Avian Anatomy and Physiology
Lori R. Arent
OUTLINE
INTRODUCTION, 502
TOPOGRAPHY, 503
INTEGUMENT, 503
Skin, 503
Glands, 503
Beaks, 503
Claws, 504
Feathers, 504
MUSCULOSKELETAL SYSTEM, 510
Skeleton, 510
Muscles, 513
SENSE ORGANS, 519
Vision, 519
Hearing and Equilibrium, 521
Taste, 522
Touch, 522
Smell, 523
ENDOCRINE SYSTEM, 523
DIGESTIVE SYSTEM, 523
Anatomy, 524
CIRCULATORY SYSTEM, 527
Anatomy, 527
Blood Flow, 529
Electrocardiogram, 530
Blood, 530
RESPIRATORY SYSTEM, 531
Anatomy, 532
Airflow, 534
Respiratory Rate, 535
Thermoregulation, 535
UROGENITAL SYSTEM, 536
Urinary System, 536
Reproductive System, 537
SUMMARY, 539
LEARNING OBJECTIVES
When you have completed this chapter you will be able to:
1.
Describe the anatomic structures of avian skin, beaks, and claws.2. Describe the structure and functions of feathers.
3. List the types of feathers and the location of each type.
4. List the unique features of the avian skeleton.
5. List the unique features of avian musculature.
6. Describe the shape and general characteristics of avian eyes and explain how those characteristics affect visual acuity.
7. Describe the unique anatomic features that affect the sense of hearing in nocturnal owls.
8. List the unique anatomic features that affect the sense of taste in birds.
9. List the components of the avian digestive system and describe the functions of each.
10. List the unique features of the avian circulatory and respiratory systems.
11. Describe the composition of avian urine.
12. List and describe the unique features of the avian male and female reproductive systems.
13. List and describe the four classifications of newly hatched chicks.
VOCABULARY FUNDAMENTALS
Airsac ear-sahck
Alula ahl-yuh-luh
Appendicular skeleton ahp-ehn-dihck-u-lar skehl-ih-tuhn
Apteria ahp-teer-e-uh
Auricular ohr-rihck-yuh-lar
Axial skeleton ahck-se-ahl skehl-ih-tuhn
Barb bahrb
Barbule bahr-bul
Blood feather bluhd fehth-ar
Brood patch brood pahtch
Bursa of Fabricius buhr-suh of fah-brihsh-uhs
Choanae ko-ah-ne
Clutch kluhtch
Coecyg∣eal vertebrae kohck-sihj-e-ahl vart-eh-bra
Columella kohl-yuh-mehl-ah
Cone kδn
Coprodeum kδp-rδ-de-uhm
Crop krohp
Determinate layer dih-tar-mih-niht la-ar
aFra ult b -ahlt bahr
Feather e fehth-ar fohl-ih-kuhl
Fibrous tunic fi-bruhs too-nihck
Fovea fo-ve-uh
Gizzard gihz-ard
Vilenoid cavity gle-noyd kahv-ih-te
Gular fluttering gyoo-luhr fluht-ar-ihng
Hamuli hahm-yuh-lι
Heterophil heht-ar-δ-fihl
Hock hohck
Indeterminate layer ihn-dih-tar-mih-niht la-ar
Keel kel
Keratin kear-ah-tehn
Mesobronchi me-sδ-brohn-kι
Molting mohlt-ihng
Mute mut
Nape nap
Neural tonic nar-ahl too-nihck
Nictitating membrane nihck-tih-ta-tihng mehm-bran
Operculum δ-pehr-kyoo-luhm
Parabronchi pear-ah-brohn-kι
Patagium puh-ta-je-uhm
Pellet pehl-iht
Pecten pehck-tehn
Pectoral crest pehck-tar-ahl krehst
Pectoralis pehck-tah-rahl-ihs
Periderm pear-ih-darm
Precocial pre-ko-shuhl
Proctodeum prohck-tuh-de-uhm
Pterylae tehr-uh-le
Pygostyle pι-guh-stιl
Remiges rehm-ih-jez
Renal portal system re-nuhl pohr-tehl sihs-tehm
Retrices reht-ruh-sez
Rod rohd
Sclerotic ring skleh-rah-tihck rihng
Semialtricial seh-me-ahl-trihsh-uhl
Semiprecocial seh-me-pre-ko-shuhl
Shell ^ιd shehl glahnd
Supracoracoideus soo-prah-kohr-uh-koy-de-uhs
Synsacrum sihn-sa-kruhm
Syrinx sihr-ihncks
Talon tahl-uhn
Trochanter trδ-kahn-tar
Tympanic membrane tihm-pahn-ihck mehm-bran
Uneinate process uhn-suh-nat proh-sehs Urodeum yar-δ-de-uhm
Uropygial ^ιd yar- δ-pihj-e -ahi glahnd
Uterus u-tar-uhs
Uveal tunic u-ve-ahl too-nihck
Vent vehnt
Zygodactyl zι-gδ-dahck-tihl
INTRODUCTION
A feathered marvel.
There is no better way to describe the avian creature. Over 10,000 unique bird species grace our planet (Table 21-1), modeling colors from the brilliant hues of tropical species to the earthy tones of many birds of prey. They are masterpieces of beauty and function. Over the course of time, a body containing specializedstructures and organ systems evolved to dominate a realm not shared by many other creatures: the sky. From the outer protective layers to the inner workings of the reproductive system, a bird is designed to fly. Let us take a look at this unique design and explore the wonders of the avian body.
From Gill FB: Ornithology, ed 2, New York, 1995, Macmillan.
TOPOGRAPHY
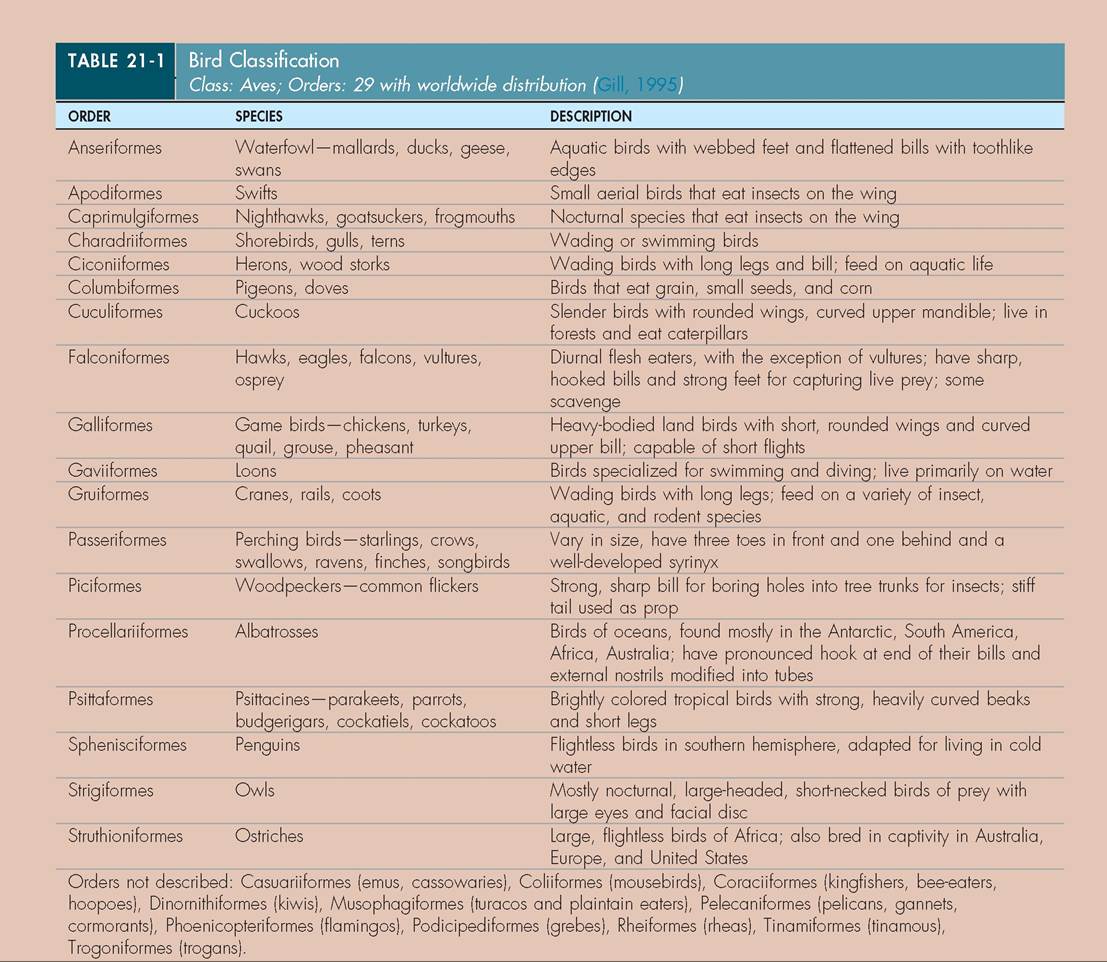
Our study of birds begins with the identification of external features (Figure 21-1). Many of these are used for identification of wild birds in the field and, in a clinical setting, for indication of illness or trauma. Familiarizing yourself with the locations of these structures will aid in your understanding of their function.
INTEGUMENT
A bird's body is covered by skin and its derivatives: the beak, claws, and feathers. These structures cover and protect the internal organs and block the entrance of disease-causing organisms.
SKIN
The skin of birds consists of two layers: an outer layer called the epidermis and an inner layer called the dermis (Figure 21-2). The epidermis is relatively thin and consists of flattened epithelial cells that produce keratin, a tough fibrous
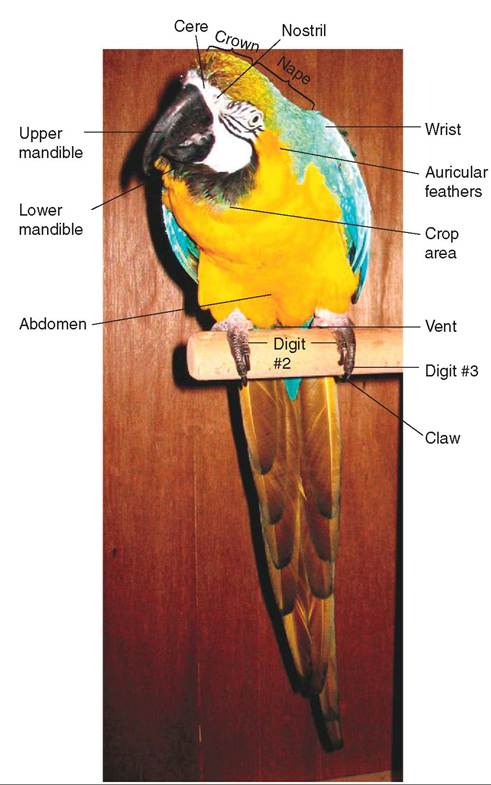
FIGURE 21-1 External features of a Blue and Gold Macaw (Propyr- rhura maracana). (Photo by Gail Buhl.)
protein necessary for the production of scales, feathers, and the outer sheaths of beaks and claws. The inner layer of skin (dermis) is thicker and consists of a tough, fibrous connective tissue. It stores fat for nutrition and insulation and supplies a pathway for nerves, blood vessels, and muscles to reach the epidermis.
Smooth muscles in the dermis innervate feather follicles to help in the regulation of heat. During hot weather, depressor muscles press the feathers against the body to promote heat loss. When a bird gets cold or does not feel well, it looks “fluffed” because erector muscles in the dermis elevate the body feathers to trap warm air near the body.GLANDS
Unlike mammals, birds do not possess sweat glands. Feathers cover such a large portion of a bird's body that sweat glands would not be effective. The one major skin gland that most birds do possess is called the uropygial gland, or preen gland. It is located on the dorsal surface at the upper base of the tail (Figure 21-3). The act of preening stimulates this gland to secrete an oily, fatty substance. A bird uses its beak to spread this oil throughout its feathers to clean and waterproof them. The gland varies in size and structure and is relatively large in aquatic species, such as waterfowl and ospreys. The gland is completely lacking in some parrots, ostriches, and in a few other species.
BEAKS
One derivative of a bird's skin is its beak, or bill. It consists of an upper and lower mandible and is covered with a tough, horny keratin layer that grows continuously. Beaks vary in their hardness and flexibility, depending on their function. Some birds use their beaks to crack seeds and nuts (parrots), tear food into bite-sized pieces (hawks), capture food (herons and woodpeckers), preen their feathers or those of a mate,
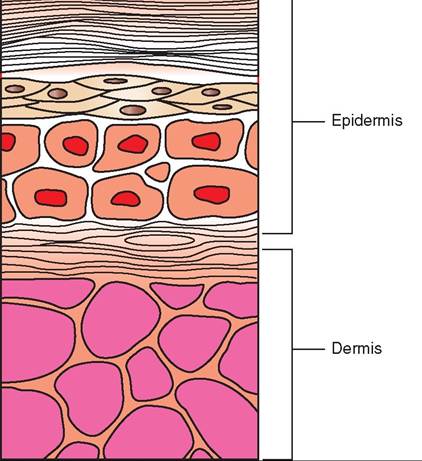
FIGURE 21-2 Layers of cells in avian skin.
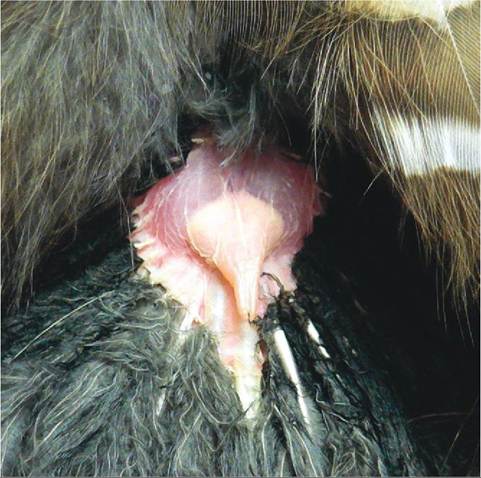
FIGURE 21-3 Uropygial gland of a Barred Owl (Strix varia).
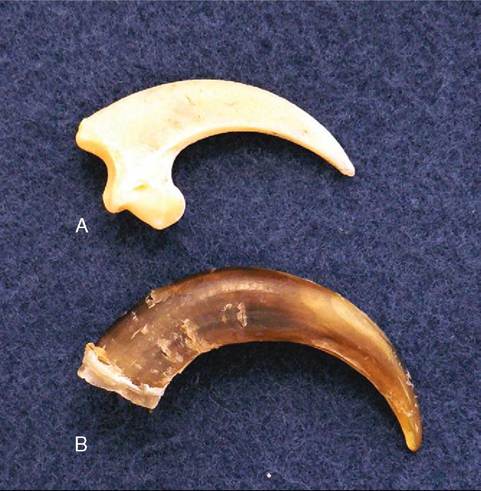
FIGURE 21-4 Talon of an American Bald Eagle (Haliaeetus leuco- cephalus). A, Bone; B, sheath.
pick up and hold things such as food and nesting material as they fly and climb, and sometimes to protect themselves.
CLAWS
Claws possess a horny sheath derived from specialized scales at the end of each toe. Like beaks, they grow continuously. Species differ in the type of claws they possess, based on their perching habits and method of procuring food. For example, ground feeders, such as chickens and pheasants, have short, sharp claws that are used to scratch the ground for food; birds of prey have claws called talons that are long, sharp, and rounded to catch and kill their prey (Figure 21-4); vultures, which are scavengers, have short, blunt claws; and climbing birds, such as woodpeckers and nuthatches, have strongly curved claws for gripping.
TEST YOURSELF 21-1
1. What structures are derivatives of a bird's skin and what are they made of?
2. Define the function of the uropygial gland. Do all birds possess this gland?
3. Describe the basic anatomy of a bird's beak and claws. When trimming these structures, what should you be careful to avoid?
FEATHERS
Birds are unique in the animal kingdom in that they possess feathers. Feathers are outgrowths of skin that are made of protein; once completely developed, they are nonliving
CLINICAL APPLICATION
Coping Beaks and Nails
In the wild, the shape and length of a bird’s beak and claws are maintained by daily activities that provide natural wear. For example, after completing a meal, birds often feak, or rub, their beaks on a rough surface to clean them and maintain their shape. In captivity, birds are provided with limited wearing surfaces, and some birds, such as psittacines and birds of prey, require frequent coping, or trimming and reshaping, of their beaks and nails. Without this maintenance, beaks can become so long that a bird has a difficult time eating, and they can develop large cracks and chips that can cause permanent damage to the beak’s growth plate, located near the cere. Claws can become so long and sharp that they cause uneven weight bearing on the foot pads or puncture the bottom of a foot.
This can result in pad abrasions, blisters, swelling, abscess formation, and, in severe cases, degeneration of the bones in the feet (osteomyelitis). The term often used to describe these conditions of the avian foot is bumblefoot.
Coping beaks and claws must be done with care, because both of these structures have a blood and nerve supply that can be hit if they are trimmed too deeply. Beaks can be coped using a fingernail file or a rotary tool. Claws can be trimmed using a cat or dog nail trimmer, depending on the size of the bird. If bleeding occurs, hemostasis can be achieved by applying topical cauterizing agents, such as silver nitrate or “Quick-stop.” The nails of parrots are often trimmed with a rotary tool, which cauterizes a blood vessel if it is hit.
∕j CLINICAL APPLICATION—cont'd

Abnormal beaks. A, Severely overgrown beak of a Budgerigar (Melopsittacus undulates). B, Cracked beak of a Peregrine Falcon (Falco peregrinus). (A from Samour J: Avian medicine, Philadelphia, 2000, Mosby.)
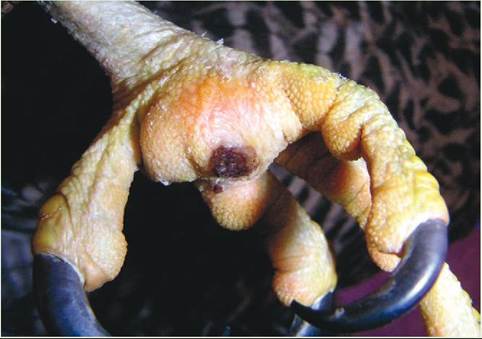
Severe bumblefoot on the metatarsal pad of a Gyrfalcon (Falco rusticolus).
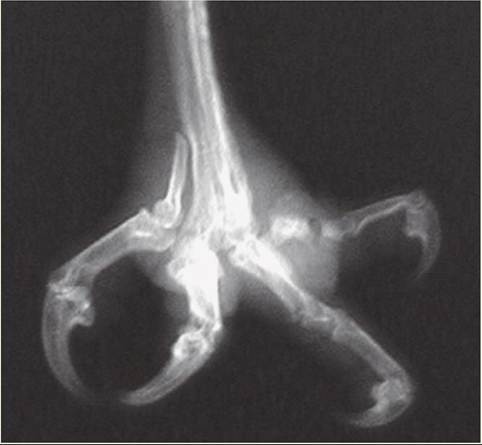
Osteomyelitis of digit number four.
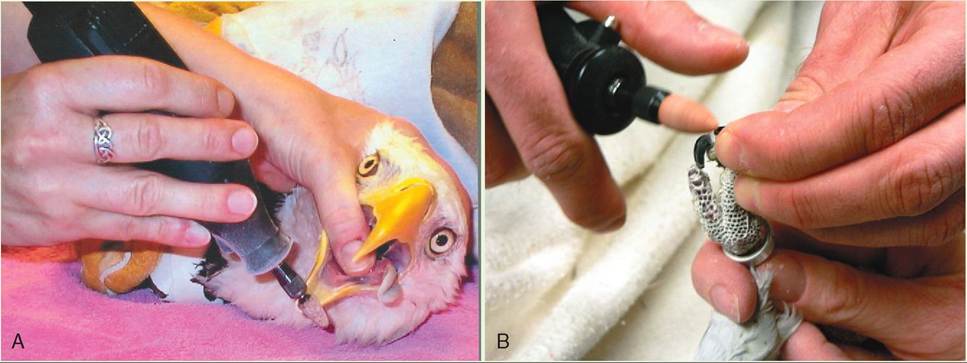
Coping with a rotary tool. A, Beak of an American Bald Eagle. B, Claws of a Cockatoo.
structures that have sensation only at the base, where they originate from a follicle under the skin.
