Pregnancy, Development, and Lactation
Thomas Colville
OUTLINE
INTRODUCTION, 487
BREEDING AND FERTILIZATION OF THE OVUM, 487
Erection, 487
Copulation, 488
Transport of Spermatozoa, 488
Capacitation, 488
Fertilization of the Ovum, 489
PREGNANCY AND DEVELOPMENT, 490
The Zygote, 490
Cleavage, 490
Implantation, 491
The Placenta, 491
Structure, 492
Attachment to the Uterus, 492
Gestation, 495
Parturition, 495
Labor, 495
Involution of the Uterus, 496
MAMMARY GLANDS AND LACTATION, 496
Characteristics, 496
Species Differences, 497
Udder of the Cow, 497
Alveoli and Duct System, 497
Mammary Gland Development, 497
Lactation, 498
Colostrum, 498
Maintenance of Lactation, 499
Milk Let-Down, 499
LEARNING OBJECTIVES
When you have completed this chapter you will be able to:
1.
Describe the processes by which spermatozoa move from the male reproductive tract to the oviduct of the female at breeding.2. Describe the processes of capacitation of spermatozoa and fertilization of the ovum.
3. Describe the events that occur between fertilization of the ovum and implantation of the blastocyst.
4. Describe the structure and functions of the placenta and explain the relationship between the fetus and the amniotic and allantoic sacs of the placenta.
5. Describe the structures and functions of the components of the umbilical cord.
6. Differentiate between diffuse, cotyledonary, zonary, and discoid placental attachments.
7. Describe the events that occur in each stage of gestation.
8. Describe the structure and development of the mammary glands.
9. Describe the general composition of colostrum and explain its importance to the health of the neonate.
10. Describe the process of lactation and milk let-down.
11. Describe the key hormones in the female that are involved in the processes of breeding, gestation, parturition, and lactation.
VOCABULARY FUNDAMENTALS
Acrosome ahck-rδ-sδm
Afterbirth ahf-tar-barth
Allantoic sac ahl-ahn-to-ihck sahck
Allantois ah-lahn-to-ihs
Alveolar duct ahl-ve-δ-lahr duhckt
Alveoli ahl-ve-δ-lι
Amnion ahm-ne-ahn
Amniotic sac ahm-ne-aw-tihck sahck
Antibody ahn-te-boh-de
Blastocyst blahs-tδ-sihst
Capacitation kah-pahs-uh-ta-shuhn
Caruncle kahr-uhn-kuhl
Chorion kohr-e-ohn
Cleavage kle-vihj
Colostrum kδ-loh-struhm
Copulation kohp-u-la-shuhn
Corona radiata kuh-ro-nah rad-e-oht-ah
Cotyledon kohd-eh-le-duhn
Cotyledonary placental attachment kohd-eh-le-duh- near-e plah-sehn-tahl ah-tahch-mehnt
Delivery of the newborn dih-lihv-ar-e of the nu-bohrn
Delivery of the placenta dih-lihv-ar-e of the plah-sehn-tah
Diffuse placental attachment dih-fyoos plah-sehn-tahl ah-tahch-mehnt
Diploid chromosome number dihp-loyd krδ-muh-sδm nuhm-bar
Discoid placental attachment dihs-koyd plah-sehn-tahl ah-tahch-mehnt
Dystocia dihs-tδ-shuh
Ejaculation eh-j ahck-u-la-shuhn
Erection eh-rehck-shuhn
Embryo ehm-bre-δ
Female pronucleus fe-mal prδ-noo-kle-uhs
Fertilization far-tihl-ih-za-shuhn
Fetal development fe-tahl dih-vehl-ehp-mehnt
Fetal growth fe-tahl grδth
Fetus fe-tuhs
Gamete gahm-et
Gestation jeh-sta-shuhn
Gland sinus glahnd sι-nuhs
Haploid chromosome number hahp-loyd krδ-muh-sδm nuhm-bar
Implantation ihm-plahn-ta-shuhn
Intromission ihn-trδh-mihsh-uhn
Involution of the mammary gland ihn-vδ-loo-shuhn of the mahm-uh-re glahnd
Involution of the uterus ihn-vδuh-loo-shuhn of the u-tar-uhs
Lactation lahck-ta-shuhn
Male pronucleus mal prδ-noo-kle-uhs
Mastitis mahs-tι-tihs
Meconium meh-kδ-ne-uhm
Milk let-down mihlk leht-doun
Morula mohr-yoo-lah
Multiparous muhl-tihp-ear-uhs
Myoepithelial cell mι-δ-ehp-eh-the-le-ahl sehl
Neonatal period ne-δ-na-tahl pihr-e-uhd
Parturition pahr-chuhr-ih-shuhn
Passive immunity pah-sihv ihm-myoo-nih-te
Placenta plah-sehn-tah
Placentome plah-sehn-tδm
Quarter kwahr-tar
Streak canal strek kuh-nahl
Stages of labor sta-jehz of la-bar
Teat sinus tet sι-nuhs
Trimester trι-mehs-tar
Udder uhd-ar
Umbilical artery uhm-bihl-ihck-ahl ahr-tar-e
Umbilical cord uhm-bihl-ihck-ahl kohrd
Umbilical vein uhm-bihl-ihck-ahl van
Urachus yar-a-kuhs
Uterine contraction u-tar-ihn kohn-trahck-shuhn
Zona pellucida zδ-nuh puh-loo-sih-duh
Zonary placental attachment zδ-nuh-re plah-sehn-tahl ah-tahch-mehnt
Zygote zι-gδt
INTRODUCTION
The processes that lead to a new animal being born are almost impossibly intricate, Unforgivingly sequential, and amazingly elegant.
A multitude of events has to happen at just the right times, and in just the right order, in, and between, two separate animals for breeding and fertilization of an ovum to take place. Then if that minor miracle occurs, a whole new collection of events has to proceed in just the right manner for the fertilized ovum (the zygote) to implant in the uterus, undergo all the stages of growth and development during pregnancy (gestation), and survive the birth process (parturition) that pushes it out of the dark, quiet, moist environment in which it has developed, into the bright, noisy, comparatively dry environment in which it will spend the rest of its life. It has to inflate its lungs for the first time and begin breathing, alter blood flow through its heart and lungs, and make its way to a teat or nipple to begin receiving nourishment via its newly functioning digestive tract.While all this is happening to junior, mom's uterus has to stay quiet as it expands during pregnancy, supply all of junior's needs for growth and development via the placenta, and, at just the right time, contract to start the birth process. Her mammary glands have to develop and be ready to produce their first secretion, colostrum, at just the right time so junior will receive what s/he needs soon after birth. She then has to maintain milk production (lactation) for as long as junior needs it before being weaned onto solid food.
