—when the body comes in contact with an object or rough surface, it thrusts itself forward. C, Sidewinding—this is used by desert species on loose sand and involves moving laterally by throwing the body in loops sideways. D, Rectilinear—this is used by large snakes, such as boas and pythons, which use the ventral skin and attached muscle to pull themselves forward in a straight line, similar to a caterpillar. (Redrawn from O'Malley B: Clinical anatomy and physiology of exotic species, New York, 2005, Saunders.)
The pelvic girdle of amphibians consists of the ilium, ischium, and pubis. The pelvic limb of anurans is elongated in comparison to the forelimb and is well adapted to swimming and jumping. The skeletal structure of the amphibian hindlimb is comparable to that of other vertebrates, the exception being the elongated metatarsal bones in anurans, which are known as the astragalus and calcaneum. Five toes are typically present on the hindlimbs of anurans and salamanders.
MUSCLES
The muscular structure of four-limbed amphibians and reptiles varies according to species, but is somewhat analogous to the muscular structure in mammals.
Snakes have well-developed epaxial muscles, as well as segmental muscles that attach the ventral scutes to the ends of the ribs. This muscular arrangement allows a number of effective methods of locomotion (Figure 22-33).
Amphibian and reptilian muscles are incapable of sustained aerobic metabolism and switch quickly to anaerobic metabolism with prolonged physical exertion. Anaerobic metabolism is less efficient than aerobic metabolism and results in rapid buildup of lactate. Lactate decreases the pH of the blood and tissues and is metabolized slowly by reptiles. For this reason, reptiles are unable to sustain physical activity and are capable of only short bursts of motion.
TEST YOURSELF 22-10
1. What adaptations are found in the snake skull that allow the ingestion of large prey?
2. What is tail autonomy?
3. What is unique about the attachment between the skull and cervical spine of a reptile?
4. Why are reptiles unable to sustain physical activity for long periods of time?
SUMMARY
Amphibians and reptiles are remarkable creatures. Collectively known as herptiles, they differ from mammals in extraordinary ways. For example, unlike mammals, herptiles are ectothermic, and regulate their body temperature by adjusting their posture and by moving to areas that are cooler or hotter as needed. Herptiles consist of four orders of reptiles and three orders of amphibians.
Like that of mammals, the skin of reptiles includes an epidermis, dermis, and subcutaneous space. The epidermis of most reptiles has distinct scales and can form unique struc- tuucrhes s as crests, tubercles, and spines. The dermis is composed of dense connective tissue, which may contain bony plates for protection. Reptiles shed their skin as they grow, in raopcess called echysis. The epidermis of amphibians, in contrast, is very thin and permeable, which allows them to labsorb al the water they need percutaneously. They do not, therefore, need to drink. Like reptiles, amphibians shed the fyoeurter la o the epidermis and often consume the exuvia.
dTihoevacsacrular system of herptiles is quite different from that of mammals. Remarkably, most reptiles and all amphibians have a three-chambered heart, not four as om, ammals d consisting of two atria and one large ventricle. Heart rate depends on species, size, temperature, activity level, and metabolic function. Therefore, it can be challeng- ieontegrmt inde whether or not a herptilian patient has an
abnormal heart rate. The lymphatic system in herptiles is evlevlreylowped d with large lymph vessels. Herptiles have
ynmo plh nodes but instead possess sections of special
leysmseplsh v that include smooth muscles in the vessel wall. These specialized lymph vessels (known as lymph hearts) actively pump lymph through the vasculature. Unlike that of empatimliamnals, r blood contains oval, nucleated red blood cells and phagocytic heterophils. The enzyme-rich neutrophil of mammals liquefies necrotic debris and produces fluid, white pus. In contrast, heterophils in reptiles lack these enzymes and therefore produce solid, cheeselike pus.
vRe ptiles ha a remarkably large pulmonary volume, effi- ceireonbticana metabolism, and cardiac shunting capabilities
that enable them to survive for long periods of time without breathing. They can function with very low oxygen levels and may possess one of three different types of lung (unicameral, paucicameral, or multicameral) depending on species. In contrast, the lungs of amphibians are typically simple saclike structures, and some species, such as salamanders, lack lungs entirely and exchange gases through their skin instead. The pulmonary tissue in herptiles has a honeycomb appearance and terminates in gas-exchange structures called faveoli (rather than alveoli, which are found in mammals).