FUNCTIONS
Feathers serve several important functions. First, they are necessary for flight. A bird without a proper complement of flight feathers simply cannot fly. Second, feathers protect the thin skin from trauma, rain, and excessive radiation from sunlight.
They also assist in thermoregulation and camouflage and are used in many communication behaviors, such as courtship, defense, and recognition.Structure
Six types of feather cover a bird's body. The most visible feathers, that give shape to a bird, are called contour feathers. Contour feathers have the most compact microstructure and consist of several components (Figure 21-5, A).
INFERIOR UMBILICUS. This structure is a tiny opening at the base of the feather, where it inserts into the skin. When a new feather is developing, it receives nourishment from blood vessels that pass through this opening.
SUPERIOR UMBILICUS. This structure is a tiny opening on the feather shaft, where the webbed part of the feather begins. In some birds, including several species of parrots, hawks, herons, and grouse, it gives rise to an afterfeather, which is an accessory feather thought to provide additional insulation to retain body heat.
CALAMUS. Also called the quill, the calamus is the round, hollow, semitransparent portion of a feather that extends from the inferior umbilicus to the superior umbilicus.
RACHIS. The rachis is the main feather shaft.
VANE. The vane is the flattened part of a feather that appears weblike, on each side of the rachis. It consists of numerous slender, closely spaced barbs. The barbs give rise to barbules, which have rolled edges and tiny hooklets known as hamuli. These hooklets interlock each barb with an adjacent one, forming a tightly linked, flexible web (see Figure 21-5, B). The degree of tightness varies with the species. For example, the contour feathers of owls have fewer barbules than do those of hawks. The result is a looser feather weave that feels softer and allows air to pass through, creating silent flight.
TYPES OF FEATHER
CONTOUR FEATHERS. Contour feathers typically cover a bird's body and constitute the flight feathers of the wings and tail (Figure 21-6, A). The flight feathers in the wing are commonly called remiges, and the tail feathers are called retrices. Small contour feathers, called auriculars, are found around the external ear openings and improve a bird's hearing ability. They are especially numerous in some species of parrots and hawks, and in owls. Contour feathers are moved by muscles attached to the walls of the follicles.
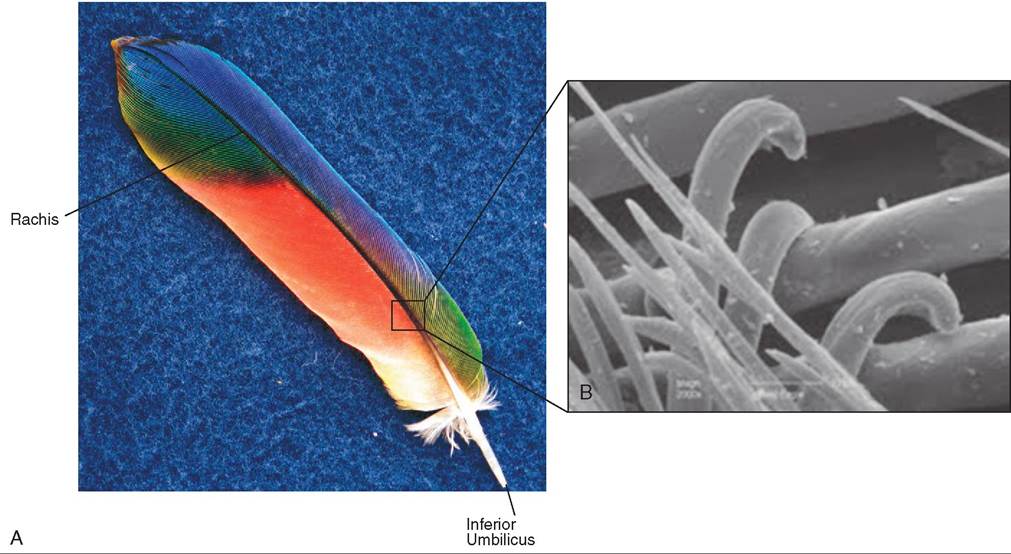
FIGURE 21-5 Contour feather. A, General structure. B, Electron microscopic view of the vane of a Bald Eagle flight feather.
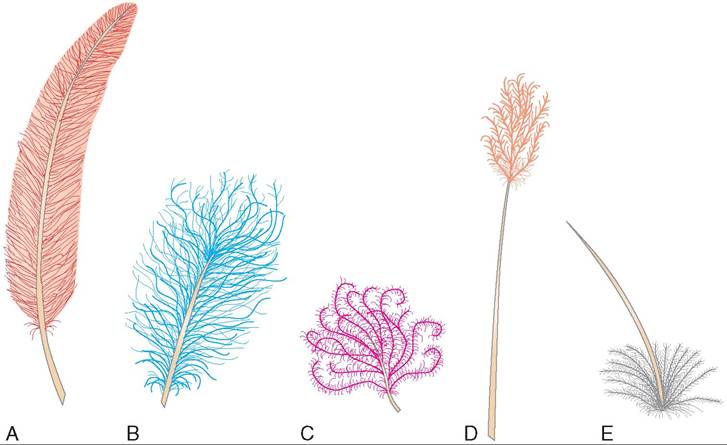
FIGURE 21-6 Types of feather. A, Contour. B, Semiplume. C, Down. D, Filoplume. E, Bristle.
SEMIPLUME FEATHERS. Semiplume feathers possess a main rachis with barbs that lack barbules and hooklets (see Figure 21-6, B). They are commonly found under contour feathers, especially on the sides of the abdomen and along the neck and back. Like down feathers, semiplumes provide insulation. They also provide flexibility for the movement of the contour feathers and help with buoyancy in water birds.
DOWN FEATHERS. Down feathers are soft, fluffy feathers that lack a true shaft, barbules, and hooklets on their barbs (see Figure 21-6, C). They are located next to the skin under contour feathers, and they function primarily as insulation.
FILOPLUME FEATHERS. Filoplume feathers have a bare shaft that lacks barbs on the majority of its length, except at the tip (see Figure 21-6, D). They are located on the nape and upper back near contour feathers, and their follicles contain sensitive nerve endings that may play a sensory role in controlling feather movement. Slight movements of the contour feathers are transmitted to pressure and vibration receptors in the skin via the filoplume feathers.
BRISTLES. Bristles are modified contour feathers with a stiff rachis and few barbs at the base (see Figure 21-6, E). They are thought to serve a bird's sense of touch. Depending on the species, they may be found in various locations. Crows, ravens, and woodpeckers have bristles around their nostrils; owls have them around the mouth and sometimes around the toes, as do grouse; other birds have bristles around their eyes (Figure 21-7).
POWDER DOWN FEATHERS. Powder down feathers are unusual feathers that never stop growing. They grow continuously at the base and disintegrate at their tip, creating

FIGURE 21-7 Bristles on the face of a Great Horned Owl (Bubo virginianus).
a waxy powder that is spread throughout the rest of the plumage to clean it and provide waterproofing. Powder down feathers are most highly developed in herons and bitterns, especially on the breast, belly, and back; they can be found more diffusely scattered in parrots and hawks. Birds without a uropygial gland have abundant powder down feathers.
LOCATION
Contrary to their appearance, feathers do not originate from the entire body. They are located in specific tracts, called pterylae, which are separated by bare areas of skin called apteria (Figure 21-8). However, the feathers in these tracts overlap one another to create the fully feathered look.
FEATHER DAMAGE
Feathers are durable structures but still can be damaged. External parasites, such as some species of feather mite, can
FIGURE 21-9 Feather damage. A, Feather mite (not drawn to scale). B, Mite damage on the tail feather of a Great Gray Owl (Strix nebulosa). C, Fault bars on the tail feather of a Red-Tailed Hawk.
chew and consume parts of the feather vanes, creating weak points (Figure 21-9, A and B). Damage also can occur from daily wear and tear. Often, the lighter tips of a bird's flight and tail feathers are worn off by the roughness of daily activities, giving the feathers a more iridescent appearance. In some species, such as Mallard ducks (Anas platyrhynchos) and European Starlings (Sturnus vulgaris), this is most noticeable in the spring, when males of many species have lost their light feather tips and look more colorful before the breeding season.
Another cause of feather damage occurs during a feather's growth phase. If a feather is stressed during its growth, even for a few hours, there is an interruption in its blood flow. What develops is called a fault bar, or stress bar, which is characterized by a weakened area on the feather vane, where the barbs lack barbules (Figure 21-9, C). When the stressor is removed, the blood supply is returned and normal development continues. The most common stressor is a poor diet. An insufficient food supply or a food supply deficient in essential nutrients often creates fault bars on developing feathers. This can have a severe effect on the plumage of nestling birds, because all of their flight feathers grow in at the same time. Any stressor that temporarily depletes the blood supply to these feathers creates fault bars on all of them.
MOLTING
The process of feather replacement is called molting, and it occurs once to several times a year depending on the species. Molting occurs in a species-specific pattern that allows a bird to continue normal activities, such as procuring food, escaping from predators, reproducing, and finding safe roosting sites. In most species, feather replacement is symmetric; one or two pairs of flight feathers are molted at a time so that a bird still can fly adequately. One major exception is found in many species of waterfowl, which molt their flight feathers all at once after the breeding season. They are flightless during this time but can forage by grazing on land or in the water.
Between 4% and 12% of a bird's body weight is made up of feathers. Replacing them is a very energy-demanding process that requires a well-balanced diet. In many North
CLINICAL APPLICATION
Feather-Picking Disorder
One condition seen in many species of psittacine and some human-imprinted raptors is feather picking. Birds with this disorder preen excessively, removing most to all of their body feathers, especially on their chest and legs. Also, in severe cases, the skin surrounding the feathers is self-mutilated.
Causes of this disorder are either pathologic or psychologic. In the first category, toxins, bacteria, viruses, fungi, parasites, and malnutrition all can lead to feather picking. To determine the cause, a thorough physical examination must be conducted. Radiographs, blood samples for complete blood count and serum chemistry, cytology of feather pulp or of a local skin scraping, feather biopsy, and endoscopy are all diagnostic tools that can be used. If the problem does not appear to be physiologic, then attention must be turned to psychologic causes. These may include changes in the environment, diet, human exposure, boredom, sexual frustration, anxiety, or exposure to new pets. Many species of parrot, especially African Greys (Psittacus erithacus), are very sensitive to these types of conditions.
Treatments for the disorder vary with the cause. Bacterial, viral, parasitic, and fungal infections can be treated with established protocols, and diet can be improved and varied. However, treatment for psychologic causes is more difficult, especially if the disorder cannot be attributed to a specific event or situation. Most often, changing components of the care and management of the bird is required, with no guarantee of inhibiting the feather-picking behavior.
American species, the major annual molt is timed so that it occurs between the end of the breeding season and the beginning of migration. Food is usually abundant during this time, and a bird's energy can be directed to its growing feathers.
Feathers develop from papillae in the dermis layer of the skin (Figure 21-10, A). These papillae are located in the feather tracts and contain germ cells with the genetic information that dictates the type, size, and color of feathers. These cells are “activated” by physiologic and environmental factors. Increasing day length stimulates the pituitary and thyroid glands to produce hormones that stimulate molting, and sex hormones also may play a concurrent role.
Molting begins when a newly developing feather pushes an old one out (Figure 21-10, B). It then emerges from the skin and is covered by an epidermal covering called the periderm. As a bird begins to preen a growing feather, it gently removes the periderm, which can be seen as small, white flakes in the plumage. Blood vessels from the dermis reach into the new feather (through the superior umbiculus) to provide nourishment. When a feather is fully grown, the blood dries up, and the rachis is pinched closed under the skin.
During feather development, a growing feather is called a blood feather. Blood can be seen in the proximal part of the feather shaft during the entire growth phase
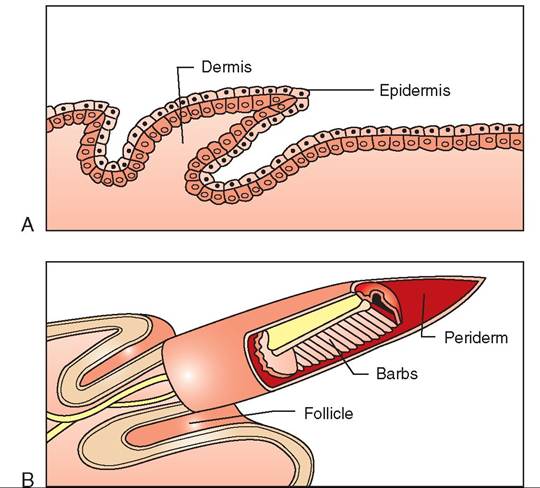
FIGURE 21-10 Stages of feather growth. A, Feather papilla. B, Newly developing feather.

FIGURE 21-11 Blood feathers on the wing of a White-Breasted Caique (Pionites leucogaster).
(Figure 21-11). Injury to a blood feather not only results in bleeding but can prevent a feather from developing normally until molted again.
TEST YOURSELF 21-2
1. List three major functions of feathers.
2. What types of feather are the flight and tail feathers? Describe their microstructure.
3. Define a fault bar. What causes it?
4. What is a blood feather?
5. How do the wing and tail feathers differ between predatory and prey bird species?
∕j CLINICAL APPLICATION
Clipping Wing Feathers
Many pet bird owners desire to have their bird’s flight feathers clipped. The goal of clipping flight feathers is not to make a bird completely flightless, it is to prevent a bird from gaining lift while still allowing it to glide safely to the ground. This prevents a bird from injuring itself when out of its cage and prevents accidental escape. Numerous pet birds are lost every year because they fly out of open doors and windows.
Several patterns are used to clip a bird’s wing feathers. The one chosen depends on the owner’s wishes and the personal preference of the clinician. One technique that many practitioners prefer is to trim the outermost five to seven flight feathers under the overlying coverts, giving a smooth appearance to the wing. Another technique leaves the outer two to four primary flight feathers intact and then trims the next five to seven flight feathers underneath the overlying covert feathers. One problem encountered with this pattern is that the long outer primaries are vulnerable to breakage. Whichever technique is used, the clipping should be done symmetrically on both wings.
Another consideration in feather clipping involves the presence of blood feathers. If blood feathers are present, it is advisable to wait until those feathers are completely developed before any are trimmed. If blood feathers are cut or damaged, a significant amount of bleeding often occurs. If waiting is not an option, completely developed feathers can be trimmed such that one feather remains intact on each side of every blood feather. Intact feathers on each side are needed to protect a growing feather from injury.
∕j CLINICAL APPLICATION
Treating a Damaged Blood Feather
If a developing feather is cut, nicked, or otherwise damaged, it will often bleed profusely. To treat minor nicks or cuts, pressure can be applied to the affected area and, when dry, “Quickstop” or a tissue glue may be applied to ensure coagulation. If more severe injury to a blood feather occurs, the most appropriate way to treat it depends on the type of bird. Many pet bird species are considered prey species and their major flight feathers are loosely seated in the follicles. Therefore, it is acceptable gently to pull out a damaged feather and pack the skin opening with “Quick-stop.”
However, because developing feathers have a good nerve supply, anesthetizing the patient to prevent pain or discomfort may be appropriate before pulling a large feather. If bleeding is persistent after a feather is pulled, the skin opening can be sutured closed for a few days to promote hemostasis, but it should be reopened to allow normal healing and new feather growth to begin.
In predatory bird species, such as birds of prey, the feathers are seated very strongly in the follicles, and pulling out a flight or tail feather can result in permanent follicle damage that results in abnormal feather growth or prevention of growth altogether. In these species, the proper approach to treating a damaged blood feather is to stop the bleeding and allow the damaged feather to be removed by the bird or to be molted during its normal cycle.
MUSCULOSKELETAL SYSTEM
Feathers are not the only components of a bird that make it unique. A mammal or reptile with feathers still could not fly. Specially designed body systems that complement the feathers are necessary to create a structure that can support aerial locomotion. The first two systems we are going to look at are the skeleton and musculature.
SKELETON
Although muscles do the work to create specific movements, they must be supported by a sturdy framework (Figure 21-12). In birds, this framework is highly specialized, because it must support two very different modes of locomotion: walking and flying. As we will see, the special design of the avian skeleton includes many unique features that all contribute to the creation of a remarkably lightweight but sturdy structure. The lightweight nature of the skeleton was a key component in the evolution of flight and can be explained by several general modifications:
• Reduction in the number of bones
• Fusion of bones to form plates that provide strength and simplify movements
• Reduction in the density of bones, which are relatively thin but strengthened by a network of internal bony braces
• Loss of internal bone matrix (the bones of birds are generally hollow and filled with air spaces)
Additional features of the avian skeleton that contribute to its lightness can be seen throughout its structure. To study these, we can divide the skeletal components into two major sections: the axial skeleton and the appendicular skeleton. The bones that provide the general framework of the avian body include the skull, vertebral column, and sternum and are collectively called the axial skeleton. The wings, shoulder bones, legs, and pelvic bones support locomotion and are collectively called the appendicular skeleton.
AXIAL SKELETON
SKULL. The bird skull possesses several adaptations for lightness (Figure 21-13). The bones of the skull are thinner than in other animals, and instead of supporting heavy teeth, the jaws extend into a keratinized bill. The shape of the bill varies with the species, and it consists of a lower and upper component. The lower bill hinges on two small, movable bones called the quadrates. The upper bill has a somewhat
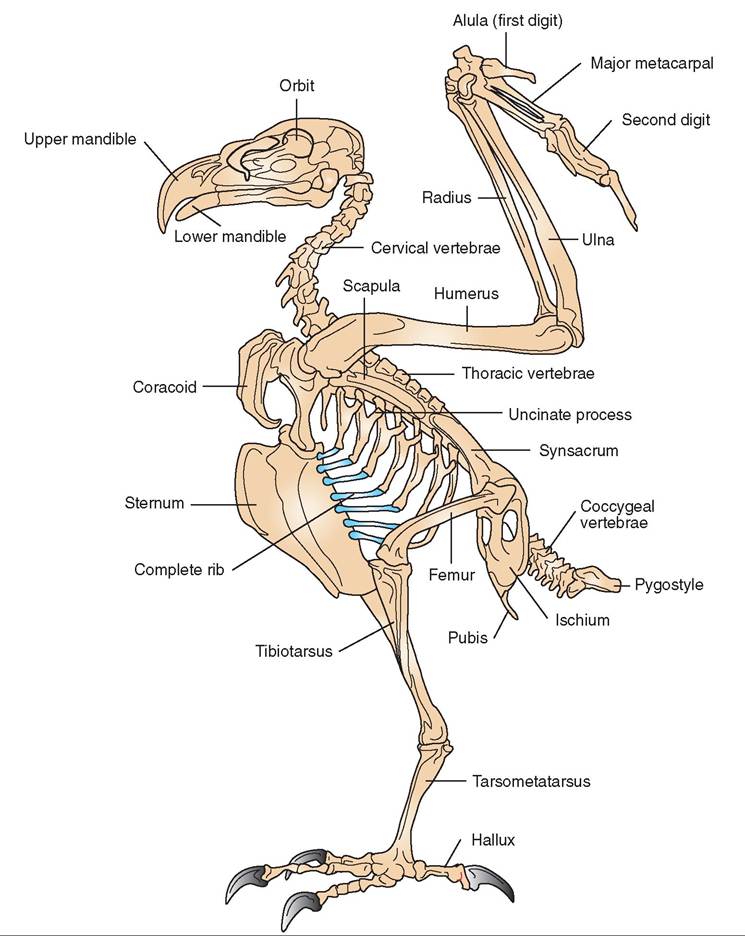
FIGURE 21-12 Skeleton of α hawk.
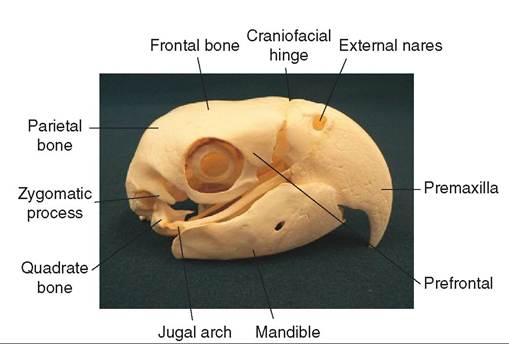
FIGURE 21-13 Skull ofαBlueond GoIdMacaw.
flexible attach ment to the skull and can move, although only slightly. Wιs, birds actually can move their upper and lower bills independently, which gives them greater control in manipulating food and increases their gape. The gape is the size of the bird’s mouth when open; the larger the gape, the larger the pieces of food that can be ingested.