Did you notice how many “and then(s)..." there were in these introductory paragraphs? They give an impression of how complex the processes of breeding, fertilization of the ovum, pregnancy, zygote/embryo/fetal development, birth, and lactation/nursing are. That is what we explore in this chapter.
BREEDING AND FERTILIZATION OF THE OVUM
ERECTION
Erection is the enlargement and stiffening of the penis that prepares it for breeding. It results from a parasympathetic reflex triggered by sexual stimuli. In most species, the important stimuli are olfactory cues (smells) and behavioral changes from the female that signal to the male that she is receptive to breeding.
These cues from the female, and her willingness to participate in breeding, are primarily due to the high level of estrogen hormones in her system. Table 20-1 shows the predominant female hormones at the times of| TABLE 20-1 | Key Female Hormones of Breeding, Gestation, Parturition, and Lactation | |
| HORMONE | SOURCE | |
| Breeding | Estrogens Oxytocin | Ovary Posterior pituitary |
| Gestation | Progestins (Progesterone) | Ovary (and placenta in some species) |
| Parturition | Relaxin ACTH Glucocorticoids Estrogens Prostaglandin F2alpha Oxytocin | Placenta Anterior pituitary (fetus) Adrenal glands (fetus) Placenta Placenta and uterine wall (dam) Posterior pituitary (dam) |
| Lactation | Growth hormone Prolactin Oxytocin | Anterior pituitary Anterior pituitary Posterior pituitary |
breeding, gestation (pregnancy), parturition (birth), and lactation (milk production).
Erection occurs when more blood enters the penis via the arteries than leaves it via the veins. The connective tissue- enclosed erectile tissue becomes engorged with blood, causing the penis to become enlarged and rigid. Mechanically, what happens is that the arteries supplying blood to the penis dilate, increasing the blood flow into the organ. At the same time, the veins carrying blood away from the penis are compressed against the cranial brim of the pelvis by contractions of the ischiocavernosus muscles (part of the roots of the penis). This acts like a tourniquet and decreases the flow of blood out of the penis.
The net effect is that more blood enters the penis than leaves it. This generates hydraulic pressure within the erectile tissue, producing the enlargement and stiffening of the penis that we call erection. In species with a sigmoid flexure in the penis, such as ruminants and the boar, the primary erection mechanism is lengthening of the penis by straightening of the sigmoid flexure. In species with a high proportion of erectile tissue in the penis, such as dogs and stallions, engorgement of the erectile tissue with blood results in increases in both length and diameter.COPULATION
Copulation, or the act of breeding, is allowed by the female during the estrus (heat) period. In most animals, it is done in a standing position with the male mounting the female from behind (Figure 20-1). Exceptions to this include the camelids (camels, llamas and alpacas) which breed in a kneeling (cushed) position (Figure 20-2). Mounting is followed by intromission (insertion of the penis into the vagina), thrusting, and ejaculation.
EJACULATION
Ejaculation is the reflex expulsion of semen from the penis. It is produced by a continuation of the stimuli that produced erection, plus the physical actions and sensations of breeding. The process has two stages. The first involves the movement of spermatozoa from the epididymis, and fluids from the accessory reproductive glands, into the pelvic portion of the urethra. At the same time, the sphincter muscle around the neck of the urinary bladder closes tightly to prevent the movement of semen into the bladder. This is quickly followed by the second stage of ejaculation: rhythmic contractions of smooth muscle around the urethra that pump the semen out into the female reproductive tract.
In most animals, ejaculation deposits semen in the upper portion of the vagina. Exceptions to this include the horse and pig, in which semen is usually deposited through the open cervix directly into the uterus.
TRANSPORT OF SPERMATOZOA
The spermatozoa have to cover a lot of territory to travel from where they are deposited up to the oviducts, where their mission is to seek out the ovum.
The distance is huge on the microscopic scale of these tiny cells. They start actively swimming as soon as they are deposited in the female reproductive tract, but if they were to cover the whole distance under their own power, it would take over an hour for them to arrive in the oviducts. In actuality, they begin arriving within a few minutes after ejaculation. How do they make the trip so quickly? The answer is that they are transported mainly by contractions of the uterus and oviducts and the action of cilia in the oviducts. Copulation causes the hormone oxytocin to be released from the posterior pituitary gland of the female. Oxytocin causes the smooth muscle of the estrogen-primed female reproductive tract to contract, giving the spermatozoa a free ride.The reason for the rush to get the spermatozoa up to the oviducts so quickly is that timing is everything at this point. The spermatozoa must arrive at the oviducts before the ovum does so they have time to undergo a process (capacita- tion) that will enhance their fertility.
Nature has an exquisite method for arranging the proper timing. Breeding is only allowed by the female during the estrous, or heat, period. So spermatozoa enter the female reproductive tract when the oocyte in the follicle is fully developed but has not yet been released. Release of the ovum (ovulation) is delayed until near the end of the estrous period in most species. This nifty bit of traffic control helps ensure that the spermatozoa arrive at the oviducts first and have time to undergo the process of capacitation before the ovum shows up ready to be fertilized.
Capacitation
Capacitation describes a series of changes that spermatozoa undergo in the female reproductive tract that collectively serve to increase their chances of successfully fertilizing an ovum. Many of the changes are subtle, involving adjustments in ion movement through the cell membranes, increases in the cells' metabolic rates, and increases in the rates at which simple sugars are used for energy production.
A more dramatic change is in the acrosome, the digestive enzymecontaining caplike structure that covers the head of the spermatozoa. Capacitation causes exposure of the enzymes that, up until now, have been sealed off. These digestive
FIGURE 20-1 Ciourtship and copulation in No hotne. Note the positionof the animals inthe lap photograph when copulation is occurring. (From McGrnnvy P: Equion bnhavior: a guidn for Vntnrioariaos aod nquion scinotists, nd 2, St Louis, 2013, Elsnvinr Hnalth Scinncns.)
enzymes will help the spermatozoa penetrate through the layers surrounding the ovum to accomplish fertilization.