Interestingly, all adult amphibians are carnivorous, ewphteilreesas r may be herbivorous, omnivorous, or carnivorous depending on species. Snakes and crocodilians are strict carnivores, whereas various species of lizards and che- lyonians ma utilize any one of the three feeding strategies. Snakes and lizards use their flickering tongues to detect their typurreiynbg cap ambient particles in the air. The tongue is subsequently inserted into the vomeronasal organ located in the roof of the mouth. There, the molecules stimulate sensory innapbulitn, ge the animal to track its prey or find water, for
ehxeample. T rest of the gastrointestinal tract in reptiles is
aqruiiatbelev among species, but most herptiles have the usual parts: teeth, salivary glands, esophagus, stomach, liver, gallbladder, intestinal tract, pancreas, spleen, and cloaca. The rsiuzcetuanred st of these organs are highly variable among species. Similarly, great variability exists in the structure of the urogenital system and the location, size, shape, and fnumber o the kidneys, ureters, urodeum, bladder, and adrenal glands. With respect to the reproductive system, all empatlielesr have internal testes and most possess a sexual poortion t the kidney tubules that develops in response to high levels of sex hormones. Male snakes and lizards possess rpuacirteudresst called hemipenes, which are everted
through the vent for mating. The hemipenes form a trough thhicrohugh w semen is carried. However, the phallus has no role in urination. Once fertilized, females can either lay eggs (oviparous) or deliver live offspring (viviparous). Gender in reeteprtmileisniesdd by either the genotype or the tempera-
thuicreh at w the eggs are incubated. Sex chromosomes are rneostenpt in some species such as crocodilians, therefore seetexrmisinded during incubation. In crocodilians and
lhizearrds, hig temperatures produce males and lower tem- rpoedrautcueres p females. In other species, the reverse may rbueet.
vRe ptiles ha a cerebral cortex separated into two hemi- sopwheevrers,. H the reptilian brain contains no sulci or gyri
ounrfaitcse s and is therefore called lissencephalic. Reptiles dososensost p a cauda equine, so the spinal cord extends to iftphe t o the tail. They are also the lowest vertebrates to have in cranial nerves, whereas amphibians have 10. Both reptiles ahnibdiaanms p depend more on segmental reflexes and
locomotion centers for the control of movement than they do eorne bcral stimulation. The amphibian brain is well adapted tuopsport basic functions such as sight, olfaction, and movement, but they have very limited integrative capabilities.
The axial and appendicular skeletons of reptiles and amphibians differ from those of mammals in many ways. xWceitpht iothne e of chelonians, the reptilian spine is
extremely flexible and is divided into the presacral, sacral, and caudal vertebral regions, rather than cervical, thoracic, sacral, and caudal as seen in mammals. A single occipital coormndsyle f the articulation between the skull and spine. uTmhebner of vertebrae and the presence of ribs vary from sopecies t species. The skeletal structures of reptilian limbs aorme pcarable to those of mammals, with a few exceptions.
Unlike mammals, the pectoral girdle of a reptile consists of a scapula and coracoid bone. In most reptiles the hindlimbs anrgeerlo than the forelimbs. Snakes are devoid of limbs entirely, though a few species have vestigial hindlimbs.
Lastly, the muscular systems of amphibians and reptiles laere incapab of sustaining aerobic metabolism for a long time. Genseqiieritly, they are capable of only quick bursts of motion. So, if you can avoid getting caught after the startling tohrrwuasrtdf and powerful chomp of an alligator, you stand a ^>d chance of getting away... a helpful titbit to itotauhke w y on your next trip to the bayou. Garnivorous ehnerdptiles t to lie in wait for their prey, but when they move. they move VERY fast!
FURTHER READING
DueUman W, Trueb L: Biology of amphibians, Baltimore, 1994, Johns Hopkins University Press.
Mader D: Reptile medicine and surgery, ed 2, St Louis, 2006, Saunders.
McArthur S, Wilkinson R, Meyer J: Medicine and surgery of tortoises and turtles, Ames, IA, 2004, Blackwell Publishing.
O'Malley B: Clinical anatomy and physiology of exotic species, New York, 2005, WB Saunders.
Pough FH, Andrews RM, Cadle JE, et al: Herpetology, ed 3, Upper Saddle River, NJ, 2004, Prentice-Hall.
Wright K, Wliilaker B: Amphibian medicine and captive husbandry, Malabar, FL, 2001, Krieger Publishing.
This page intentionally left blank