Another prominent feature of the skull is the eye sockets. Good vision is important for an aerial creature, and thus a large portion of a bird's skull is devoted to supporting and protecting the eyes. The avian skull has large eye sockets that are bordered by a ring of protective bony plates called the sclerotic ring. Iie most species, a relatively small portion of lthe skul is devoted to the olfactory system and, as we will later see, the size of the ear canal varies with the species and ietsst ylilfe.
VERTEBRAL COLUMN. The vertebral column of birds is similar to that of other animals in that it consists of five general groups of vertebrae: cervical, thoracic, lumbar, sacral, and coccygeal. Birds have fewer vertebrae than other animals in the three central regions, but they have more vertebrae in the cervical and coccygeal areas, allowing greater mobility of their neck and tail, respectively.
CERVICAL VERTEBRAE. The first cervical vertebra, the atlas, contains a single condyle (ball and socket type of structure) for attachment to the skull. This allows a greater range of head movements when compared with mammals, which have two condyles attaching the skull to the vertebral column. In addition, birds have more cervical vertebrae than mammals, ranging from 11 in parakeets to 25 in swans, whereas all mammals have seven. In birds, these vertebrae have special connecting surfaces that allow movement, thus contributing to the flexibility of their necks.
THORACIC VERTEBRAE. The thoracic vertebrae are rigid and provide a strong support for the rib cage. In birds, the first few ribs are relatively short and incomplete. The other ribs are complete, attach to the underside of the sternum, and possess a projection called the uncinate process that overlaps the adjoining rear rib to strengthen the rib cage (see Figure 21-12). One exception to the rigidity of these vertebrae is found in penguins; these birds swim like fish and require a high degree of flexibility in their backs.
LUMBAR AND SACRAL VERTEBRAE. These two groups of vertebra are also rigid. Several of the distal lumbar vertebrae fuse with the sacral vertebrae and the first few coccygeal vertebrae to form a light, strong, bony plate called the syn- sacrum. This plate in turn fuses with the pelvis to provide a stiff framework for support of the legs. When a bird lands, the synsacrum acts as a shock absorber to protect the legs and backbone from injury.
COCCYGEAL VERTEBRAE. Birds have an average of 12 coccygeal vertebrae. The first few are mobile to allow movement of the tail feathers during flight. The rest are fused into a bony structure called the pygostyle that supports the tail feathers.
STERNUM. In most species of bird, the sternum is large and concave. It not only protects the chest from traumatic injuries but also acts as the place of origin of the flight muscles (see Figure 21-18 later in the chapter). In strong fliers, the sternum has a large bony ridge, or keel, to which the muscles attach (Figure 21-14). In flightless birds, such as the ostrich, the sternum lacks a keel entirely.
APPENDICULAR SKELETON
PECTORAL GIRDLE. The pectoral (shoulder) girdle consists of three pairs of bones: the coracoids, scapulas, and clavicles (see Figure 21-14). On each side, the coracoid and scapula are joined, forming a depression called the glenoid
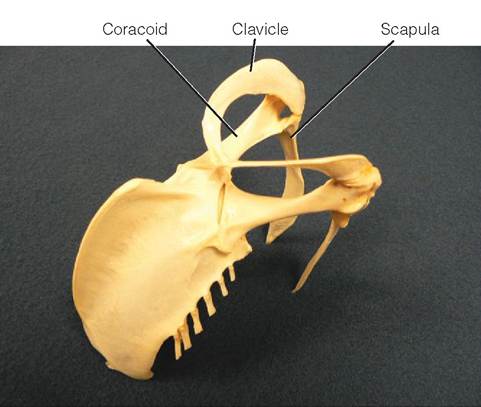
FIGURE 21-14 Anatomy of the pectoral girdle of a Rough-Legged Hawk (Buteo lagopus).
cavity, or triosseal canal. The wing attaches to the body by forming a joint in this cavity. During contraction of the powerful flight muscles, the strong, broad coracoids help to protect the sternum; the scapulas, positioned along the backbone, help protect the rib cage; and the clavicles, also known as the wishbone, are positioned outward and forward from the body and keep a bird's shoulders separated. Trauma to the pectoral girdle is not uncommon when birds collide with glass windows and doors (Box 21-1).
WINGS. The wings are connected to the body by forming a joint with the shoulder girdle. This joint is highly flexible, allowing rotation of the wing in several planes. The humerus extends from the shoulder to the elbow joint and possesses a pectoral crest for the attachment of wing muscles (Figure 21-15). The length of the humerus varies among species, being relatively short in birds that depend primarily on flapping flight and relatively long in birds that glide and soar.
The elbow joint is less flexible than the shoulder, and it only allows wing movement parallel to the wing. The radius and ulna extend from this joint to the wrist. In birds, the ulna has a larger diameter than the radius (it is the weightbearing bone) and acts as an attachment point for the secondary flight feathers. These two bones create the forearm of the wing and slide past each other slightly during flight.
Extending from the shoulder to the wrist is a web of skin called the patagium, or propatagium. This skin is lightly vascularized and possesses a ligament that runs along its cranial edge. It provides elasticity to the wing and assists in the aerodynamics of flight. If a bird damages its patagium, it may be grounded permanently.
The wrist joint consists of two bones and, like the elbow, allows movement only in the plane of the wing. The first finger, called the alula bone, originates from the wrist and carries the alula feathers, which are important for steering. The major and minor metacarpal bones extend from the wrist and join with the second and third fingers near the
BOX 21-1
Trauma and Skeletal Fracture of a Cockatiel Sample SOAP
"Polly," a 7-year-old female Cockatiel, was presented to the veterinary clinic in the afternoon of 3/13. According to the owner, "Polly" had been perched on her shoulder the previous morning, was spooked by the household cat, and flew hard into the kitchen window. Subsequently, she appeared dazed, would not eat, and stood on the bottom of her cage. Subjective: Adult female cockatiel with recent window strike.
On a pelleted diet and no history of egg laying. Lethargic with eyes closed, feathers fluffed, slight left wing droop at shoulder.
Objective: Weight = 93 g, T = 106.5° F, RR = 45 per minute, HR 300 bpm. Small amount of blood present in oral cavity. Crepitation of L shoulder girdle during extension of L wing. PE otherwise WNL.
Assessment: Probable L shoulder girdle trauma, anorexia, lethargy.
Plan:
1. Whole body radiographs (VD/R lateral): closed simple midshaft fracture of L coracoids.
2. CBC/Avian Chemistry Panel: WNL except for elevated CPK (2300 IU) and low normal calcium (8.0 mg/dl).
3. LRS SQ at 50 ml/kg.
4. Meloxicam PO at 0.5 mg/kg q24h.
5. Stabilize overnight and consider release tomorrow.
6. Provide small convalescent cage with soft substrate.
7. Client education: cage rest with reduced activity for a minimum of 3 weeks, maintain Polly in small convalescent cage with a floor perch and food and water dishes on floor, administer meloxicam daily for 10 days (can be placed in favorite food), offer extra calcium (e.g., cuttle bone, egg with shell), discuss feather trimming.
8. Dispense meloxicam as per order.
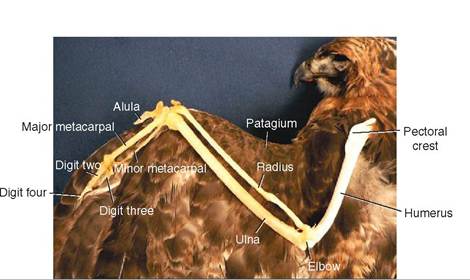
FIGURE 21-15 Wing bones identified in a Red-Tailed Hawk.
distal end of the wing. These two fingers, along with the metacarpal bones, support the primary flight feathers.
PELVIC GIRDLE. The pelvic girdle provides a rigid framework for support of the legs (Figure 21-16). Each side is made up of three bones that join where the leg attaches to the body: the ileum is relatively broad and is fused to the synsacrum; the ischium and pubis are thin and long and are fused to the anterior ileum and directed rearward, parallel to the backbone. The distal ends of these three bones are not fused, leaving the lower part of the pelvis open. This provides room for the abdomen and facilitates egg laying in hens.
LEGS. The hip joint in birds is well hidden by thigh muscles and is the place where the femur attaches to the pelvis. The femur is often referred to as the drumstick and is relatively short but wide. It ends at the knee joint (stifle), where it is directed a little forward so that the lower part of a bird's leg is placed under its center of gravity. Similar to the pectoral crest of the humerus, the femur possesses two crests called the greater and lesser trochanters, where leg muscles attach. Two bones are located in the middle section of the leg, called the tibiotarsus and fibula. The fibula is relatively small in diameter and acts as a splint. These two bones end at the ankle or hock joint, and the ankle itself consists of a single, elongated bone called the tarsometatarsus.
FEET. The bottom of a bird's foot is called the metatarsal pad. It is surrounded by two, three, or four toes, with the majority of species possessing four. Often one toe faces the rear and the other three face forward, which is called anisodactyl. However, in some species, such as parrots and woodpeckers, the second and third toes face forward, and the first and fourth are directed backward; this is known as zygodactyl. Owls, ospreys, and cuckoos also have this arrangement, but the fourth toe is opposable and can face either forward or backward.
The digits are referred to by a numbering system based on the number of joints they have. The rear toe is digit number one and has one joint, where it hinges to the metatarsal pad. Digit two is the innermost, medial digit and possesses two joints. The middle toe is digit three; it has three joints. The outer, lateral toe has four joints and is digit four.
MUSCLES
The average bird has 175 to 200 muscles, many of which have been placed ventrally, near the center of gravity. Reptiles, the avian predecessor, have muscles on their dorsal surface. In birds, these have been replaced with strong plates of fused vertebrae, which protect the skeleton during contraction of the powerful flight muscles.
CLASSIFICATION
As in other animals, the muscles of birds are classified as smooth or striated and voluntary or involuntary. Many of the muscles that contain smooth muscle fibers are also involuntary, stimulating the movement of the internal organs. Many muscles with striated fibers are skeletal and are associated with bone movement, so they are under voluntary control.
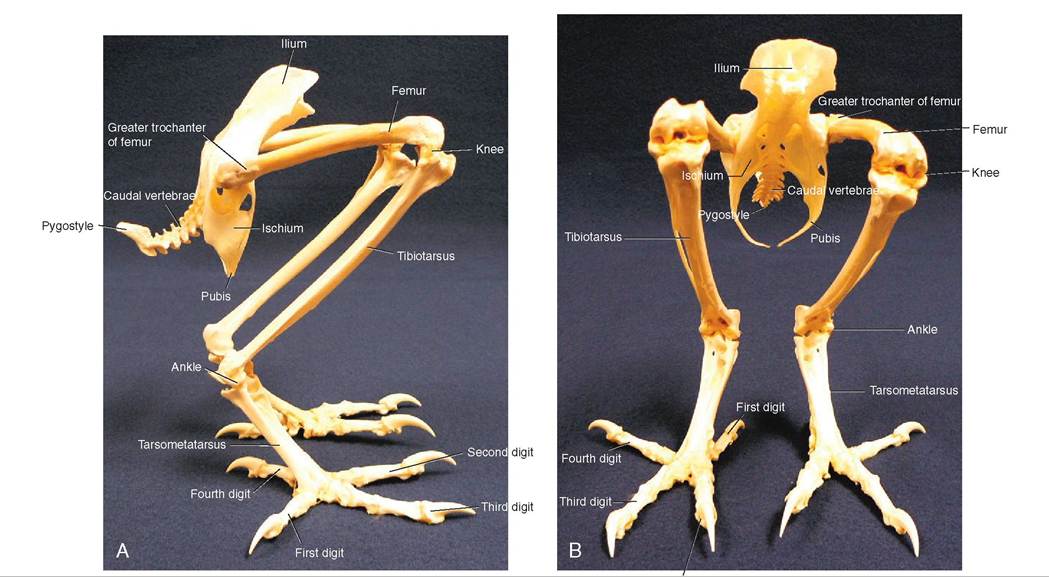
FIGURE 21-16 Anatomy of the leg bones and pelvic girdle of a Great Horned Owl. A, Lateral view. B, Ventral view.
∕j CLINICAL APPLICATION
Handling Birds
Understanding bird anatomy is critical to handling them safely and effectively. Different types of bird have different protective strategies and you must quickly restrain them and protect yourself from what can hurt you. For example, heron species will strike at your eyes with their long sharp bills and birds of prey have sharp beaks and talons, both of which they will use to defend themselves. Psittacines have extremely hard, sharp beaks and strong jaws that can inflict serious bite wounds. These birds are often grabbed/restrained by covering the body with a towel, quickly restraining the neck with one hand and gaining control of the legs/feet with the other hand. The wings are contained with aid from the towel.
No matter which species you handle, for your safety and that of the patient, it is critical to have a firm hold. People that are afraid of hurting a bird while restraining it are often the ones who get hurt or allow a co-worker to be hurt. As you will learn later, birds have complete tracheal rings, so neck holds are safe if indicated. One action to be cautious about, however, is applying excessive pressure over the keel. Birds don’t possess a diaphragm (inspiratory muscle), and air is pushed not pulled into the respiratory system. Thus, it is important not to press down on the keel too hard or this will prevent the passage of air.
Cardiac muscle is also striated but has its own intrinsic rhythm that does not require external innervation.
The skeletal muscles of birds can have white or red muscle fibers. Some muscles consist primarily of one type or the other, but many have a mixture of both. People celebrating the Thanksgiving holiday with a traditional turkey dinner are familiar with these fibers as they are often referred to as the “light meat” and “dark meat,” respectively. White fibers are thick and have a low blood supply, have little myoglobin for carrying oxygen, and use stores of glycogen to sustain muscle contraction. They predominate in the flight muscles of shortdistance fliers, such as chickens, quail, grouse, and many other gallinaceous birds that have rapid takeoffs but are capable of only short flights. If these birds are forced to fly repetitively, they quickly become fatigued and cannot fly at all until they recover. In contrast, red fibers are thinner and have a rich supply of blood, fat, myoglobin, and mitochondria. Using these components, they can produce enough energy to sustain muscle contractions for long periods. Red fibers are found in the flight muscles of long-distance fliers, including many species of songbirds, water birds, pigeons, and birds of prey.
WING MUSCLES
Wings possess several pairs of muscles that are each responsible for a specific action or counteraction: raising or depressing the leading edge of the wing, pulling the wing forward or backward, extending or flexing the wing, or controlling movements of the alula bone (thumb) (Figure 21-17). However, the
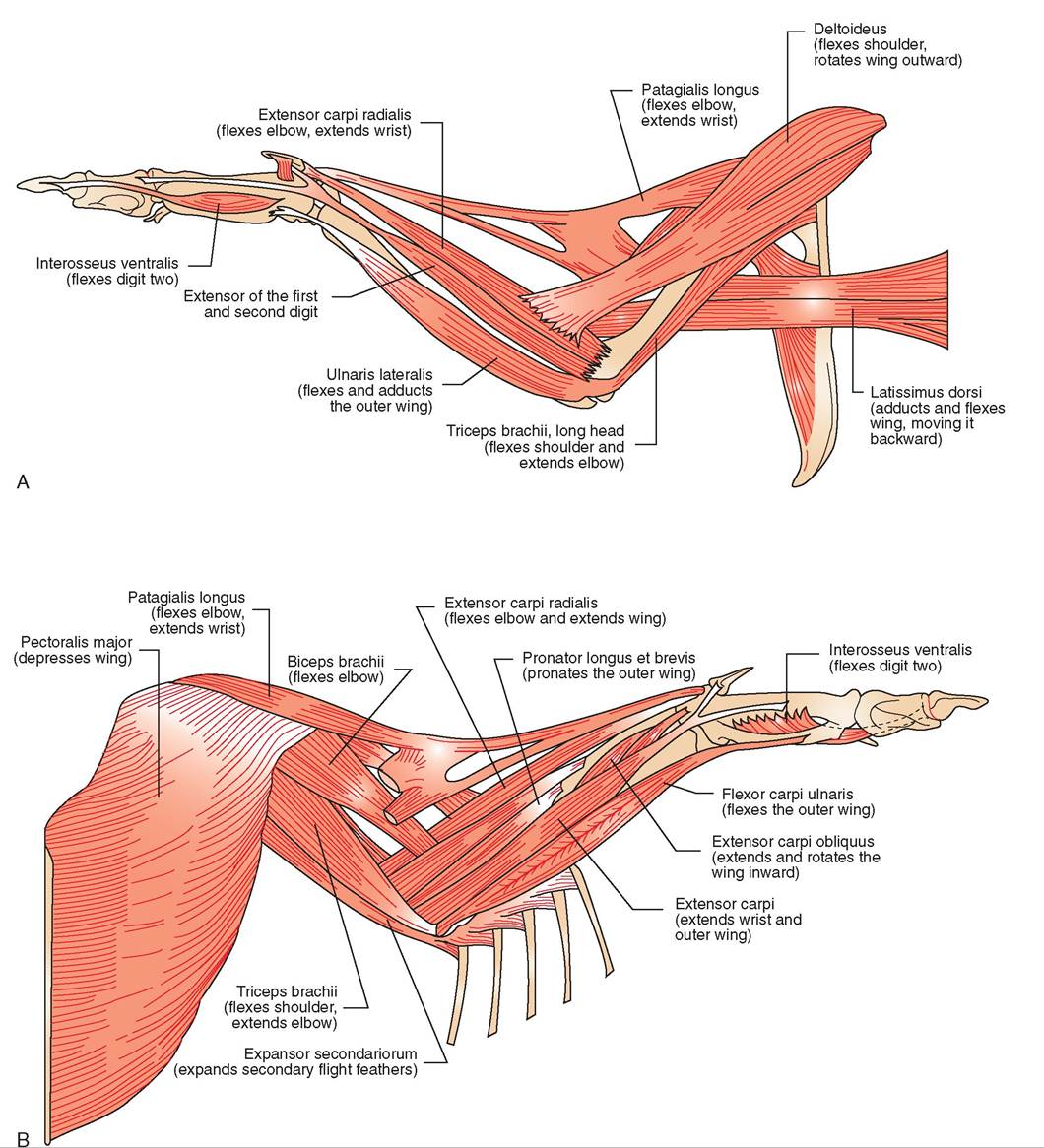
FIGURE 21-17 Wing muscles and their function. A, Dorsal view. B, Ventral view.
two most prominent muscle pairs are those responsible for depressing and elevating the wing. Both pairs originate on the sternum but differ in where they insert (Figure 21-18). The larger, more superficial muscle is called the pectoralis, and it inserts on the underside of the humerus. When it contracts, it depresses the wing, causing the downstroke. This stroke requires a large muscle because it works against two resistant forces: gravity and a tight wing structure formed when the leading edge of each flight feather touches the adjacent feather. Because of its relatively large size and accessibility, the pectoralis is the muscle of choice for administering intramuscular injections, such as vitamins and antibiotics.
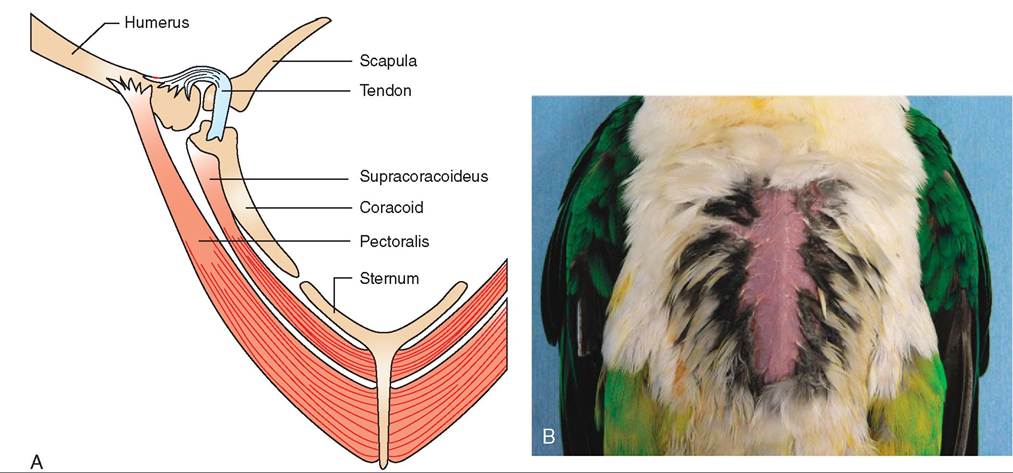
FIGURE 21-18 Flight muscles. A, Cross section. B, Pectoralis major muscles of a White-Breasted Caique.