FERTILIZATION OF THE OVUM
Spermatozoa are preprogrammed to seek out anything large and round and attempt to penetrate it. Many go astray and attempt to fertiIize things other than ova, such as epithelial cells in the oviducts. This is one of the reasons why such a large number of spermatozoa must be introduced into the female reproductive tract in the first place: to increase the odds that one will successfully reach and fertilize (penetrate into) the ovum. A significant number of spermatozoa find amnd swar around the ovum once it moves down into the oviduct. Many begin tunneling through the corona radiata (layers of cumulus cells from the follicle) and zona pellucida (thick, gel-1ike membrane that surrounds the ovum's cell membrane) surrounding the ovum, aided by the digestive enzymes of their acrosomes. Only one spermatozoon, however, will achieve fertilization—physical penetration through the cell membrane of the ovum, delivering its genetic material into the ovum's cytoplasm. Once the head loef a sing spermatozoon has entered the ovum, a change takes place in the cell membrane that blocks other sperma- toomzoa fr entering.

FIGURE 20-2 Camelid copulation position — Bactrian Camels. Note that the female is kneeling in a sternal (cushed) position and the male is kneeling over her. (Photograph by the author and used with permission from the Red River Zoo, Fargo, ND; www.redriverzoo.org)
PREGNANCY AND DEVELOPMENT
THE ZYGOTE
Once an ovum is fertilized, it gets a name change and becomes known as a zygote. Immediately after fertilization, the nucleus of the spermatozoon is called the male pronucleus, and the nucleus of the ovum is called the female pronucleus. Each carries the haploid chromosome number, that is, one half the number in the rest of the body's cells. The male and female pronuclei quickly join together to restore the diploid chromosome number and determine the unique genetic makeup of the offspring. The full genetic map for the new animal has now been established.
CLEAVAGE
As soon as the two pronuclei join to form a single nucleus, each chromosome makes a copy of itself, and the zygote begins to divide rapidly by the normal process of mitosis. This rapid division is called cleavage. Figure 20-3 shows what happens to the genetic material from the ovum and the sperm from just after fertilization up to the first cleavage division. The single cell divides into two cells, which quickly divide into four, then eight, then 16, and so forth. Cleavage occurs so rapidly that the cells of the zygote do not have time to grow between divisions. The number of cells making up the zygote is increasing dramatically, but its overall size is still about the same as the original ovum, even after several days.
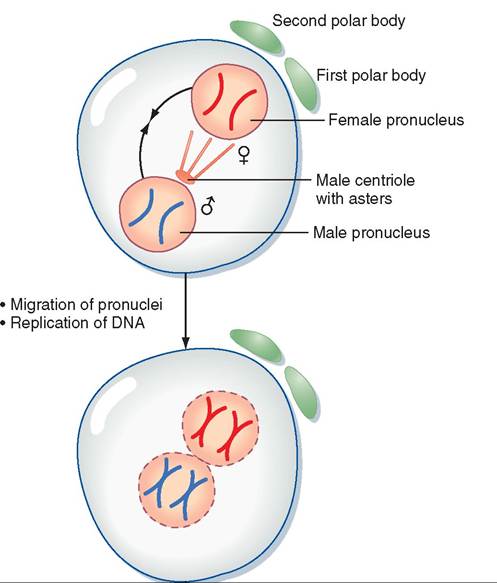
• Breakdown of both pronuclei
• Chromosomes align along metaphase plate
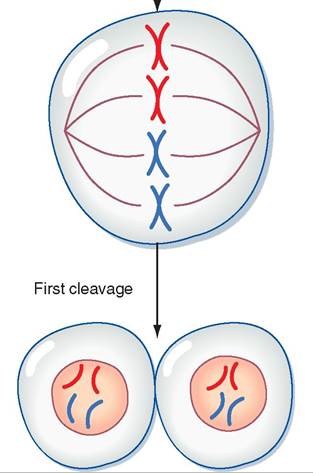
FIGURE 20-3 Overview of events in a zygote from just after fertilization up to the first cleavage division. (Modified from Porterfield SP, White BA: Endocrine physiology, ed 3, Philadelphia, 2007, Mosby. In Koeppen B, Stanton B: Berne & Levy physiology [updated], ed 6, St Louis, 2010, Mosby.)
While cleavage is taking place, the zygote is slowly moving down the oviduct toward the uterus (Figure 20-4). Delicate muscular contractions and the movements of cilia are gently propelling it along. After a few days the zygote is a solid mass of cells that looks like a tiny raspberry; this is known as the
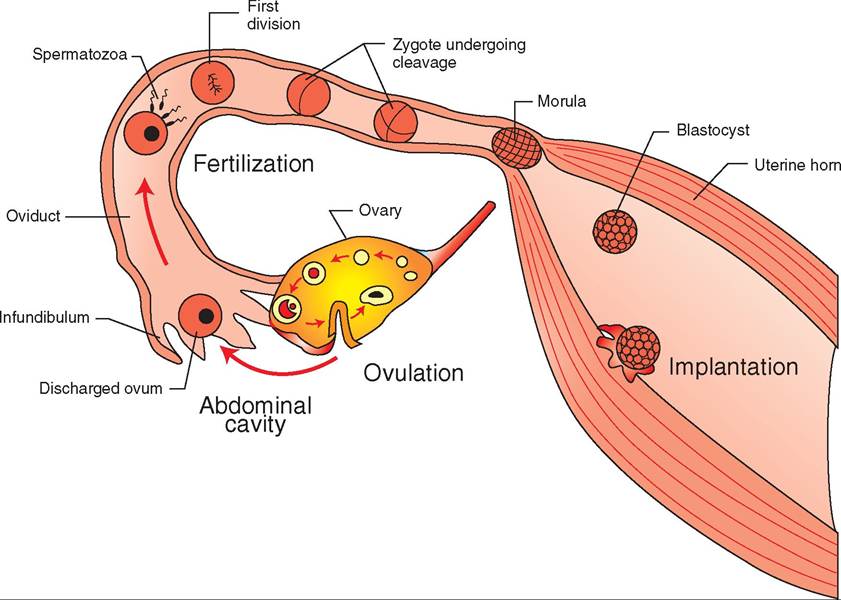
FIGURE 20-4 Fertilization and implantation. After ovulation, the ovum starts moving slowly down the oviduct toward the uterus. A spermatozoon in the oviduct fertilizes the ovum, forming the single-celled zygote. Cleavage of the zygote begins almost immediately as the single cell divides into two cells, two cells divide into four, and so on. After a day or two, the zygote has formed into a solid mass of cells called the morula. The morula continues to develop into a hollow ball of cells, the blastocyst, which enters the uterus and implants in its wall.
morula stage. The cells of the morula continue to divide and gradually form a hollow cavity in the center. By the time it reaches the uterus a few days later, it is formed into a hollow ball of cel Is with a “bump” on one side that eventually forms into the embryo (Figure 20-5). It is now called a blastocyst, and it is ready to implant itself in the lining of the uterus.