The smaller, deeper flight muscle is called the supracora- coideus; it turns into a tendon that passes through the glenoid cavity formed by the shoulder girdle and inserts on top of the humerus. When it contracts, it causes the counteraction, which is the elevation of the wing during the upstroke. During this stroke, the flight feathers separate slightly from each other, allowing air to pass through. This creates less resistance and allows the wing to move more easily. In strong, long-distance fliers, the two pairs of flight muscles constitute between 20% and 25% of a bird's weight.
LEG MUSCLES
Like the wing muscles, the leg muscles also have been moved close to the center of gravity. The majority are located in the thigh region over the femur, a smaller number are located over the tibiotarsus, and very few are found over the tarso- metatarsus (Figure 21-19). The large group of muscles over the femur can control movements in the distal leg and toes via strong tendons. For example, in perching species, tendons that control movement of the toes originate from flexor muscles in the thigh and extend over the heel joint into the digits. Extensor tendons run down the front of the tibiotar- sus and metatarsus, whereas the flexor tendons run along the back. The flexor tendons sit in a groove at the top of the metatarsus. When a bird bends its legs to perch, the tendons also bend and pull the toes closed around the perch. This is called the perching reflex and allows a bird to grip its perch firmly while sleeping.
MUSCLES OF THE HEAD AND NECK
The most pronounced muscles of the head are the jaw muscles, which control the beak. The extent of this musculature varies among species and is generally correlated with a bird's diet. For example, species such as parrots use their beaks to crack open large, coarse seeds, so they have relatively large and strong jaw muscles compared with species, such as doves, that consume smaller seeds, or those that consume insects.
A bird has great flexibility in its neck. It has several thin, stringy neck muscles that weave through each other and allow movement in different directions. Some muscles are attached to the connective tissue, or fascia, of adjacent neck muscles. When one muscle is stimulated, neighboring muscles contract, making a variety of movements possible. This is best demonstrated in parrot species that move their heads up and fdtown, le and right, and many combinations thereof.
One highly specialized neck muscle in birds is referred to as a hatching muscle. This muscle, located on the dorsal side of a chick's head, develops during the embryonic stage and is needed to help a chick break out of its shell. It is largest a day or two before hatching, and once a chick reaches the outside world it rapidly atrophies.
From the tip of a bird's beak all the way down to the tips of its claws, a bird's bone and muscle framework is uniquely designed to support aerial locomotion. Now, let us take a peek at the organ systems that actually drive this incredible flying machine.
TEST YOURSELF 21-3
1. Describe the attachment of the skull to the vertebral column. What does this type of attachment provide?
2. List the bones in the avian wing from the shoulder to the wing tip.
3. List the bones in the avian leg beginning at the hip and extending down to the toes.
4. List the two types of skeletal muscle fiber and describe their energy use.
5. Why can a bird perch while sleeping?
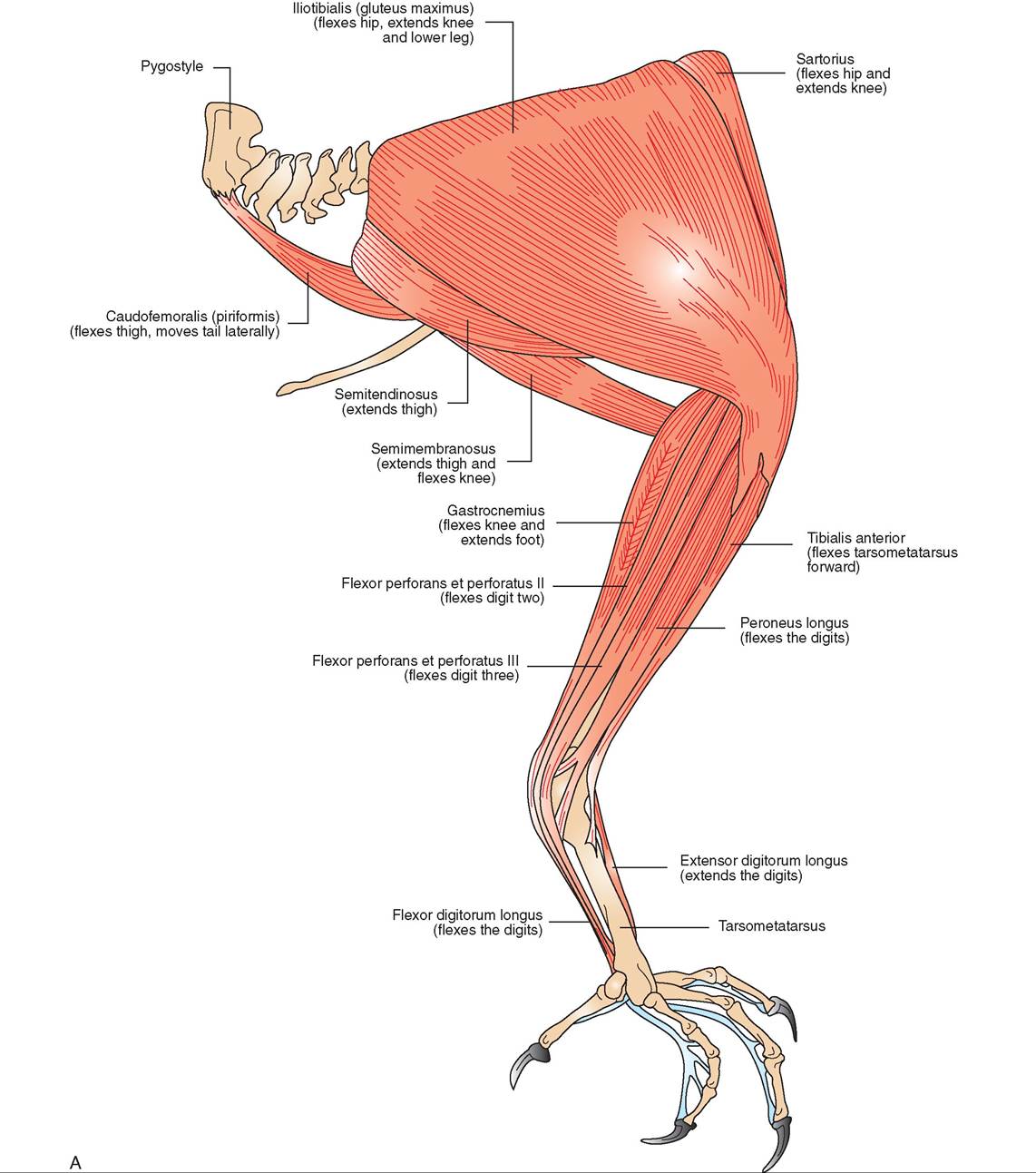
FIGURE 21-19 Leg muscles αed their function. A, Lateral view.
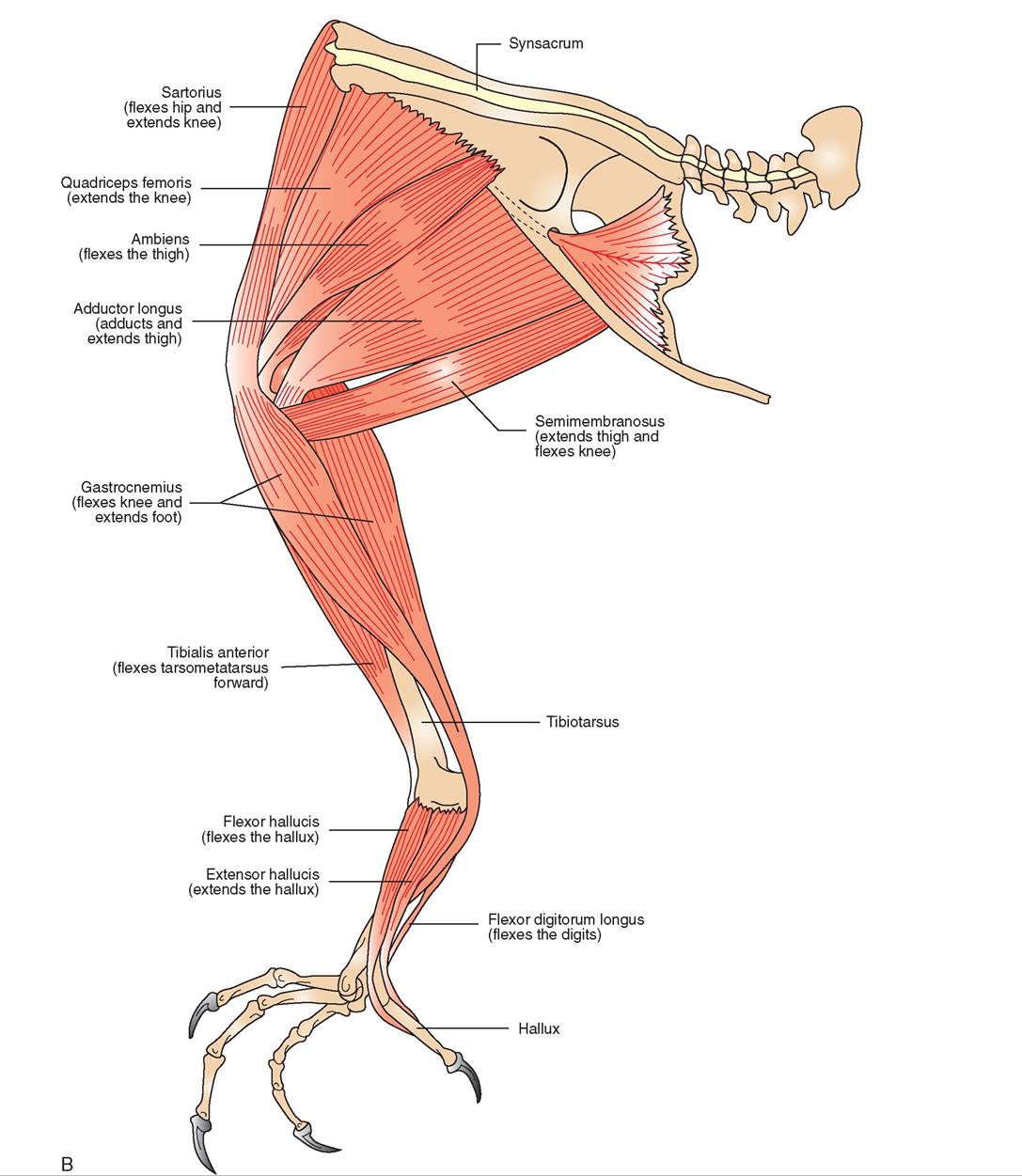
FIGURE 21-19, cont'd B, Medial view.
SENSE ORGANS
The term “bird brain” has been used in reference to a small brain and therefore a lack of intelligence. This usage is misleading; in fact, the avian brain is large in proportion to its body size compared with the brains of all other vertebrates, with the exception of mammals. The location of the control centers within the brain that receive nervous stimuli from the senses is similar to that in mammals (see Chapter 10). In birds, the control centers for vision and hearing are relatively large, whereas those for taste, touch, and smell are relatively small (Figure 21-20).
VISION
The phrase “I'm going to watch you like a hawk” alludes to the fact that the sense of vision is highly developed in birds. An aerial creature needs good vision to fly at variable speeds, find food, escape predators, identify individuals, and participate in courtship rituals. The optic lobes take up the majority of the midbrain, and a large part of the avian skull is devoted to housing and protecting the eyes. The shape of the eyes is determined by the orbits. Unlike mammals, which all have round eyes, bird eyes can be round, flat, or tubular, depending on the species. Generally, diurnal birds have round or relatively flat eyes (hawks and swans, respectively), whereas nocturnal species (owls) have tubular eyes (Figure 21-21). In tubular eyes, the diameter of the pupil is large relative to the size of the retina, and thus more light can be gathered. The eyes fill the eye sockets, leaving little room for muscles or movement.

FIGURE 21-21 Shapes of the avian eye. A, Flat. B, Round. C, Tubular.
In birds, the position of the eyes on the head differs among species and appears to depend on their feeding habits. For example, seed and grain eaters have eyes placed laterally to allow them to view potential predators from many angles. Owls have eyes that primarily face forward, thereby increasing their binocular vision but reducing their overall field of vision. Perhaps the most unusual placement occurs in bitterns, which have eyes placed low on their heads. These heronlike birds feed in shallow water and may use the low eye placement to get a better view of the water. Also, when an intruder approaches, a bittern goes into a protective posture in which it freezes in an erect position with its bill tip facing directly up. This puts its eyes in the best position to view the intruder.
ANATOMY OF THE EYE
The general structure of the avian eye is similar to that of mammals, with a few exceptions (Figure 21-22, A). The avian eye consists of three layers of tissue. The outermost layer is referred to as the fibrous tunic. It consists of the sclera in the back, which functions to protect the eye, and toward the front it becomes the transparent cornea. At this transition point, the sclera is reinforced by a ring of small bones called the sclerotic ring.
The cornea is protected by three eyelids: an upper lid, a lower lid, and a third lid called the nictitating membrane (Figure 21-23). This membrane is thin and transparent and consists of specialized epithelial cells that brush moisture over the eye from the nasal corner laterally. It possesses striated muscles that allow a bird to control its movement voluntarily. In many species of diving birds, such as loons and some ducks, the nictitating membrane has a clear window in its center to act as a contact lens under water.
The middle layer is referred to as the uveal tunic. It is a vascular, pigmented layer consisting of the choroid in back, the iris in front, and ciliary, or striated, muscles. In birds, the iris contains striated muscles that allow voluntary control over the size of the pupils. Thus, a pupillary light response is not a good diagnostic indicator in birds.
The inner layer is referred to as the neural tunic. It consists of the retina, which is composed of photoreceptor cells and neural cells that transmit light images to the brain.
The lens and anterior chamber of the avian eye are similar to those of mammals, as is the posterior chamber (vitreous) with the exception of the presence of the pecten. The pecten is a highly vascular, ribbonlike structure attached to the retina (see Figure 21-22, B and C). It floats in the vitreous humor in the direction of the lens and is believed to distribute nutrition to the eye. More than 30% of traumatized wild birds suffer from hemorrhages arising from the pecten. These are only detected when the intraocular eye structures are examined (Korbel, 2001).
PHOTORECEPTION
Birds possess rods and cones that are similar in function to those in mammals (see Chapter 10). Nocturnal birds, such as many species of owls, have more rods than cones in their
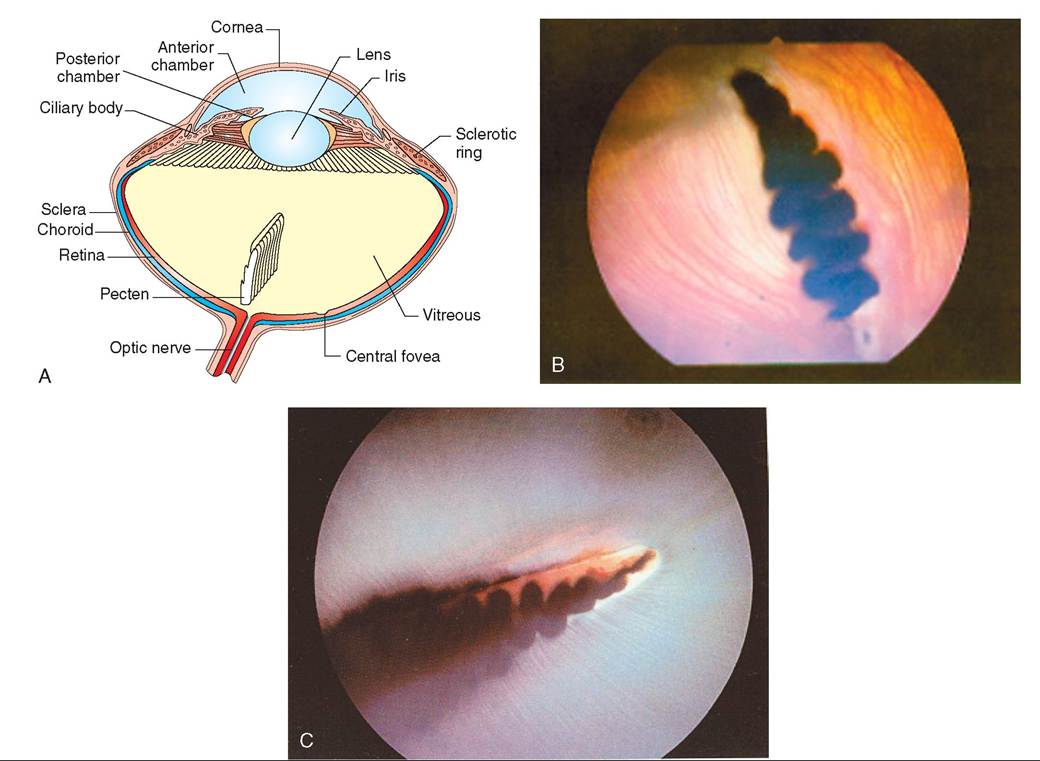
FIGURE 21-22 The avion eye. A, Diagram of transverse section. B, Ophthalmoscopic view through the pupil of intraocular structures in Great Horned Owl (nocturnal bird). C, Ophthalmoscopic view through the pupil of a Red-Tailed Hawk (diurnal bird). (B and C, Courtesy of R. Korbel.)

FIGURE 21-23 Nictitating membranes of a Great Horned Owl. (Photo by Gail Buhl.)
retinas. These rods are specialized in containing a high concentration of rhodopsin, which is the night-vision pigment that aids in absorbing light.
Birds hwe a relatively high level of visual acuity that results from several anatomic features. First, a bird's retina is only lightly vascularized. A reduced number of blood evcersesealssesd interruption of an image as it reaches the
back of' the eye. Second, a bird's retina is packed with photoreceptor cells—sometimes twice as many as in other vertebrates. Think about the difference between a 2 and a 10 megapixel digital camera. The more megapixels a camera rheaast,etrhe g the picture clarity achieved. The avian eye has more cells that receive and transmit information, thus resulting in a clearer image in the brain. A third factor that contributes to the superior visual acuity of birds is the connection between the photosensitive cells and nerve fibers. In many avian species, each cone has a single connection with a bipolar nm;e cell. This means that each cone has individual representation in the brain. In mammals, multiple cones cnonverge o a bipolar cell, and thus their information is pdionogled, lea to a lower level of visual acuity.
Like the eyes of other vertebrates, the avian eye possesses a central, funnel-shaped area containing a high concentration of cones. This area is called the central fovea and is the area of sharpest vision. However, many diurnal birds, such as parrots, hawks, and hummingbirds, also have a second fovea placed temporally. This provides another highly sensitive area and helps in binocular vision.
COLOR VISION
Cone cells in the retina are responsible for processing color images. Each cone contains one colored oil droplet. Diurnal birds typically have yellow, red, green, and orange droplets, whereas nocturnal birds have pale or transparent droplets. How these droplets function is not completely understood, but they have a role in increasing the receptive ability of visual pigments.
VISUAL SPECTRUM
Birds can see a wide spectrum of light wavelengths. Although they cannot see infrared light, many diurnal species can see ultraviolet (UV) light. For example, American Kestrels, which are small falcons (Falco sparverius), can find a mouse by seeing the UV light reflected off its urine. The ability to detect differences in the reflection of UV light is important in other species as well. Some use this ability for distinguishing between ripe and unripe fruit, and others use it to identify males versus females in species that have similar plumages. Ostriches and nocturnal species, such as owls and kiwis, cannot detect UV light.
HEARING AND EQUILIBRIUM
Hearing is another extremely important sense for birds. It is critical for daily activities, such as finding food, hiding from predators, defending territories, and communicating with other members of a family or flock. Although the structure of the avian ear is simpler than that of mammals, it has exceptional acoustic ability.