IMPLANTATION
Implantation is the means by which the blastocyst makes itself a home by embedding itself in the endometrium (lining) of the uterus. When it comes to rest against the endometrium, enzymes from the blastocyst dissolve away a small pit. The blastocyst implants itself in the pit. In multiparous species (those that normally give birth to multiple offspring), such as swine, the multiple blastocysts randomly space themselves along the horns and body of the uterus as they implant.
During the early part of pregnancy, the developing offspring is called an embryo. Later it is referred to as a fetus. We discuss this further later in the chapter.
Up until now, the dividing zygote has obtained its nourishment by diffusion from the fluid of the oviduct and uterus. By the time of implantation, it has become too large and metabolically active for its needs to be met by this mechanism. A more effective method of supplying oxygen and nutrients and carrying wastes away must be developed. This role is carried out by the placenta, a complex structure that begins to form as soon as the blastocyst implants in the uterus.
TEST YOURSELF 20-1
1. Why is the timing of copulation so important? How is the precise timing accomplished?
2. Describe what happens to a zygote between fertilization and implantation.
THE PLACENTA
The placenta is a life-support system for the developing offspring that surrounds it and connects it with the uterus. During gestation (pregnancy), the offspring leads a parasitic existence as it grows and develops; that is, it receives all the nutrients and other substances it needs to grow and develop from its mother. It also depends on her to dispose of the waste products it produces. Remember that, during the period of pregnancy, the offspring is undergoing incredibly rapid growth, and its cells are differentiating into all of the tissues, organs, and systems needed to support independent life after it is born. Its need for nutrients and waste elimination is huge and grows minute by minute along with its body.
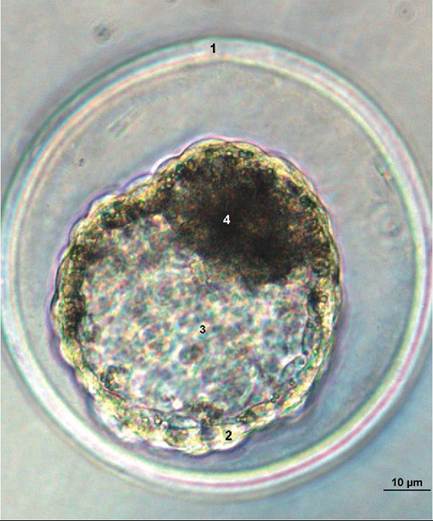
FIGURE 20-5 Pig blastocyst at day 6 of development, as seen through a microscope. 1, Zona pellucida; 2, Trophoblast (future placenta); 3, Blastocyst cavity; 4, Inner cell mass (future embryo). (From Hyttel P: Essentials of domestic animal embryology, St Louis, 2010, Saunders.)
exchanged across the very thin layers that separate the two sets of blood vessels.
Grossly, the placenta consists of layers of soft membranes that form two fluid-filled sacs around the developing fetus. The layer immediately around the fetus is called the amnion. It forms a sac around the fetus called the amniotic sac. The fetus floats in amniotic fluid inside this sac. Surrounding the amniotic sac is another layer called the allantois, which forms the fluid-filled allantoic sac. The outside of the allantoic sac is covered by the chorion, which attaches to the uterine lining. The chorion is linked to the fetus by the umbilical cord.
The umbilical cord is the link between the fetus and the nutrient and waste exchange structures of the placenta. As its name implies, it is a cordlike structure that contains blood vessels (the umbilical arteries and umbilical vein) and a drainage tube from the fetus' urinary bladder (the urachus). The two umbilical arteries carry unoxygenated, waste-filled blood from the fetus to the placenta. The single umbilical vein carries nutrient- and oxygen-rich blood back from the placenta to the fetus. The urachus is a tube that runs from the cranial tip of the fetus' urinary bladder through the umbilical cord to the allantoic sac. The kidneys of the developing fetus are not fully functional through most of the pregnancy, therefore they do not produce urine as we know it. They do, however, produce a watery fluid that must be removed from the urinary bladder. The urachus drains this fluid out into the allantoic sac. It normally closes at birth.
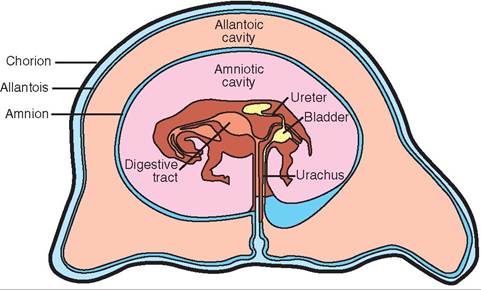
FIGURE 20-6 Fetus and placenta of a horse.
Fortunately, the placenta grows right along with the fetus and meets its needs every step of the way.
STRUCTURE
The placenta is a multilayered, fluid-filled, membranous sac that develops around the embryo and is connected to it by the umbilical cord (Figures 20-6 and 20-7). In some areas the outermost layer of the placenta attaches to the lining of the uterus. It is at these areas of attachment that the actual exchange of nutrients and wastes takes place between the fetal circulation and the maternal circulation. The fetal and maternal blood vessels are separate but in close proximity to each other in this area. Normally, no direct mixing of fetal and maternal blood occurs. The nutrients and wastes are
CLINICAL APPLICATION
The Afterbirth
At birth, the placenta is often referred to as the afterbirth because it is delivered after the offspring. What actually happens is that the fetus is delivered through the placenta. The powerful contractions of uterine and abdominal muscles during labor cause the membranes of the placenta to rupture and release their fluid. This is called the water breaking. Often the amnion still partially covers the newborn after it is delivered. The dam will usually lick this off, starting at the face so that the newborn can draw those important first breaths. If the mother does not perform this important duty, humans must intervene if they are present. The membrane is very soft and is easily broken and pulled away from the newborn's face. Few things in veterinary medicine are more rewarding than removing the membrane from the face of a newborn animal and watching it take its first breaths!