ANATOMY OF THE EAR
A bird's ears are located on the sides of its head, behind and slightly below its eyes. They consist of three ear chambers: external, middle, and inner (Figure 21-24). The external ear is an opening that funnels sound into the eardrum. It is often bordered with special auricular feathers that protect the ear during turbulent flight and yet still allow sound to pass through.
The external ear is separated from the middle ear by the tympanic membrane. The middle ear consists of a single bone, the columella, which connects to the inner ear and acts as a funnel to transmit sound; this contrasts with mammals, which possess three middle ear bones. The cochlear window is located adjacent to where the columella connects with the inner ear. It has a flexible membrane that protects the inner ear from pressure damage.
The inner ear is similar to that in mammals. It consists of a membranous labyrinth, which functions to maintain balance and equilibrium, and the cochlea, which converts sound waves into nerve impulses that are sent to the brain for processing.
HEARING IN NOCTURNAL OWLS
Hearing in birds reaches its highest level of development in highly nocturnal owls, such as the Common Barn Owl (Tyto alba), Long-Eared Owl (Asio otus), and Northern Saw-Whet Owl (Aegolius acadicus). These species can hunt mice and voles in extremely low light conditions, relying on their incredible hearing ability to capture prey. They have a fleshy flap of skin, called the operculum, at each external ear opening that helps to funnel the sound into the ears (Figure 21-25, A). In addition, their ear openings are asymmetric,
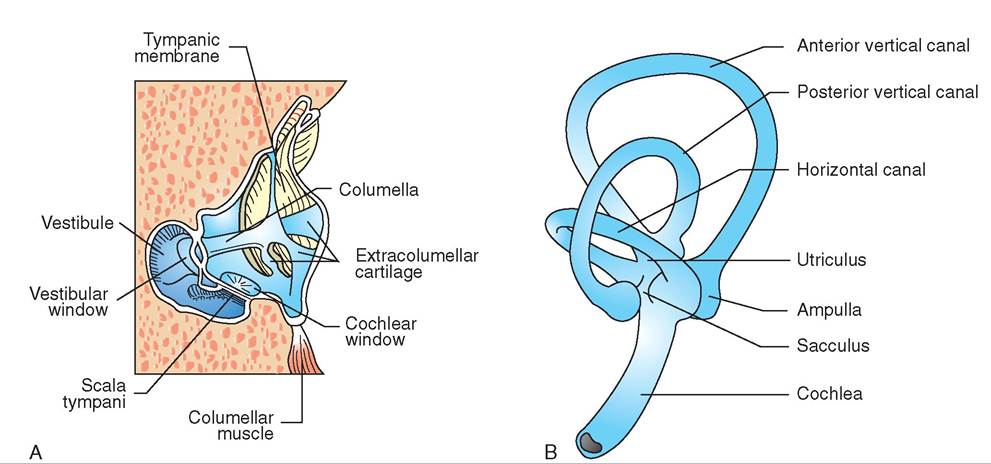
FIGURE 21-24 Anatomy of the avian ear. A, Middle ear. B, Inner ear.
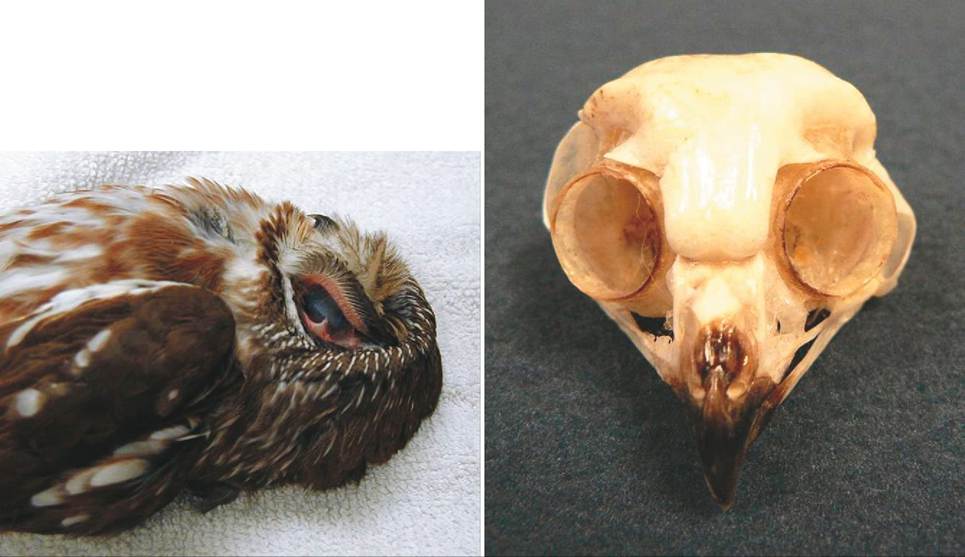
FIGURE 21-25 Ears of an owl. A, Operculum and outer ear of a Northern Saw-Whet Owl (Aegolius acadicus). B, Skull of Northern Saw-Whet Owl showing asymmetric ear placement.
with one slightly above the midpoint of the eye and the other slightly below it (Figure 21-25, B). This feature helps with the vertical location of sound. These nocturnal owls also have large eardrums, columellae, and cochleas, and also a well-developed acoustic center in the hindbrain. In the Common Barn Owl, the number of auditory neurons this center receives is about 95,000, compared with 27,000 in the Carrion Crow (Corvus corone) (Welty and Baptista, 1988).
TASTE
Birds have a relatively poor sense of taste. They possess taste buds, but these are few in number and are scattered on the sides of their tongue and soft palate. Compared with humans, who have about 10,000 taste buds, some parrots have up to 400, and adult domestic pigeons (Columba livia) only have about 50 to 60 (Terres, 1980). Based on experiments with pigeons and chickens, sensitivities and thresholds for bitter, salty, and sour tastes appear to be species specific.
TOUCH
The skin of birds contains two types of sensory nerve ending that respond to pain, heat, cold, and touch. For many species, the sense of touch is important for finding food. Therefore the nerve endings responsible for touch are often prevalent in the tongues, palates, and bills of birds. The first type of nerve ending is called a Grandry’s corpuscle, and groups of these are located in the tongue and palate of many species that dig for food, such as woodcock and sandpipers (Figure 21-26, A). The second type of nerve ending is called a Herbst
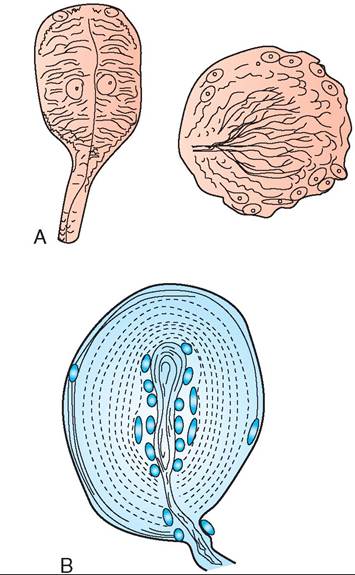
FIGURE 21-26 Touch corpuscles. A, Grandry's. B, Herbst.
corpuscle (Figure 21-26, B). These are also often located in areas of the mouth, such as on the tongues of woodpeckers, on the palates and beaks of ducks, and on the mouth folds of young birds. In addition, Herbst corpuscles are located in the cloaca, legs, wings, uropygial gland, and the bases of many feathers, including the primary flight feathers. These corpuscles are very responsive to even the slightest feather movement. This characteristic explains why birds are sensitive when just the tips of their feathers are touched.
SMELL
The sense of smell varies widely in birds. In a few species, such as the Turkey Vulture (Cathartes aura), Northern Bobwhite Quail (Colinus virginianus), and albatrosses, the sense of smell appears to be well developed and important for
TEST YOURSELF 21-4
1. What are the two most important senses in birds?
2. Which eye structures are found in birds but not in mammals?
3. Where are the bird's ears located?
4. Name the two types of sensory nerve ending in the skin and describe where they are located. locating food. Water birds have a less developed but adequate sense of smell. Goslings learn to accept or reject food by smell at an early age, and Mallard hens emit a breeding odor that stimulates drakes. The sense of smell in passerines and raptors is thought to be poor, but additional research needs to be conducted.
ENDOCRINE SYSTEM
The function of the endocrine system in birds is similar to that in mammals (see Chapter 11). The hormones produced by the glands influence many body systems and control such things as the stress response, courtship and reproduction, body growth, and, in birds, the process of molting. There are seven major endocrine glands; and the pancreas, in addition to having a digestive function, also has an endocrine component (Figure 21-27). As we continue our study of the avian body, we will touch on the function of some of these glands and the substances they produce (Box 21-2).
DIGESTIVE SYSTEM
Birds have a rapid metabolic rate and thus a high energy demand. To meet this need, they have a digestive system that
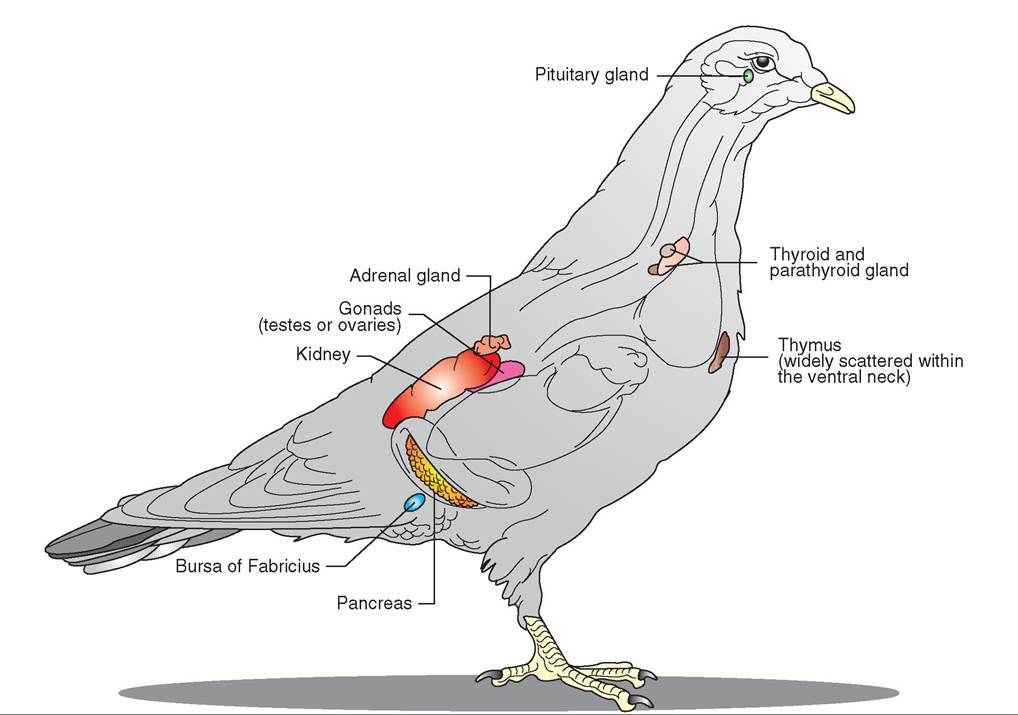
FIGURE 21 -27 Major endocrine glands in a Rock Dove (Columbia livia).
Adrenal Gland
Secretes a stress hormone (corticosterone), sex hormones (androgens in males and estrogens and progesterone in females), and hormones that control concentrations of minerals in the body.
Bursa of Fabricius
Stimulates production of antibodies and lymphocytes.
Gonads
Testes in the male produce testosterone, ovaries in the female produce estrogens and testosterone. These regulate the secondary sex characteristics and control behavioral responses to the opposite sex.
Pancreas
Synthesizes hormones that regulate blood sugar and sugar metabolism in the liver (insulin, glucagon, and somatostatin). Also produces pancreatic polypeptide that inhibits gastrointestinal motility and secretion and induces a sense of satiety.
Parathyroid Gland
Produces parathormone that regulates calcium and phosphorus levels in the body.
Pituitary Gland
Anterior lobe secretes hormones that regulate other glands: thyroid-stimulating hormone (thyroid), adrenocorticotropic hormone (adrenal glands), follicle-stimulating hormone, luteinizing hormone, prolactin (female reproductive system). Posterior lobe secretes antidiuretic hormone to conserve water in the kidney and oxytocin to stimulate uterine contractions for egg laying.
Thymus Gland
Stimulates production of antibodies and lymphocytes.
Thyroid Gland
Secretes thyroxin to regulate growth of the body and feathers and may stimulate the migration urge.
can absorb energy from foods in a rapid and efficient manner with relatively little waste. Depending on the species of bird and its diet, birds assimilate between 60% and 99% of the energy in the food they consume.
ANATOMY
The basic anatomy of a bird's digestive system is similar to that of reptiles and mammals. However, it has been refined and adapted to meet the high and variable energy demands of different bird species (Figure 21-28).
BEAKS AND BILLS
The beaks of birds vary with their diet and foraging strategies. Seed eaters have a thick beak that acts as a forceps and crushes their food; woodpeckers have a heavy, blunt beak that acts as a chisel to bore holes; raptors have a sharp- edged, hooked beak for tearing meat; and shorebirds have a long, delicate beak to probe for food in sandy areas. Beaks enable birds to find, grab, and sometimes kill food items and to tear food into smaller pieces to begin the digestive process.
MOUTH
A bird's mouth consists of a hard upper palate, a soft lower palate, a distinctive tongue, salivary glands, and scattered taste buds (see Figure 21-37). In some species of finches and pelicans, the soft palate enlarges to become a pouch for temporary storage of food. The tongue aids in manipulating food and moving it to the back of the mouth for easy swallowing. In parrots it is highly muscular, but in many other species it has few muscles and is moved by muscles of the jaw apparatus. Most birds have salivary glands located in the back of the mouth (pharynx), but seed eaters also have them in the soft palate. In these birds, the saliva moistens and lubricates dry seeds, and the glands secrete a starch-digesting enzyme. Some species, such as swifts and swallows, use dried saliva to build nests, and woodpeckers use their sticky saliva to hold on to insects.
ESOPHAGUS
The esophagus is a somewhat muscular tube that extends from the pharynx to the stomach along the right side of the neck. The lining contains mucous glands that lubricate food to facilitate its passage into the stomach. In several species, the esophagus expands over the interclavicular space to create a crop, which is a storage pouch for food. This crop can be a dilation of the esophagus, as seen in some fish eaters; a single pouch, as seen in parrots, grain eaters, and hawks; or a double pouch, as seen in pigeons. Birds that have well- developed crops generally eat a few larger meals per day instead of foraging and nibbling all day long.
Little if any digestion occurs in the crop. Its main function is to store, lubricate, and regulate the passage of food. In some species, it is modified for additional purposes. For example, in pigeons and doves, the mucosal lining of the crop thickens at breeding time and is broken down to form “pigeon milk” that is fed to recently hatched chicks. Also, the crop lining in insect-eating birds consists of a heavy epithelium to give protection from insects that are swallowed alive.
STOMACH
A bird's stomach consists of two separate components: the glandular stomach and the muscular stomach. The anterior, glandular stomach is called the proventriculus. This structure is unique to birds and is the organ in which chemical digestion begins. Its mucosa consists of columnar epithelial cells and mucosal glands that produce mucus to moisten food. The submucosal layer possesses digestive glands. These glands secrete pepsin, which begins the breakdown of proteins, and hydrochloric acid, which increases the acidity of
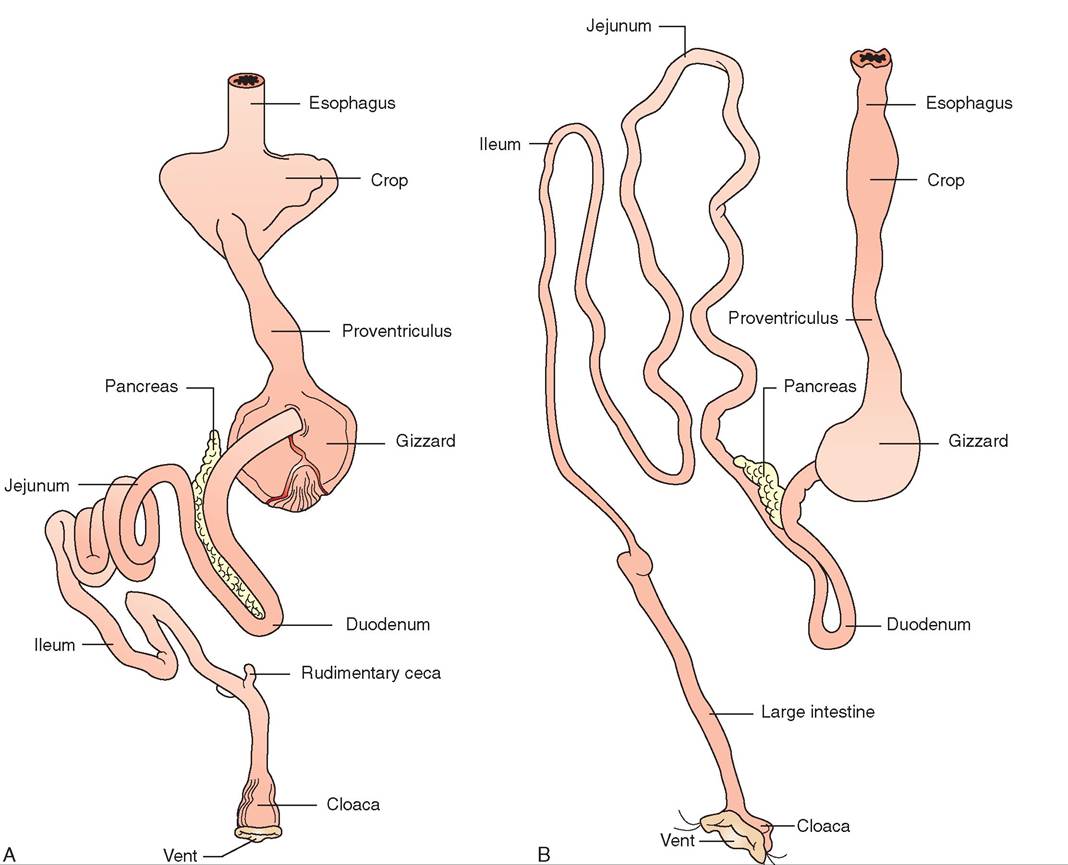
FIGURE 21-28 General diagrams of avian digestive system. A, Rock Dove. B, Hawk.
the stomach to enhance the action of digestive enzymes. In birds, the gastric juices can have a pH between 0.7 and 2.5.
The muscular stomach is called the ventriculus or gizzard. It consists of distinct bands of striated muscles that work to grind food components, such as bones, scales, and nuts. Also, many seed eaters and grain-eating birds actively seek and ingest small pieces of grit to aid in grinding food. In chickens, this grit enhances the digestibility of grain by about 10% (Welty and Baptista, 1988).
Some characteristics of the gizzard are species specific. For example, the thickness of its walls varies with diet. Grain eaters, such as turkeys, have the thickest gizzard walls and can pulverize hard objects, such as steel needles and walnuts. The gizzard walls in carnivores are relatively thin, and those in omnivores are quite variable. In addition, the gizzards of owls, hawks, swifts, grouse, and herons grind indigestible food components into a pellet that is then regurgitated (Figure 21-29). Egestion of pellets in these species can be used as a clinical indicator of normal gastrointestinal motility.
LIVER
The liver in birds is bilobed, and the right lobe is usually larger than the left. The liver stores excess fats and sugars, makes certain proteins, produces bile to neutralize the stomach acid and emulsify fats, and excretes waste products from the blood.
PANCREAS
The pancreas is a relatively large gland in birds and rests in the loop of the duodenum. It is larger in fish eaters and grain eaters and smaller in carnivores. It serves both an exocrine and endocrine function (see Chapter 11). In birds, the endocrine portion of the pancreas occupies more tissue mass than in mammals, and the distribution of endocrine cells within the pancreas is more random.
DUODENUM
The duodenum, or small intestine, is the major organ responsible for the digestion and absorption of nutrients. In meat and fruit eaters, it is relatively short and thin walled. In seed eaters, it is long with several loops. In fish eaters, the duodenum is also relatively long but small in diameter.
CECA
Ceca are paired sacs located at the junction of the small and large intestines in some species. Although their function continues to be studied, they appear to be important for water reabsorption and the bacterial fermentation of
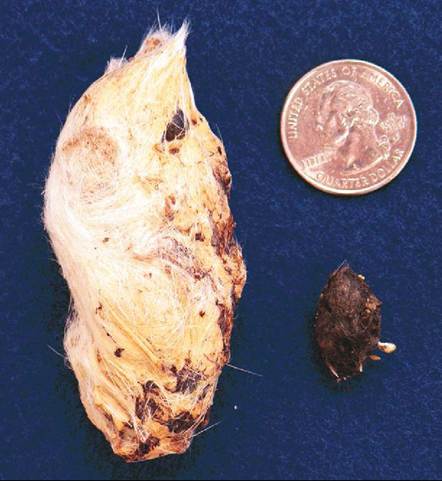
FIGURE 21-29 Regurgitated pellets. A, Snowy Owl (Nyctea scan- diaca). B, Northern Saw-Whet Owl.
cellulose. The output from these sacs is dark brown and moist and has a distinctive odor. It is excreted a few times per week, independently of the intestinal fecal material. Ceca are present in ducks, geese, Galliformes, and owls but are absent in other species, such as parrots, hawks, passerines, and woodpeckers.
LARGE INTESTINE
The large intestine is the segment that extends from the end of the small intestine to the cloaca. Its major role is the reabsorption of water and minerals.
CLOACA
The cloaca is located at the end of the digestive tract and is divided into three sections. The anterior section is called the coprodeum and receives excrement from the intestine. The urodeum receives discharge from the kidneys and genital ducts. The posterior proctodeum is accessed by the other two sections, and it stores the excrement (see Figure 21-40). It is closed by a muscular anus that has powerful ejection muscles for the elimination of waste products through an opening called the vent (see Figure 21-28). The waste products are organized into a mute, consisting of a dark fecal center surrounded by a ring of urates (Figure 21-30).
TEST YOURSELF 21-5
1. Which endocrine gland secretes hormones that regulate molting and the migratory urge?
2. List the endocrine and exocrine functions of the pancreas.
3. On which side of the neck is the esophagus located in birds? Does this differ from mammals?
4. List the two separate components of the avian stomach and their functions.
5. What is a mute? What can it tell us about the health of a bird?
FIGURE 21-30 Normal mutes. A, Blue and Gold Macaw. B, Red-Tailed Hawk.
CLINICAL APPLICATION
Mutes: A Diagnostic Tool
Evaluation of a bird’s mutes is an important diagnostic tool in assessing overall health. In many species the mutes normally have a dark fecal center surrounded by a white ring of urates. However, the color and consistency of a bird’s mutes can be altered by diet, parasites, or disease, and any change in an individual bird’s normal excreta should be investigated to discover the cause. For example, parrots fed a fruity, pelleted diet often have a variety of colors to their mutes, and hawks fed day-old cockerels will have gold fecal centers in their mutes instead of dark ones. These are normal changes and can be explained by the diet. Birds suffering from the actions of some intestinal parasites, such as coccidia, often have a tinge of green in their output, and sometimes blood. An infection with clostridial bacteria often presents as a copious amount of light brown runny mutes. These conditions need to be diagnosed and treated.
Obtaining a complete patient history is important in evaluating these clinical observations. This is easy in the case of pet birds; however, for wild birds with unknown histories, extensive laboratory testing may be necessary to explain abnormal output from the digestive and urinary systems. Tests performed often include fecal analysis for parasites, fecal Gram stain for bacteria, a complete blood count, and a chemistry profile to assess organ function. Blood lead levels are also checked in wild species highly vulnerable to lead poisoning, such as Bald Eagles (Haliaeetus Ieucocephalus), Golden Eagles (Aquila chrysaetos), and vultures, as well as in pet birds.
CIRCULATORY SYSTEM
It has been said that the way to a man’s heart is through his stomach. Although this phrase does not directly refer to the physiologic functions of the two organs, it can be used to explain an important connection between them. The digestive system, as mentioned previously, functions to break down foodstuffs into nutrients that can be absorbed into the blood. From there, the circulatory system takes over to deliver nutrient-rich blood to the tissues and remove metabolic waste. The circulatory system also functions to carry oxygen, minerals, and hormones to cells; in addition, blood helps to control and prevent diseases, and to maintain body temperature.
ANATOMY
HEART
In birds, as in mammals, the driving force behind this delivery system is a four-chambered heart that consists of a right atrium, right ventricle, left atrium, and left ventricle (Figure 21-31, A). The right side of the heart is smaller and less muscular, pumping blood only to the lungs. The left side is larger, and it has well-developed muscles that pump blood to the rest of the body. The heart is located in the cranial portion of the thoracoabdominal space (Figure 21-31, B). It is enclosed by a thin, fibrous pericardial sac, which contains fluid that aids in the lubrication of the heart muscle. This sac adheres to several internal surfaces to keep the heart anchored in place.
VESSELS
The heart is supported by a group of vessels that provide channels for the passage of blood. Arteries carry oxygenated blood from the heart to the tissues, and veins carry blood containing metabolic waste products away from the tissues and back to the heart (Figure 21-32, A). Capillaries are small vessels in which the exchange of gases and nutrients occurs. To meet the specific demands of the avian
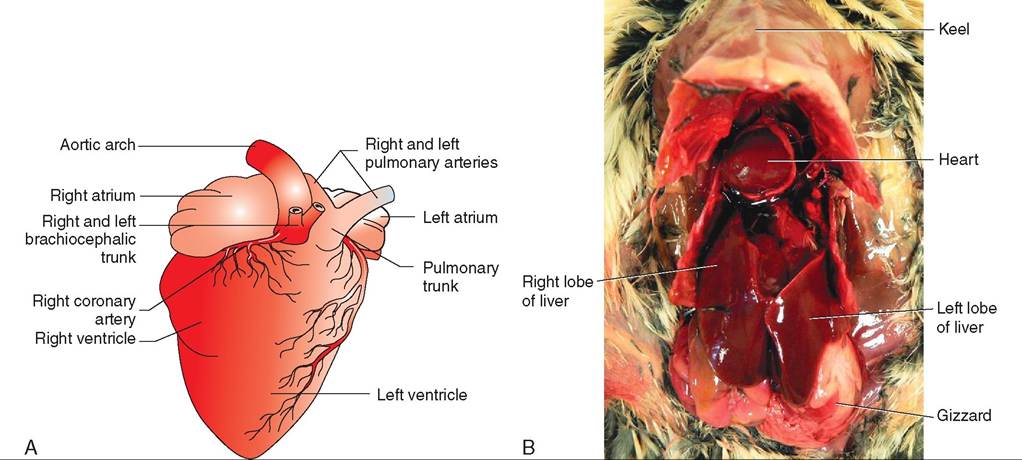
FIGURE 21-31 The avian heart. A, Diagram of ventral surface. B, Location of the heart in a Coturnix Quail (Coturnix Iaponica).
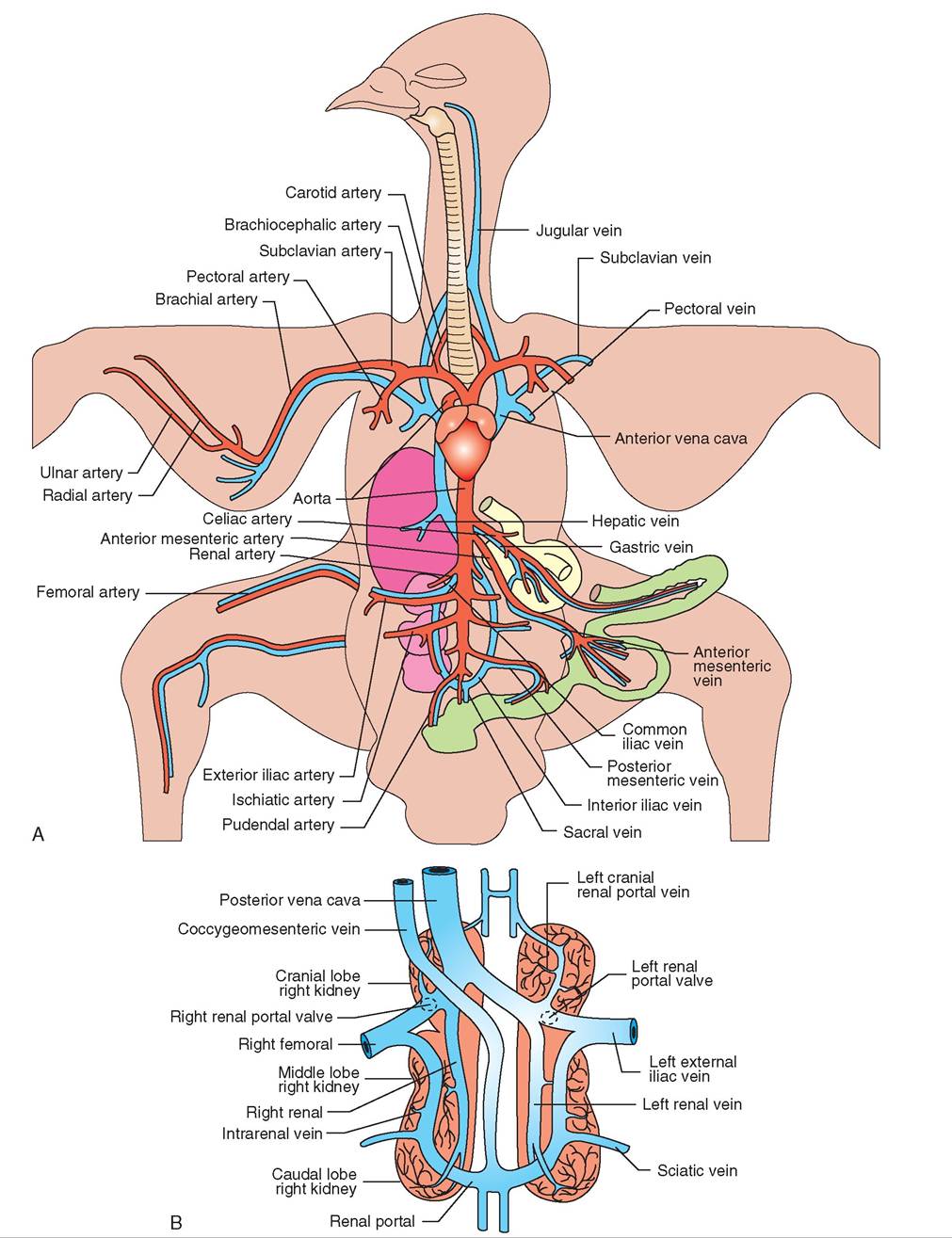
FIGURE 21-32 Vasculature of the avian circulatory system. A, Blood vessels. B, Renal portal system.
body, some of these vessels are highly specialized in the following ways:
• The pectoral and brachial arteries, which provide blood to the flight muscles and wings, respectively, are relatively large.
• Birds possess a renal portal system (Figure 21-32, B) that begins and ends in a network of capillaries. Blood returning from the extremities via the iliac veins travels to the kidneys. Valves at the junction of the iliac veins and renal (kidney) veins steer blood either to the kidneys, so metabolic waste products can be removed, or directly to the heart via the posterior vena cava.
• Many aquatic and terrestrial species possess a countercurrent system of heat exchange in their lower extremities (Figure 21-33). This system consists of a network of arteries and veins that are placed close together. Heat from arterial blood traveling to the lower extremities is transferred to the cooler, venous blood returning to the heart. Thus blood reaching the lower extremities is cooler, and less of a temperature gradient exists with the environment. This feature reduces the amount of heat loss.
BLOOD FLOW
Birds are active creatures and have a relatively high body temperature—between 37° and 42° C (Ritchie, Harrison, and Harrison, 1994). To maintain this temperature and
generate body heat, they also have a relatively fast metabolism. This places high demands on the circulatory system to deliver oxygen and nutrients to the tissues quickly and efficiently. These demands are met with a relatively fast heart rate; smaller birds have faster heart rates than larger species (Table 21-2) and, consequently, more rapid blood flow. In unstressed chickens, it takes only 6 seconds for blood to make a complete circuit from the heart through the body and back (Welty and Baptista, 1988).
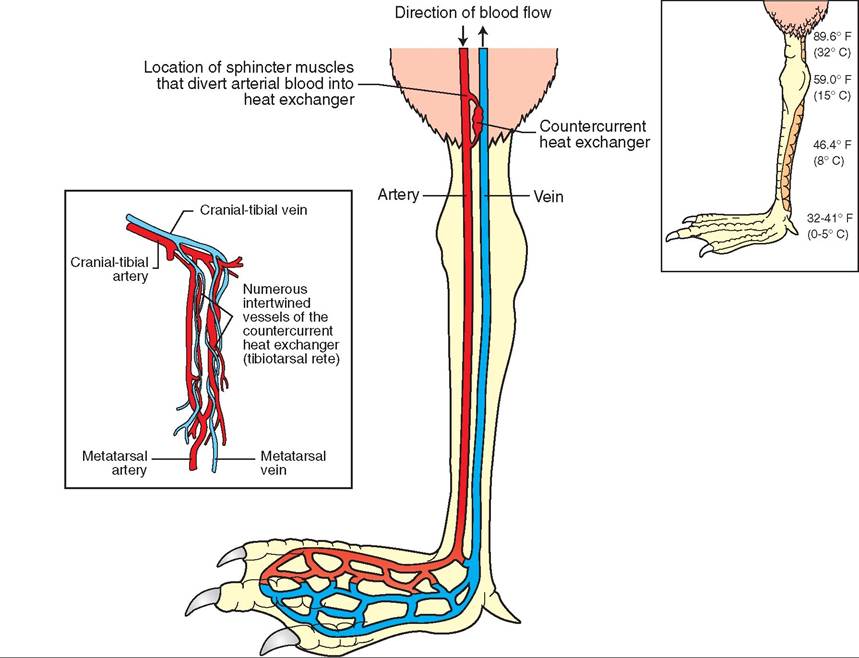
FIGURE 21 -33 Countercurrent heat exchange system in the leg of a gull.
| TABLE 21-2 | Heart Rates in Clinically Normal Birds | |
| BODY | RESTING HEART | RESTRAINED HEART |
| WEIGHT (g) | RATE (beats/min) | RATE (beats/min) |
| 25 | 274 | 400-600 |
| 100 | 206 | 500-600 |
| 200 | 178 | 300-500 |
| 500 | 147 | 160-300 |
| 1000 | 127 | 150-350 |
| 1500 | 117 | 120-200 |
| 2000 | 110 | 110-175 |
| 5000 | 91 | 105-160 |
| 10,000 | 79 | 100-150 |
From Ritchie B, Harrison G, Harrison L: Avian medicine: principles and application, Lake Worth, FL, 1994, Wingers Publishing.
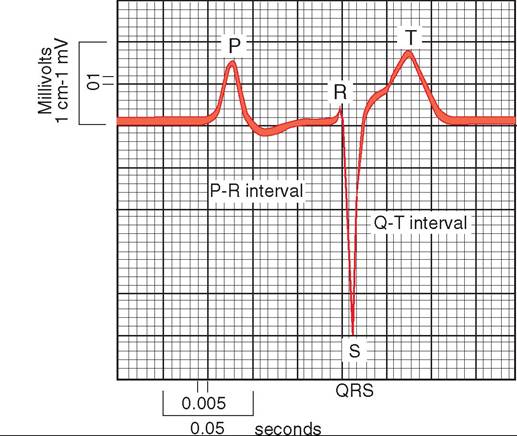
FIGURE 21-34 Electrocardiog ram of a healthy racing pigeon.
ELECTROCARDIOGRAM
As a bird's heart chambers contract and relax, the resulting changes in electrical voltage can be detected by placing electrodes in strategic locations on the wings and legs. The voltage changes are converted into visual peaks and valleys with the help of an electrocardiograph (ECG) machine. The ECG consists of P, QRS, and T waves that correspond to the following muscular activities (Figure 21-34):
P wave—contraction and relaxation of the atria
QRS wave complex—contraction of the ventricles
T wave—relaxation of the ventricles
In birds, the existence of the Q wave is in question. In many species, such as chickens and turkeys, it is believed to be completely absent, but in other species, such as some ducks, it appears to be prominent. The time intervals represented in the ECG are relatively fixed, with the exception of the T to P interval, which changes based on changes in the heart rate.
The ECG is an important tool to monitor a patient's stability during anesthesia and to diagnose malfunctions of the heart and major vessels.
BLOOD
We previously mentioned that the heart drives the circulatory system; the transport vehicle that it drives is the blood. Blood is made up of several components, and it functions to carry nutrients, oxygen, and hormones to cells; to carry metabolic wastes from cells to the lungs and kidneys; to control and prevent disease; and to regulate a bird's body temperature. Blood consists of red cells, white cells, platelets, and plasma (Figure 21-35).
ERYTHROCYTES
Erythrocytes, or red blood cells, are oval, nucleated, and larger than those in mammals (Figure 21-35, A). In most species, they are formed in the bone marrow of adult birds, but in passerines (songbirds), they are formed in the spleen and liver. The red cells possess hemoglobin for carrying oxygen to the tissues. The total number of red blood cells is dependent on several factors, including age, sex, diet, and time of year. In general, the percentage of red blood cells to total blood volume in a healthy adult bird should fall between 35% and 55%.
LEUKOCYTES
Leukocytes, or white blood cells, are important in helping fight disease. In adult birds, white blood cells are primarily produced by the spleen. In young birds, they are also formed by the liver, kidneys, pancreas, and the bursa of Fabricius, which is located on the dorsal wall of the proctodeum in the cloaca. There are several types of white cell, and each type has a different function.
HETEROPHILS. These cells are equivalent to the mammalian neutrophil. They are generally round, have a bilobed nucleus with clumped chromatin, and have rod-shaped, red- orange granules in the cytoplasm (Figure 21-35, A). Heterophils are phagocytic cells that engulf foreign matter. A rise in the number of heterophils is usually seen with the onset of acute diseases.
EOSINOPHILS. These cells are the same as the mammalian eosinophils. They are round cells with a lobed nucleus and large, red-orange, round granules in the cytoplasm (Figure 21-35, B). Their numbers increase in response to allergic reactions and heavy internal parasite loads.
BASOPHILS. These cells are identified by a round, centrally placed nucleus and they stain dark blue (Figure 21-35, C). The function of basophils is still being investigated.
MONOCYTES. These are phagocytic cells that act as a body's second line of cellular defense. The nucleus is often shaped like a kidney bean and can be located centrally or off to one side (Figure 21-35, D). An increase in monocyte production is often seen in cases of tuberculosis and aspergillosis.
LYMPHOCYTES. These cells are the essential components of the immune system. Their eccentrically placed nucleus is round and contains densely clumped chromatin (Figure 21-35, E). They are produced by the thymus and bursa of Fabricius; lymphocytes originating from the bursa of Fabricius produce humeral antibodies to help fight off infections.
THROMBOCYTES
Thrombocytes are nucleated cells that act as platelets. They are smaller than red blood cells and have a large, round to oval nucleus (Figure 21-35, C). They are important in blood clotting and are produced by the bone marrow in adult birds.