ATTACHMENT TO THE UTERUS
The area where the chorion attaches to the lining of the uterus is where the fetal and maternal blood vessels intertwine with each other. The exchange of nutrients and wastes between the fetal and maternal bloodstreams takes place here. The type of attachment varies among species but can be categorized into one of four general types: diffuse, cotyledonary, zonary, and discoid.
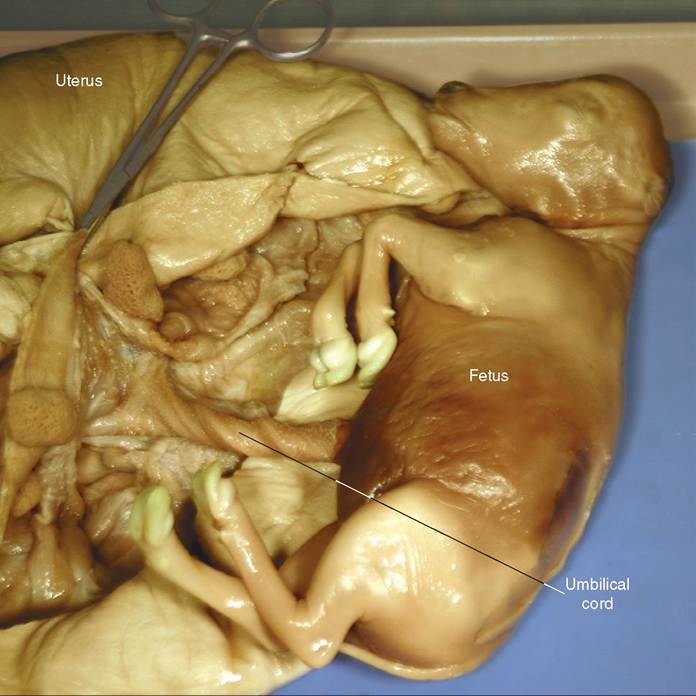
FIGURE 20-7 Umbilical cord in preserved pregnant cow uterus. The umbilical cord links the fetus with the placenta. It contains arteries that bring waste-filled blood from the fetus to the placenta, a vein that brings nutrient and oxygen-rich blood back to the fetus from the placenta, and the urachus, a tube that runs from the urinary bladder of the fetus to one of the placental sacs around the fetus (the allantoic sac). (From Colville T, Bassert J: Clinical anatomy and physiology laboratory manual for veterinary technicians, St Louis, 2009, Mosby.)
DIFFUSE PLACENTAL ATTACHMENT. Diffuse placental attachment means that the attachment sites are spread diffusely over the whole surface of the placenta and the whole lining of the uterus. There are no small, focal areas of attachment. Species with this type of placenta include pigs, horses, and camelids (Figure 20-8). Because the attachment sites are so diffuse and loosely attached, this type of placenta usually detaches easily from the uterine lining and is readily passed after the delivery of the newborn.
COTYLEDONARY PLACENTAL ATTACHMENT. Cotyledonary placental attachment is the most complicated type and is somewhat the opposite of the diffuse placental attachment. The areas of attachment are small, discrete, and numerous. Each of the many separate attachment sites is called a placentome. Each placentome consists of an area on the surface of the placenta called a cotyledon that joins with a mushroomlike caruncle in the lining of the uterus. The cotyledon and caruncle interdigitate tightly with each other. This type of placenta is found in ruminants, such as cattle, sheep, and goats. See Figure 20-9.

FIGURE 20-8 Diffuse placentation. Afterbirth from a Bactrian Camel. Like those of horses and swine, the attachment of the placenta of the camel to the lining of the uterus is diffuse. This afterbirth is about 6 feet long by about 12 to 18 inches wide. Note the relatively smooth surface. (Photograph by the author and used with permission from the Red River Zoo, Fargo, ND; www.redriverzoo.org)
For a cotyledonary placenta to pass after the birth of the offspring, each of the individual placentomes must separate completely. Sometimes this does not occur, and part or all of the placenta is retained in the uterus after the birth process. This retained portion of placenta dies and
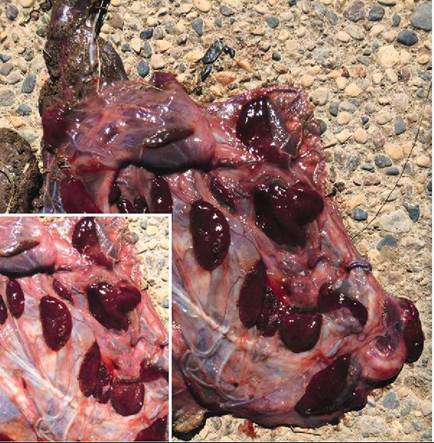
FIGURE 20-9 Cotyledonary placentation. Afterbirth from an African Pygmy Goat with detail. Note the dark cotyledons, the former attachment sites of the placenta with the caruncles in the lining of the uterus during gestation. (Photograph by the author and used with permission from the Red River Zoo, Fargo, ND; www.redriverzoo.org)
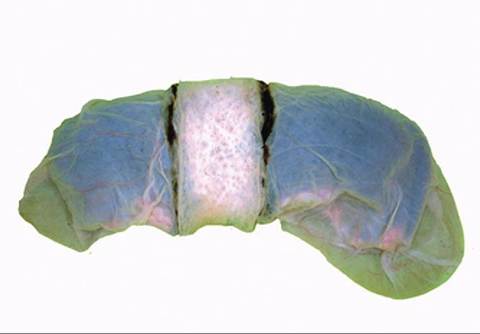
FIGURE 20-10 Zonary placentation. Note the pale area of attachment encircling the placenta like a belt. (From Dyce KM, Sack WO, Wensing CJG: Textbook of veterinary anatomy, ed 4, St Louis, 2010, Saunders.)
degenerates within the uterus, creating a potentially dangerous situation for the already stressed dam. It can lead to a serious postpartum (after birth) metritis (infection of the uterus) because bacteria can live and multiply in the dead placental tissue. Ensuring that the placenta is delivered by ruminant animals is important, and appropriate treatment must be given if it is not.
ZONARY PLACENTAL ATTACHMENT. With zonary placental attachment, the placenta attaches to the uterus in a belt-shaped zone that encircles the placenta. See Figure 20-10. Found in dogs and cats, this type of attachment also detaches fairly easily after delivery of the newborn. Retained placentas are not common in dogs and cats.
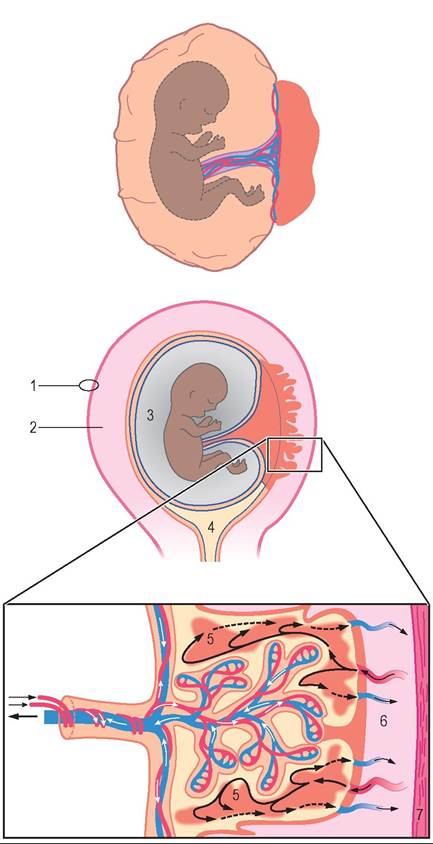
FIGURE 20-1 1 The discoid primate placenta. (From Hyttel P: Essentials of domestic animal embryology, St Louis, 2010, Saunders.)
DISCOID PLACENTAL ATTACHMENT. The pattern of discoid placental attachment is fairly self-descriptive. The area of attachment between the placenta and uterus is a single, discrete, disc-shaped area. This type of attachment is found in humans and other primates, as well as in rabbits and many rodents. See Figure 20-11.
/ TEST YOURSELF 20-2
1. Why is the placenta so important to a successful pregnancy?
2. Describe the relationship between the fetus and the amniotic and allantoic sacs of the placenta.
3. Describe the main structures that make up the umbilical cord and the function of each.
4. Which type of placental attachment to the uterus is the simplest and detaches most easily after parturition? Which is most complicated and most often results in retention of the placenta?
GESTATION
The period of pregnancy is called gestation, or the gestation period. It is the time from fertilization of the ovum to delivery of the newborn. It is convenient to divide it into three, often unequal, segments, called trimesters. The first trimester is the period of the embryo, when the newly implanted zygote is getting itself organized and developing its lifesupporting placenta. During this period, the developing offspring is often referred to as an embryo. Starting with the second trimester, the developing offspring is usually called a fetus. The second trimester is the fetal development period, when all the various parts of the fetus are taking shape and differentiating from each other. All the body tissues, organs, and systems develop during this period. The third trimester is the period of fetal growth. All parts of the fetus grow dramatically during this last period of development, preparing it to transition from a parasitic to a free-living existence after birth. The lengths of the gestation period in some common species are listed in Table 20-2.
PARTURITION
The whole, grand purpose of the male and female reproductive systems is fulfilled when parturition, the birth process, successfully takes place. At birth the fetus, which has led a parasitic existence up until now, is pushed out of the warm, dark, moist environment of the uterus into the cold, bright, outside world as a free-living, independent animal. In a very short time the body of the fetus has to undergo some dramatic changes. Its lungs, which have been nonfunctional until now, must suddenly expand with air and immediately start supplying the newborn with the oxygen it needs and eliminating the carbon dioxide its cells are producing as a waste product. Contributing to this process are some
| TABLE 20-2 | Gestation Periods of Selected Species | |
| SPECIES | RANGE | APPROXIMATE GESTATION PERIOD |
| Alpacas | 335-345 days | 11 months |
| Camels | 360-420 days | 13 months |
| Cats | 56-69 days | 2 months |
| Cattle | 271-291 days | 9 months |
| Dogs | 59-68 days | 2 months |
| Elephants | 615-650 days | 21 months |
| Ferrets | 42 days | 6 weeks |
| Goats | 146-155 days | 5 months |
| Hamsters | 19-20 days | 3 weeks |
| Horses | 321-346 days | 11 months |
| Humans | 280 days | 9 months |
| Llamas | 330-360 days | 11.5 months |
| Pigs | 110-116 days | 3 months, 3 weeks, and 3 days |
| Porcupines | 205-217 days | 7 months |
| Rabbits | 30-32 days | 1 month |
| Sheep | 143-151 days | 5 months |
dramatic changes in blood flow in and around the heart and lungs. The foramen ovale and ductus arteriosus, which shunted most of the blood from the right side of the fetal heart around, rather than through, the lungs, must both close fairly quickly. (See Chapter 14 for a complete description of these two fetal structures.) The lungs now need their full blood supply to oxygenate the blood properly and eliminate carbon dioxide.
Precisely what triggers parturition is not known. Many factors seem to be involved, including the size and weight of the uterus and fetus, and changing hormone levels in the fetus and the dam. The hormonal changes form a chain of events that leads to the onset of labor. Late in pregnancy the placenta of some species produces the hormone relaxin which helps relax ligaments between the bones around the birth canal, to ease the passage of the newborn. When it is time for parturition to get started, things start happening quickly, both in mom and in the fetus. The progesterone level in the bloodstream of the dam plummets (decreases rapidly) owing to the luteolytic effect of prostaglandin F2alpha. This causes the corpus luteum of the ovary to regress quickly and stop producing progesterone. Up until now, the high progesterone level from the corpus luteum, and other sources, has kept the myometrium (muscle of the uterus) quiet, preventing it from prematurely expelling the fetus. The luteolytic prostaglandin F2alpha is released from the placenta and the uterine wall as a result of stimulation from increased levels of glucocorticoid hormones from the adrenal glands of the fetus. The prostaglandin also causes a rise in the dam's estrogen level. The combined estrogen and prostaglandin levels increase the myometrium's sensitivity to oxytocin, which is released from the dam's posterior pituitary gland. Oxytocin stimulates the myometrium to contract, and this starts the labor process.
LABOR
The three distinct stages of parturition are referred to as the three stages of labor.