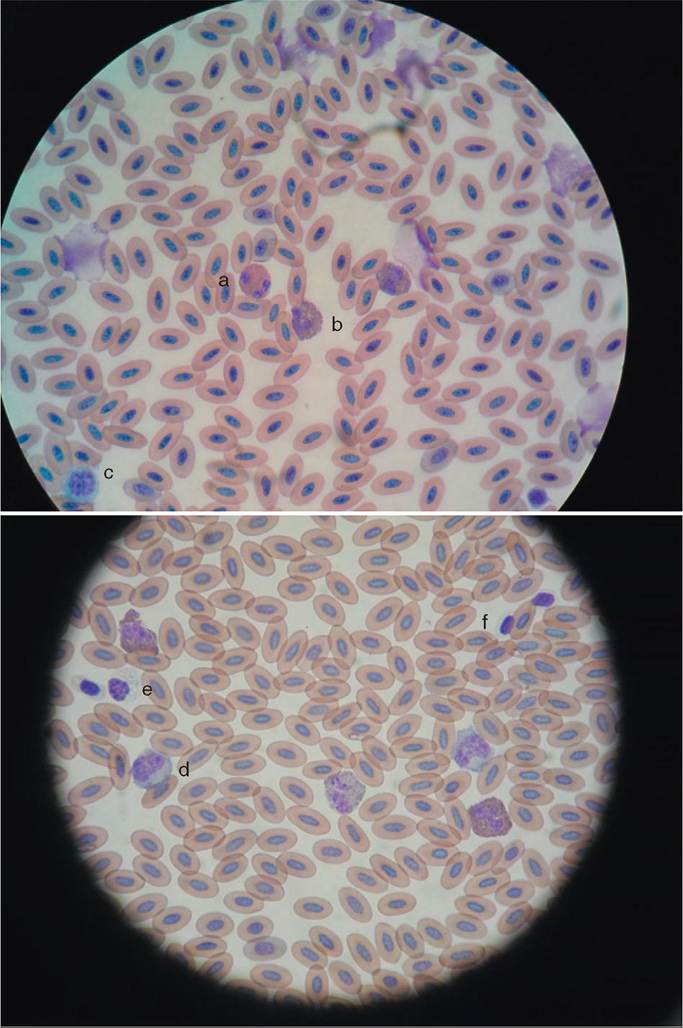
FIGURE 21-35 Avian blood cells. a, Eosin- phil. b, Heterophil. c, Lymphocyte. d, Monocyte. e, Basophil. f, Thrombocyte. (Courtesy Jamie Karlin.)
PLASMA
The plasma is about 80% water. The remaining 20% consists of a variety of substances, including salts, glucose, fats, amino
TEST YOURSELF 21-6 acids, hormones, antibodies, vitamins, enzymes, waste products, and special blood proteins. These proteins are important for providing the osmotic pressure to maintain normal levels of water and blood in the tissues.
1. What is the renal portal system?
2. What is the body temperature range of birds?
3. Describe the components of an avian ECG.
4. How do avian red blood cells differ from those in mammals?
5. List the three veins that are commonly used for venipuncture in birds. Where are they located?
Respiratory system
Because of the fast metabolism and high energy level of birds, the delivery of oxygen and removal of carbon dioxide from the body tissues must be quick and efficient. To accommodate this need, birds possess a respiratory system with highly specialized components (Figure 21-36).
∕ j CLINICAL APPLICATION
Sites of Venipuncture
Blood samples are taken for a variety of diagnostic purposes. In birds, blood can be drawn most easily from one of three vessels: a jugular vein, a brachial vein, or a medial metatarsal vein. The jugular veins are located ventrally, on each side of the trachea. The right jugular is larger than the left and is most commonly used for venipuncture in psittacines. The brachial vein is located on the ventral side of the wing, extending over the elbow and up the humerus. About halfway up the humerus, it joins the cutaneous ulnar vein and increases slightly in size. In raptors, the brachial vein is often used to take blood samples or insert temporary catheters for repeated intravenous treatments. The medial metatarsal vein is located on the ventral medial side of the leg, extending from the metatarsus dorsally over the heel joint. Blood samples are sometimes taken from this site in raptors, but it is more commonly used for waterfowl species.
Venipuncture sites in birds. A, Jugular vein (Northern Goshawk, Accipiter gentilis). B, Brachial vein (Northern Saw-Whet Owl). C, Median metatarsal vein (Red-Shouldered Hawk).
ANATOMY
ORAL CAVITY
The oral cavity contains several structures involved in respiration (Figure 21-37). The glottis is the opening of the trachea, which is located at the back of the tongue. Air is directed to the glottis via the mouth and nasal chambers. These two are linked via the choanae, which are two internal nares that open from the nasal chambers into the roof of the mouth. The larynx, a cartilaginous structure surrounding the glottis, has ligaments and muscle attachments that enable it to act as a valve to prevent solids and liquids from entering the trachea and lungs. It does not function in the production of sound as in mammals.
TRACHEA
The trachea consists of cartilaginous rings that are held together by bands of fibrous connective tissue. In a few species, such as swans and Whooping Cranes (Grus americana), the trachea is very long and coiled. For species that migrate at high altitudes, the long trachea helps provide moisture to the inhaled air and aids in the production of sound.
SYRINX
The enlargement of the trachea above the sternum is called the syrinx. It is essentially the voice box of a bird and contains muscles, air sacs, and vibrating membranes (Figure 21-38). To create sound, air from the lungs and air sacs is forced over the membranes during expiration, causing changes in muscle tension and air pressure. These changes cause vibrations of the membranes. The complexity of a bird's vocalizations depends on the number of muscles present. Birds capable of complex vocalizations, such as many species of songbirds, have an average
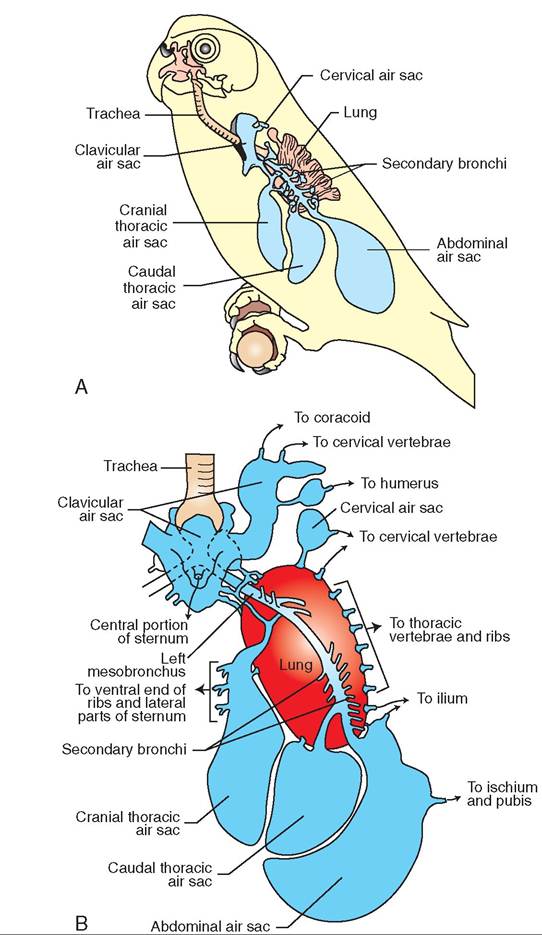
FIGURE 21-36 Diagramof theavian respiratory system. A, Lateral view. B, Ventral view.
of sewn pairs, whereas, on the other end of the spectrum, stories, vultures, and ostriches do not have any. Parrots fall in the middle, with three pairs of muscles in their syrinx.
BRONCHI
At the level of the sternum, the trachea bifurcates into two branches called the bronchi. These branches pass through the ventral side of each lung and end in the posterior air sacs. On« they enter the lung, the bronchi lose their reinforcing cartilaginous rings and are called mesobronchi. The mesobronchi give rise to four to six ventrobronchi, or recondary bronchi, which in turn divide into the parabron- chi.ahbTeronpchair are connected to air capillaries, where
gxcahs aenge occurs.
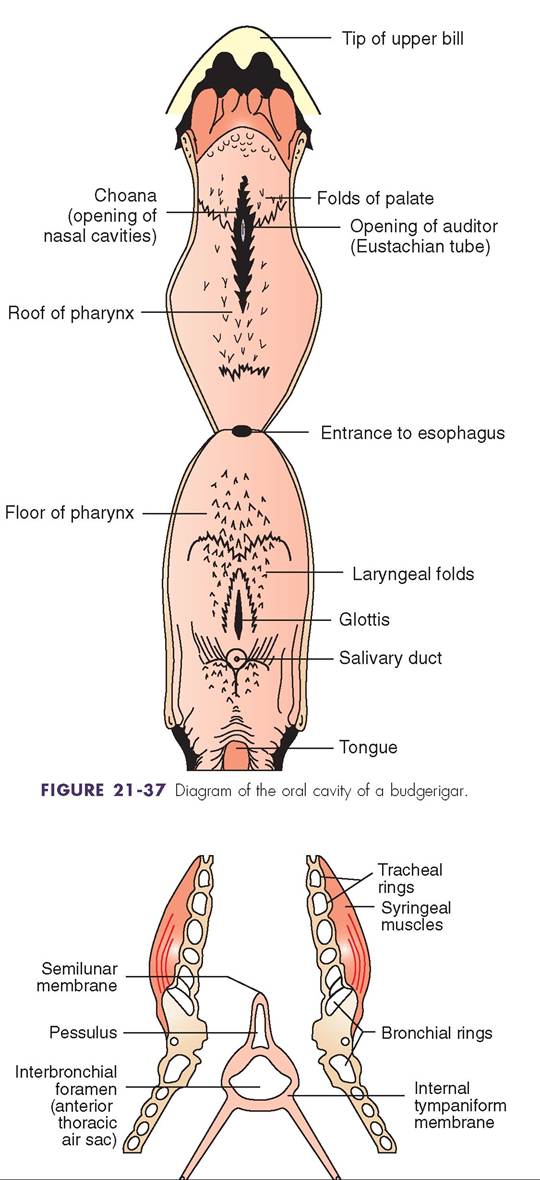
FIGURE 21-38 Crass sectiec ofthe syrinx.
PARABRONCHI
lT, hese smal parallel tubes originate from the ventrobronchi in the lungs and are connected to the tiny openings of air capillaries. The air capillaries, in turn, are surrounded by small blood capillaries. Gas exchange occurs between these trwouopgs of capillaries.
AIR SACS
Air sacs are thin-walled, lightly vascularized, transparent membranes that make up about 80% of the total volume of the respiratory system. There are nine air sacs, four of which are paired. The pairs include cranial thoracic, caudal thoracic, cervical, and abdominal air sacs. The unpaired sac is the interclavicular air sac, which is located in the thoracic inlet between the clavicles. Air sacs are connected to the primary bronchi (abdominal sacs) or Ventrobronchi (cervical, cranial thoracic, caudal thoracic, and interclavicular sacs) and serve the following functions:
• They act as reservoirs for air, and provide warmth and moisture to facilitate the diffusion of air through the lung capillaries.
• They help in thermoregulation, cooling the body by the internal evaporation of water.
• They help provide buoyancy to water birds. Many species of penguins and diving birds have large posterior and abdominal sacs, the volume of which can be adjusted during diving and floating.
Diverticula of some of the air sacs penetrate the skeleton. In many species, the interclavicular sac extends into the humerus bones, sternum, syrinx, and pectoral girdle. The abdominal sacs often extend into the legs and pelvic girdle (see Figure 21-36, B).
LUNGS
In the avian respiratory system, the lungs are relatively small, occupying only about 2% of the total body volume. They are attached to the thoracic vertebrae and ribs and are bright red, highly vascularized, and inelastic. They house the network of blood and air capillaries between which the exchange of gases occurs.
AIRFLOW
Two inhalations and two expirations are required to transport one pocket of air through the entire respiratory system (Figure 21-39). To begin the cycle, the first inhalation involves an expansion of the thoracoabdominal space which creates a gradient that brings air into the body. Birds do not have a diaphragm, the major inspiratory muscle in mammals. Most of the air flows into the posterior air sacs (abdominal and caudal thoracic), where it is warmed and humidified. With the first expiration, this air is pushed into the lungs, where gas exchange occurs. The second inspiration results in the air moving out of the lungs and into the anterior air sacs
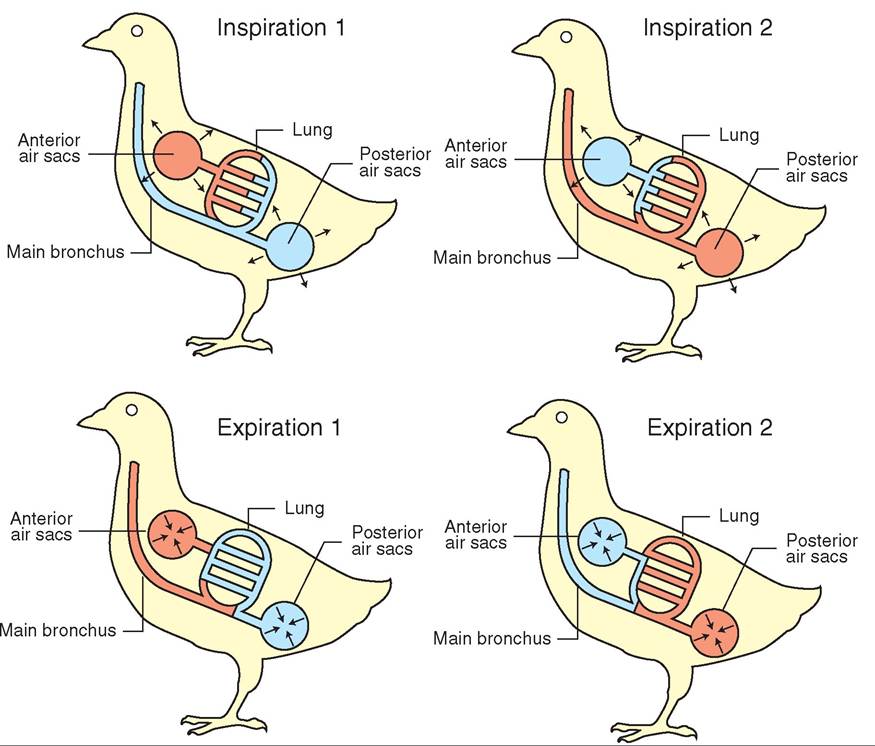
(cranial thoracic, cervical, interclavicular), and the second expiration results in the air leaving the body through the trachea.
The key factor in the flow of air through a bird's respiratory system is that air is pushed, not pulled, into the lungs. More than any other group of animals, birds truly can get a “breath of fresh air.” As previously mentioned, a bird's lungs are inelastic; they do not inflate and deflate as in mammals. Fresh air flows in a continuous, unidirectional path in the lungs and does not get mixed with dirty air containing waste products. This unique feature allows each breath of air to reach the lung capillaries with the maximum amount of oxygen possible—close to the 21% present in atmospheric air. In contrast, the lungs of mammals inflate and deflate, always leaving a quantity of residual air. When new air comes in, it mixes with the air remaining in the lungs, diluting the percentage of oxygen available for gas exchange.
Another unique feature of the avian respiratory system that contributes to its high efficiency is the flow between the air and blood capillaries. The air capillaries are positioned at right angles to the blood capillaries so that carbon dioxide is continually removed from the blood and oxygen is continually added.
RESPIRATORY RATE
The breathing rate of birds varies with species, activity level, age, sex, time of day, and outdoor temperature. Smaller birds usually breathe faster than larger birds, and birds in flight have a higher respiratory rate than nonflying birds. In a thermoneutral environment, chickens at rest breathe 16 to 18 times per minute (Terres, 1980), whereas red-tailed hawks breathe about 40 times per minute (Chaplin, Mueller, and Degernes, 1989). One studyfound that, at rest, house sparrows breathe 50 times per minute; when held in the hand, 102 times per minute; and when released and flying around a room, 212 times per minute. The variability in rate under different conditions can make it difficult to use respiratory rate as a diagnostic tool.
THERMOREGULATION
In addition to the exchange of gases, the respiratory system in birds also helps in thermoregulation. Air in the lungs picks up heat radiated by warm body tissues and blood and removes it from the body during the breathing process. The evaporation of water via the air sacs, lungs, and mouth cavity helps to cool the body to maintain a safe core temperature. This is most critical when outdoor air temperatures rise or when a bird participates in strenuous exercise, such as flight.
To increase the amount of cooling, a bird can increase the airflow over its mouth, pharynx, bronchi, and air sacs by increasing its breathing rate. This often results in panting or gular fluttering. The latter is often seen in doves, Great Blue Herons (Ardea herodias), pigeons, quail, and owls, and it involves rapid vibrations of the upper throat patch.
In addition to a bird's ability to use its respiratory system to remove excess heat, birds can control their body temperature in several other ways. To keep cool, birds bathe or reduce their activity level during the warmest parts of the day. Some species, such as Turkey Vultures and Wood Storks (Mycteria americana), defecate on their legs for evaporative cooling. As mentioned previously, birds also can adjust the position of their body feathers to promote both heat loss and retention. To retain heat, many land and water species possess a special artery-vein arrangement in their lower extremities that acts as a countercurrent heat exchange system to limit heat loss through their bare legs (see Figure 21-33). Birds also change their posture to conserve heat. This may involve perching on one leg to reduce exposure of bare skin to cold air or tucking thee beak behind the feathers on their back. Shivering to increase muscle heat production and moving to more protected locations also help to prevent loss of body heat during times of cold stress. In some smaller species, such as hummingbirds, short-term nocturnal torpor—in which body temperature is decreased by several degrees and heart rate and oxygen consumption are reduced—is another mechanism of heat conservation.
TEST YOURSELF 21-7
1. Where does gas exchange occur in the avian respiratory system?
2. List the nine air sacs and describe their main function in respiration.
3. Do birds have a diaphragm?
4. Describe the path of one breath of air through the respiratory system.
CLINICAL APPLICATION
Toxic Fumes
The large volume of space present in a bird's respiratory tract, the unidirectional flow of air, and the spatial relationship between the air and blood capillaries create an extremely efficient system that is highly sensitive to gaseous molecules in the air. Fumes from disinfectants, such as concentrated bleach; heated Teflon-coated pans or light bulb covers; moth balls (naphthalene); oil-based paints and varnishes; and exhaust fumes can cause illness and even death in birds. Symptoms of exposure to noxious fumes are rapid in onset and may include loss of balance, so a bird may fall off its perch and stand fluffed on the floor. Other symptoms are respiratory rales, or raspy breathing sounds; tail bobbing, in which the tail moves with each labored breath; and severe respiratory distress. Depending on the source of the fumes and extent of exposure, death can occur quickly. Diagnosing the problem relies on obtaining a detailed patient history. Treatment revolves around providing support to the respiratory system, which includes reducing stress, providing oxygen therapy and warmth, placing an air sac cannula to assist with the movement of air, and sometimes administering a mild diuretic to remove fluid from the lungs. However, because of the low rate of survival after exposure to toxic vapors, it is critical to educate all bird owners on the potential deadly effects of fumes to prevent exposure in the first place.
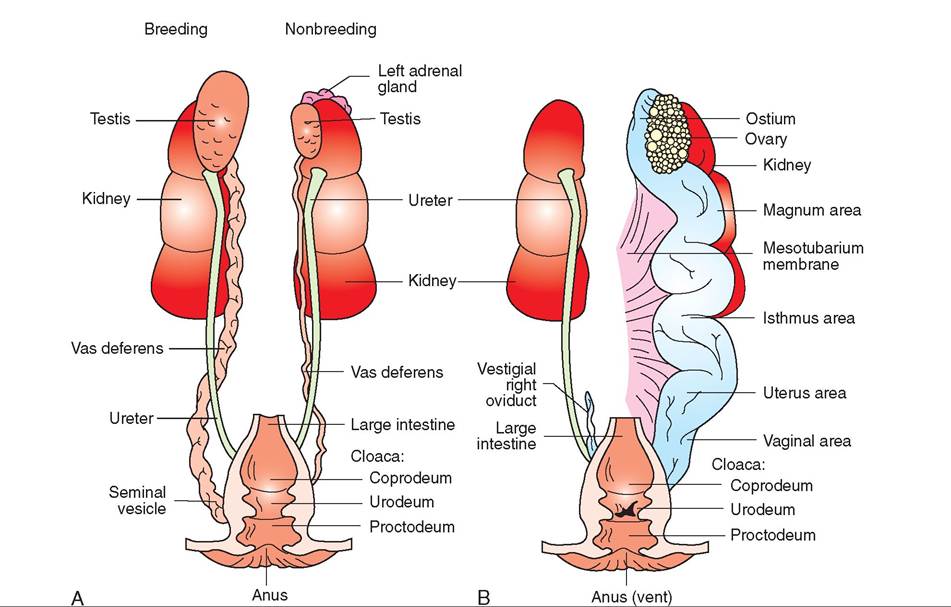
FIGURE 21-40 Avian urogenital system. A, Male. B, Female.
UROGENITAL SYSTEM
The urogenital system encompasses both the urinary and the reproductive systems. Although these systems have very different functions, they are often studied together because of their close anatomic relation in a bird's body (Figure 21-40).
URINARY SYSTEM
Like that of mammals, the avian urinary system consists of two main components: kidneys and ureters. The main function of the urinary system is to remove nitrogenous waste products from the blood and eliminate them from the body; however, it also plays a role in maintaining the osmotic balance of electrolytes and salt in the blood and body tissues.