The first stage of labor consists of uterine contractions. The myometrium (muscle layer of the uterus) contracts as the cervix relaxes, pressing the membrane-covered fetus down against the cervix. This causes the cervix gradually to dilate. Externally, the dam appears restless and uncomfortable as a result of these uterine contractions. She may repeatedly lie down and get up and may urinate frequently. Some species, such as dogs and pigs, may attempt to build a nest into which they will deliver their young.
The second stage of labor consists of the actual delivery of the newborn. This is accomplished by a combination of strong uterine and abdominal muscle contractions. The dam typically lies down and strains in a rhythmic pattern of contractions that gradually become stronger and closer together. Rupture of the “water bags,” the amniotic and allantoic sacs of the placenta, usually precedes the actual delivery of the newborn.
The third stage of labor consists of delivery of the placenta (afterbirth). The placenta separates from the wall of the uterus and is expelled by weaker uterine contractions. The dam often eats the placenta. See Figure 20-12.
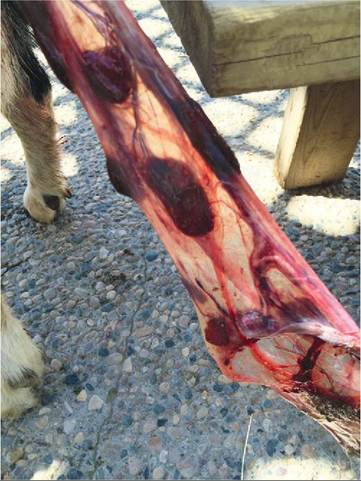
FIGURE 20-1 2 Afterbirth of an African Pygmy Goat during the third stage of labor. The goat is off the figure to the left. Mild traction is being applied to the placenta to aid its passage. (Photograph by the author and used with permission from the Red River Zoo, Fargo, ND; www.redriverzoo.org)
CLINICAL APPLICATION
Dystocia
Usually parturition comes off without a hitch. After all, animals have been giving birth for millions of years without any help. However, sometimes problems develop that interfere with the birth process, causing dystocia, or a difficult birth. The most common causes of dystocia include a fetus that is too large for the dam to pass and a fetus that is in the wrong position for delivery, which is called an abnormal presentation. Abnormal presentations may involve deviations in the position of the head, one or more legs, or the overall orientation of the fetus from the normal snout-first or rear- legs-first position. Sometimes the fetus can be pushed back far enough to allow it to be repositioned for delivery; this is called repelling the fetus. At times this is not possible, and the fetus must be removed surgically by an operation called a cesarean section, or C-section. In some cases, particularly in cattle, when the fetus is dead, it may have to be cut up (called an embryotomy) into small enough segments to be removed through the birth canal to save the life of the dam.
In multiparous species, such as dogs, cats, and pigs, that normally give birth to multiple offspring, the second and third stages of parturition intermix with one another. Typically newborns and placentas are delivered alternately; that is, after a newborn is delivered, its placenta is usually expelled before the next newborn is delivered.
Table 20-3 lists the names for offspring of selected species.
INVOLUTION OF THE UTERUS
After parturition is complete, the uterus immediately begins contracting down to its nonpregnant size through a process
| TABLE 20-3 | Offspring Names of Selected Species | ||
| SPECIES | OFFSPRING | SPECIES | OFFSPRING |
| Alpaca | Cria | Hedgehog | Pup |
| Ass/Donkey | Foal | Horse | Foal |
| Camel | Calf | Llama | Cria |
| Cat | Kitten | Monkey | Infant |
| Cattle | Calf | Mouse | Pup |
| Chicken | Chick | Mule | Foal |
| Cockroach | Nymph | Octopus | Fry |
| Deer | Fawn | Otter | Pup |
| Dog | Puppy | Parrot | Chick |
| Elk | Calf | Pig | Piglet |
| Ferret | Kit | Rabbit | Kit |
| Fly | Maggot | Rat | Pup |
| Gerbil | Pup | Sheep | Lamb |
| Goat | Kid | Snake | Hatchling |
| Goose | Gosling | Swan | Cygnet |
| Guinea Pig | Pup | Wolf | Pup |
| Hamster | Pup | Worm | Wormlet |
called involution of the uterus. At the placental attachment site(s), the endometrium sloughs (dies and detaches) into the lumen of the uterus, and the exposed areas heal over. The myometrium slowly continues to contract, squeezing the contents of the uterus out through the birth canal as the organ returns to its nonpregnant size. Initially the discharge contains fresh blood resulting from separation of the placenta from its attachment site(s). Pressure from the continued uterine contractions usually stops the bleeding relatively quickly. The discharge gradually turns darker as dead tissue is liquefied and sloughed from the uterine wall. It generally takes from a few weeks to a month or more for involution to be complete.
TEST YOURSELF 20-3
1. What are the basic events of the three trimesters of pregnancy?
2. Describe the three stages of labor.
3. Why is it important that uterine contractions continue after the fetus and placenta have been delivered?
MAMMARY GLANDS AND LACTATION
Once parturition is completed, the newborn animal must be nourished and cared for during the neonatal period. The mammary glands are very important now.
CHARACTERISTICS
The mammary (milk) glands are specialized skin glands. They produce colostrum and milk, which are needed by the newly born animal during the crucial first few hours, days, and weeks of life. Although, strictly speaking, they are not part of the reproductive process, their function is vital to the survival of the newborn, therefore we discuss them along with pregnancy.
Mammary glands are present in both male and female animals, although they can be difficult or impossible to find in stallions. They normally only function in females because males do not secrete the proper blend of hormones to make them work. (It is theoretically possible to make a bull give milk with administration of the proper combination of hormones, but who would want to milk him?)
SPECIES DIFFERENCES
Mammary glands look very different among common species of animals. Their number varies, from a low of two in horses to a high of 14 in swine, and their locations range from the inguinal (groin) region alone in cattle and horses, to locations that span the inguinal, abdominal, and thoracic regions in dogs, cats, and swine. Also, considerable variety is found in the number of openings from which milk emerges in the teats or nipples. Cattle, sheep, and goats only have one opening per teat, whereas dogs can have up to 20 openings per nipple. (Humans have up to 24 openings per nipple.) The mammary gland characteristics of some common species are listed in Table 20-4.