ANATOMY
KIDNEYS. The kidneys in birds are located dorsally, in the slight depression formed at the level of the synsacrum. They are elongated and consist of three divisions, each of which is layered, with an exterior cortex and an interior medulla. Within each layer, the divisions subdivide into lobules that possess nephrons, which are the “workhorses” of the urinary system. These nephrons radiate around a central vein (Figure 21-41). Unlike mammals, birds do not possess a renal pelvis, and thus the gross anatomy of the kidneys looks more homogeneous.
Each nephron consists of a glomerulus and a tubule. The glomerulus is a filter that removes wastes, salt, glucose, gases, and water from the blood and passes them on to its tubule.
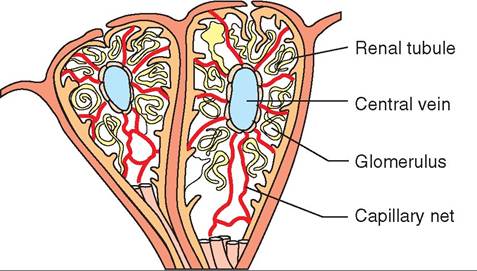
FIGURE 21-41 Internal anatomy of the avian nephron.
The two types of tubules are looped and unlooped. The looped tubules, called the loops of Henle, resemble those in mammalian nephrons and are located in large nephrons in the medulla of the kidney. The unlooped tubules are located in the smaller, cortical nephrons and resemble those of reptiles. In both types of tubule, water, salt, and glucose are selectively reabsorbed into the blood through capillaries, and the waste products are concentrated and passed into collecting ducts that empty into the ureters. Between 66% and 99% of the water filtered into the kidney glomeruli is reabsorbed in the tubules.
URETERS. The ureters are an extension of the main collecting ducts. They are surrounded by smooth muscle and can “milk” urates from the kidney or inhibit their flow into the cloaca. Urine passing through the ureters ends in the urodeum of the cloaca and is either moved upward to the colon and ceca for further reabsorption of water or propelled outward through a bird's vent, which is the sphincter separating the cloaca from the outside.
URINE COMPOSITION
In chickens, the nitrogenous wastes eliminated by the kidneys consist of 75% uric acid, 10% to 15% ammonia, 2% to 10% urea, 1% to 5% creatine, and 2% amino acids (Welty and Baptista, 1988). Clearly, the predominant component is uric acid and not urea, as in mammals. This finding is significant for several reasons. First, uric acid is more efficient in eliminating nitrogen; each molecule removes twice as much nitrogen as a urea molecule. Second, uric acid conserves water because it is relatively insoluble and can be eliminated with only a small volume of water. It takes 60 ml of water to excrete 1 g of urea, whereas it only takes 1.5 to 3 ml of water to excrete the same amount of uric acid. Finally, the production of uric acid is critical to the survival of embryos within the egg. The only waste products that can be eliminated through an eggshell are gases. Other waste products must be stored in the egg throughout the embryo's development. Because urea requires a relatively large volume of water for excretion, the egg would not have enough room to house it. Also, high quantities of urea are toxic and would kill the embryo. As it is, the relatively small amount of nontoxic uric acid produced can be stored in the egg with no ill effects.
The final waste product is eliminated from the body as a paste. It is usually excreted in combination with a small quantity of fecal material from the digestive system. Collectively, the excreted product is called a mute, and, in a healthy bird, it is characterized by a dark fecal center—the color of which will vary with a bird's diet—surrounded by a ring of white urates (see Figure 21-30). Diet, internal parasites, toxins, and certain illnesses can all cause changes in the color of a bird's mutes.
Reproductive system
Finally, we have reached the system responsible for creating the little package of genetic material that will develop into another amazing avian creature. Like most of the body systems in birds, the reproductive system is highly specialized and unique. The entire process of reproduction is periodic and under hormonal control. In temperate-zone birds, increasing daylight stimulates the hypothalamus to secrete substances that travel to the pituitary and cause the release of hormones that directly affect the reproductive process.
ANATOMY
The major reproductive organs are called gonads. In males, these are the testes, and in females, the ovaries. In both sexes, the organ on the left side of the body is larger; in females, the right ovary is rudimentary and does not function at all. During the nonbreeding season, the gonads are relatively small. As the breeding season approaches, hormones stimulate enlargement of the gonads, with up to a 300-fold increase in size of the testes in males. After the season ends, the gonads shrink. This is another adaptation that has allowed birds to reduce unnecessary weight for flight. Many species migrate long distances during the nonbreeding season and do not need to carry large, inactive gonads.
MALE REPRODUCTIVE SYSTEM
The testes in birds are bean-shaped organs located ventral to the anterior part of the kidney (see Figure 21-40, A). Their anatomy and function are similar to those in mammals, because they produce the spermatozoa that are genetically ready to penetrate a developing ovum.
The spermatozoa of birds resemble tadpoles with relatively large heads and mobile tails. They swim out of the testes in seminal fluid produced by tubules in the testes, unlike mammals, whose seminal fluid is produced by accessory glands. The spermatozoa travel down a curvy, ciliated tube, called the vas deferens, to a storage pouch called the seminal vesicle. The temperature in this pouch is about 4° C cooler than the core body temperature and can safely house the heat-sensitive spermatozoa.
COPULATION. To pass sperm to the female, copulation must occur. This behavior is stimulated by androgens, which are male hormones produced in the testes by the cells of Leydig. The androgens travel to the hypothalamus in the brain and stimulate copulatory behavior. Copulation is achieved in one of two ways. In some species, such as ducks, geese, Gal- liformes, storks, and flamingos, a grooved, erectile penis is attached to the wall of the cloaca to help transfer sperm into the female's vagina. However, in most species, sperm transfer occurs when the male and female bring their cloacae into close proximity. In pigeons, 200 million sperm are transferred during a single copulatory event. In the domestic chicken, 8 billion sperm are transferred (Welty and Baptista, 1988).
FEMALE REPRODUCTIVE SYSTEM
Eggs, or ova, are produced from follicles in the cortex of the ovary (see Figure 21-40, B) and are released in a manner similar to those of mammals. Several factors, including increasing day length, stimulate the anterior pituitary gland to secrete follicle-stimulating hormone (FSH), which increases the size of the follicles, and luteinizing hormone (LH), which stimulates both discharge of an egg from its follicle and development of the interstitial cells that produce sex hormones.
OVULATION. Ovulation is the process whereby the ovum leaves its follicle and enters the oviduct. This duct consists of three layers: an outer connective tissue layer, a middle muscular layer that moves the egg down the oviduct by peristalsis, and an inner glandular layer that secretes various substances that are added to the egg during its passage. Lengthwise, the avian oviduct can be divided into the following five sections:
1. Infundibulum—possesses folds that grab an ovum as it comes out of the ovary.
2. Magnum—secretes layers of albumin (egg white) around the egg.
3. Isthmus—deposits the keratin shell membrane.
4. Uterus—deposits watery albumin, a hard external shell, and pigmentation; large and muscular, it is also called the shell gland, and an egg spends the most time here.
5. Vagina—secretes mucus to assist in egg laying; stores sperm for hours to several days.
Eggs come in different colors and may have speckles, blotches, or other irregular markings, depending on how much they move when the tiny pigment glands apply color. During the egg production process, hormones alter a female's blood composition. Estrogen produced from the interstitial cells of the ovary stimulates a 3-fold to 18-fold increase in fatty substances in the blood. Hormones from the thyroid, adrenals, and pancreas are responsible for doubling the blood sugar concentration, and the ovary and parathyroid glands stimulate an increase in blood calcium.
CLUTCH. The group of eggs that a female lays and incubates is called a clutch. Clutch size varies among species but is fairly constant from year to year. Many species lay one egg per day. Others, such as ducks and geese, lay eggs every other day. Some larger species, such as eagles and condors, may lay at 4- to 5-day intervals.
The total number of eggs that a female can lay varies with the species. The two types of egg layers are determinate and indeterminate. Determinate layers, such as budgies, crows, and passerines, have a specific number of follicles that develop in the ovary. Once these are laid, the clutch is complete, even if the eggs are removed. Indeterminate layers, such as parrots, poultry, and birds of prey, can produce more eggs than their normal clutch size and will continue to lay eggs if their eggs disappear. Visual, tactile, and hormonal influences cause them to stop laying when their clutch is complete. This concept has been used to incubate eggs of endangered species artificially, because the eggs can be removed and incubated elsewhere, allowing an endangered hen to lay more eggs.
INCUBATION. Eggs must be kept warm and humidified during their development. An average incubation temperature for many species is 35° C. When a female bird is ready to lay her eggs, the pituitary acts again to secrete prolactin, which decreases the activity of the FSH and LH and promotes broodiness, or incubation behavior. Hormones also stimulate the development of a brood patch, which is an area of skin on the lower abdomen where heat is transferred to the egg. The hen plucks the feathers in this area because of the influence of prolactin, and estrogen stimulates thickening and wrinkling of the epidermis and an increase in the size and number of blood vessels.
The amount of time required for an egg to complete its development varies with the species. Generally, smaller birds have shorter incubation times than larger birds (Table 21-3).
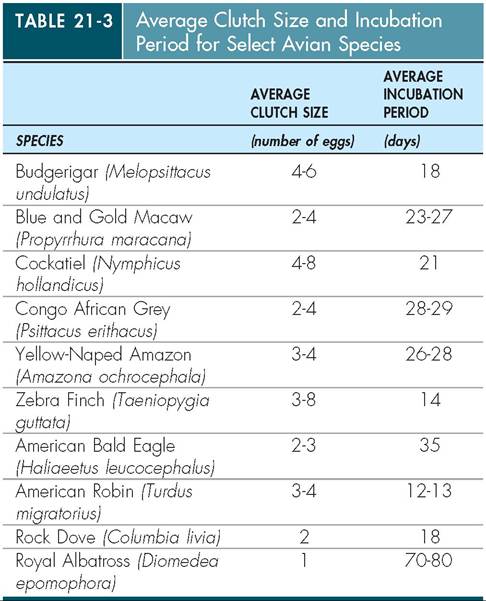
CLINICAL APPLICATION
Egg Binding
Eggs passing through the oviduct sometimes get lodged. This condition is called egg binding, and it is seen most often in small companion birds, such as parakeets, canaries, finches, and cockatiels. Birds suffering from this condition may be sluggish and reluctant to fly or perch, or they may stand with a wide stance, droop their wings, have paralysis of the rear limbs, be anorexic, or have strained abdominal movements. Malformed eggs, excessive egg production, obesity, vitamin deficiencies, inactivity, environmental stress, or malfunction of the oviduct muscles can all result in egg binding. Removing the lodged egg may require drug therapy, such as oxytocin or prostaglandin to stimulate contraction of the uterus, or it may require manual manipulation of the patient. The latter may include massaging the abdomen, administering an enema to lubricate the urodeum, or inserting a speculum into the urodeum to facilitate egg passage. If these techniques are unsuccessful, more intricate procedures may be necessary.
THE CHICK
From hatchlings to adults, the rate of development and life span of birds varies dramatically among its numerous species (Table 21-4). A chick hatches from the egg with the help of powerful neck muscles and a specialized egg tooth on the outside of a chick's bill (Figure 21-42). The neck muscles atrophy greatly after hatching, and the egg tooth disappears.
The sex of the chick is determined by the genetic information passed on by the female. The female chromosome, Z, is
| TABLE 21-4 Life Spans of Common Pet Bird Species | ||
| BIRD | MAXIMUM (YEARS) | AVERAGE (YEARS) |
| African Grey Parrot (Psittacus erithacus) | 50 | 15 |
| Amazon Parrot | 80 | 15 |
| BuHgerigar (Melopsittacus undulates) | 18 | 6 |
| Canary | 20 | 8 |
| Cockatiel (Nymphicus hollandicus) | 32 | 5 |
| Conure | 25 | 10 |
| Domestic Pigeon (Columbia livia) | 26 | 15 |
| Eclectus Parrot | 20 | 8 |
| Grey-CheekeH Parakeet (Brotogeris pyrrhopterus) | 15 | 8 |
| GuolHian Finch (Chloebia gouldiae) | Unknown | 4 |
| Macaw | 50 | 15 |
| Mynah | 8 | 3 |
| Pionus Parrot | 15 | 5 |
| Rainbow Lorikeet (Trichaglassus haematadus) | 15 | 3 |
| Rosella | 15 | 3 |
| Sulfur-CresteH Cockatoo (Cacatua galerita) | 40 | 15 |
| Superb Parrot (Polytelis swainsonii) | 36 | 6 |
| Toucan | Unknown | 4 |
| Zebra Finch (Taeniopygia guttata) | 17 | 5 |
From Ritchie B, Harrison G, Harrison L: Avian medicine: principles and application, Lake Worth, FL, 1994, Wingers Publishing.

FIGURE 21-42 Egg tooth ona recentlyhd tched GroatHorr∣ed Owl. (Photo by Gail Buhl.)
dominant in eggs; the male chromosome, W is recessive. Females can lay two types of egg: those with a male chromosome (ZW), and those without (ZZ). SpCTm all have the same sex chromosome—WW Tlefrrfee only when the female contributes the W chromosome will a male result.
When hatched, chicks differ in the amount of feather cover they have, the status of their eyes (open or closed), and their mobility. The four different classifications of newly hatched chicks are as follows:
• Altricial—Some chicks are hatched with their eyes closed and their skin bare. They require a great deal of care yefore they can leave the nest. Parrots and songbirds fall into this category.
• Semialtricial—At hatching, these chicks are covered with down and are immobile. They may have their eyes open, srounchs as he and hawks, or closed, like owls.
• Precocial—These chicks are covered with downy feath- ievrres, ha the eyes open, and are quite mobile, leaving the nest quickly. Ducks and geese are good examples.
• Semiprecocial—Some species, such as gulls and terns, ore born with a downy covering and open eyes, but nmlyove o a short distance from the nest and are fed by their parents. Chicks develop at varying rates into adult birds.
TEST YOURSELF 21-8
1. What is themajorcomponent inthenitrogenous waste of birds? What arn thn advactagne te producing this typn of waetn?
2. Explain how saorm ard frenmterredfro rs the male to the fnmaln ic birds.
3. List thehivesestiono af the avian ogiductasd their fnchtioce.
4. What isthe ohnrageiecunption temperature?
5. Whatd p bι^oodpatsOand tow done it relate to ichnbatioc?
6. Doehribo tho four hlaeeificatioce of cowly hathhod chicks.
SUMMARY
eWnietrhalthe g description of newly hatched chicks, we have come to the end of our brief study of the anatomy ahnysdioplogy of birds. For most undomesticated species, ahmiceckrges e from its shell and over the next few oweeks t months develops physically and psychologically into a being destined to fly. Its body systems mature with all the specialized features needed to make this one aocstsipble.
Outwardly, the bird is covered in a lightweight coat of feathers that efficiently traps air, providing insulation and enabling lift. Each of the many feather types is important in tcarrying ou a specific function for survival. Only birds have feathers, and it is one of the most obvious features unique thmoe.the T musculoskeletal system of birds is remarkably lfifigchite,net, and strong. Powerful flight muscles are located
vnegntrally alo the central axis of the animal. Dorsally, fused voerrmtebrae f a rigid plate that anchors the skeleton and protects it from the crushing forces of the flight muscles. A reduction in the number and density of bones, many of which are pneumatic (air filled), further optimizes both the rkeletons strength and its lightness. Vision and hearing ayre famousl acute in birds because they rely on these senses ifnorg, mat defending territories, and locating prey. The digestive and urinary systems are efficient in processing food without the long-term storage of weighty excrement seen in mammals. Birds lack bladders. Feces and urates are expelled forcefully together from a single vent and, in some species, indigestible components of food are crushed in and then regurgitated from the gizzard, in the form of a pellet.
eBirds ar metabolically active creatures with a relatively high body temperature, and heart and respiratory rates. Respiration is designed to maximize gas exchange and enables birds to fly at high altitudes without succumbing to hypoxia. Two inhalations and two expirations are needed to circulate one pocket of air through the entire respiratory system. The aviers mo in only one direction through the avian respiratory saythsteerm r than in two directions as seen in mammals. Diverticulae of the respiratory system form a series of air sacs that make up the largest portion of the respiratory system (80%); the lungs make up a relatively small portion. cTshese sa are contiguous with pneumatic bones and add to a bii'd’s buoyancy in water. In addition, the red blood cells aurcelenated and much larger than mammalian erythrocytes, making them particularly efficient at carrying oxygen. Like air through the lungs, oxygen-rich blood moves quickly through the bird. Indeed, it takes only 6 seconds in resting chickens for blood to circulate through the entire body and roeturn t the heart.
Even the reproductive system complies with the avian omission t be as light as possible. During the nonbreeding raantdo rmy ig periods, sex organs such as the testes and
ovaries are atrophied and small. However, when needed, tιttei' migration and during the breeding season, these gonads hypertrophy into much larger, functional sex organs. Like empatinleys,r birds lay eggs. The number of eggs and frequency of egg laying vary among species. Once hatched, precocious chicks can feed themselves whereas altricial young Itiov be fed by regurgitation from adults. Birds do not nurse their young and lack the heavy, milk-filled mammary fglands o mammals.
In all of these ways, birds are truly remarkable. Although there are some mammals, such as bats, that fly, only birds dominate the skies. No other animal can fly as high, as far, and as continuously without rest. Every aspect of avian anatomy and physiology has evolved in some way to perform lteh,is sing miraculous feat.
REFERENCES AND FURTHER READING
Altman R, Clubb S, Dorreoteid G, et o1: Avian medicine and surgery, Philadelphia, 1997, WB Saunders.
Anderson Brown AF, Robbins GES: The new incubation book, Blaine, Wk, 2002, Hancock House.
Calder WA: Respiratory and heart rates of birds at rest. Condor 70(4):358-365, 1968.
Campbell T, Ellis C: Avian and exotic animal hematology and cytology, «1 3, Hoboken, NJ, 2007, Wiley.
Chaplin S, Mueller L, Degemes L: Physiological assessment of rehabilitated raptors prior to release. Wildlife J 12(1):7-8, 17-18, 1989.
Dorst J: Re life of birds, vol 1, New York, 1974, Columbia Univer- srietsys.P
Faaborg J: Ornithology: an ecological approach, Englewood Cliffs, NJ, 1988, Pientice-Hall.
oreethy R: Hw birds work, Gloucester, UK, 1982, Blandford Books. Gill F: Oriiilhology, ed 2, New York, 1995, WH Freeman.
Harrison G, Lightfoot T: Clinical avian medicine, vol 1, Palm Beach, FL, 2006, Spix Publishing.
Harrison G, Lightfoot T: Clinical avian medicine, vol 2, Palm Beach, FL, 2006, Spix Publishing.
King AS, McLeeland J: Form and function in birds, vol 2, London, 1981, Academic Press.
Korbel R: Klinischer untersuchungsgang. In Koenig H, Liebich lit,, edilors: Anatomie und propaedeutik des gefluegels, Stuttgart, New York, 2001, Schattauer Verlag.
Pettingill OS Jr, editor: Seminars in ornithology, Ithaca, NY, 1972, oCroartnoerlyl Lab of Ornithology.
Ritchie B Harrison G, Harrison L: Avian medicine: principles and application, Lake Worth, FL, 1994, Wingers Publishing.
Samour J: Avian medicine, ed 2, Philadelphia, 2008, Elsevier Health Sciences.
Silverman S, Tell L: Radiology of birds: an atlas of normal anatomy aonsidtiopning, Philadelphia, 2009, Elsevier Health Sciences.
Stettenheim P: The integument of birds. In Farner DS, King JR, Cililors: Avian biology, vol 2, New York, 1972, Academic Press.
Terres JK: Re Audubon Society encyclopedia of North American birds, New York, 1980, Alfred A Knopf.
Tully T Dorrestein G, Jones A: Handbook of avian medicine, ed 2, Philadelphia, 2009, Elsevier Health Sciences.
Welty JC, Baptista L: The life of birds, ed 4, Orlando, FL, 1988, Soalluegneders C Publishing.
Whittow GC: Sturkie's avian physiology, ed 5, San Diego, 2000, rAecsasd. emic P