UDDER OF THE COW
Because they are so large and specialized, a cow's mammary glands, commonly known as the udder, seem like exaggerated caricatures of other species' mammary glands. Their general makeup, however, is just a larger version of other animals' mammary glands. For this reason, we use them as our model mammary system (Figure 20-13).
The udder of the cow consists of four mammary glands that are called quarters. Each quarter is a completely separate unit from the other three. They each have their own milk-secreting systems and ducts leading down to their own teats. This has important clinical significance if infection of the mammary gland, a condition called mastitis, develops.
| TABLE 20-4 | Mammary Gland Characteristics of Some Common Species | ||
| USUAL NUMBER SPECIES OF GLANDS | LOCATION OF GLANDS | NUMBER OF OPENINGS IN TEATS OR NIPPLES | |
| Cats | 10 | Inguinal, abdominal, and thoracic regions | 3-7 |
| Cattle | 4 | Inguinal | 1 |
| Dogs | 10 | Inguinal, abdominal, and thoracic regions | 8-20 |
| Goats | 2 | Inguinal region | 1 |
| Horses | 2 | Inguinal region | 2-4 |
| Humans | 2 | Thoracic region | 15-24 |
| Pigs | 14 | Inguinal, abdominal, and thoracic regions | 2-3 |
| Sheep | 2 | Inguinal region | 1 |
Infection does not spread directly from one quarter to another. It has to spread down through the teat and duct system of one quarter and up another, or on rare occasions, it can spread systemically (through the bloodstream). The spread of mastitis in a herd can be prevented through good milking hygiene, which includes milking infected cows last and keeping facilities clean.
The udder of a high-producing dairy cow can weigh more than 100 lb at milking time. It needs a strong suspensory system to support the heavy weight and attach the udder to the body wall. This suspensory system consists of a slinglike arrangement of ligaments that run down the center and around the sides of the udder. The median suspensory ligament contains many elastic fibers so that it can stretch. It passes down the center (along the median plane) between the left and right halves of the udder. The lateral suspensory ligaments are composed largely of strong but relatively inelastic collagen fibers. They pass down and around the lateral sides of each half of the udder. The strong lateral ligaments provide firm support for the udder, and the elastic medial ligament acts as a shock absorber for the udder as the animal moves around. Note: When the udder is empty, the teats point pretty much straight down. When the udder is full of milk, however, the teats angle off to the left and right sides a bit. This is due to stretching of the elastic median ligament that runs down the center of the udder.
ALVEOLI AND DUCT SYSTEM
The milk-secreting units of the mammary gland are small structures called alveoli. Each alveolus is a tiny, saclike arrangement of cells that secretes milk into an equally tiny tube called the alveolar duct. The alveoli are arranged like clusters of grapes around the alveolar ducts. The tiny alveolar ducts join to form larger ducts, which join to form even larger ducts, and so on. The duct system of the mammary gland is similar to the arrangement of branches in a tree. The clusters of alveoli would be similar to the leaves, and the duct system would be like the small tree branches joining to form larger branches. The largest ducts (similar to the trunks of trees) empty into a large space called the gland sinus, located just above (dorsal to) the teat. The gland sinus is continuous with the teat sinus inside the teat. The two large sinuses form a space shaped like an upside-down pear. They are the spaces where milk accumulates when milk let-down (described later) has occurred. The milk can be readily extracted from these large spaces by the suckling of a nursing newborn or the gentle vacuum of a milking machine.
At the tip of the teat is the streak canal, which is the passageway from the teat sinus to the outside. It is surrounded by elastic fibers and a ringlike sphincter muscle that keeps it closed most of the time. This helps minimize milk leakage.
MAMMARY GLAND DEVELOPMENT
The mammary glands stay small and undeveloped until puberty (the time of sexual maturity). Up until that time,
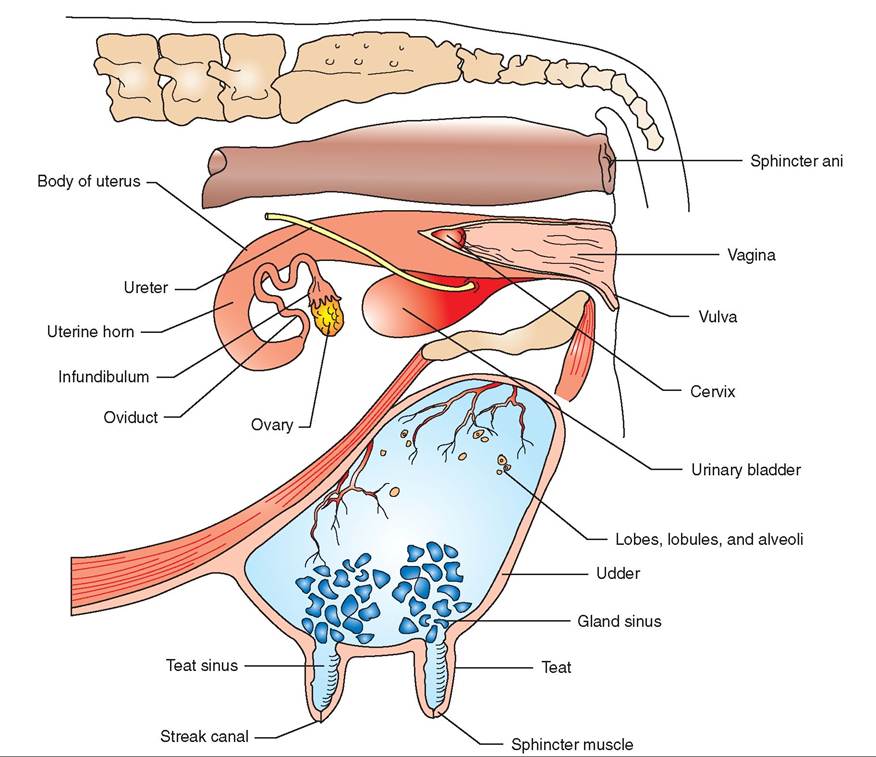
FIGURE 20-13 Reproductive system and udder of cow. (Modified from McBride DF: Learning veterinary terminology, ed 2, St Louis, 2002, Mosby.)
the proper mix of hormones has not occurred to stimulate their growth and development. Once estrous cycles begin at puberty, however, the mammary glands respond to the new hormones flowing through the body by enlarging and gearing up to produce milk.
A complex balance of hormones stimulates the mammary olands to develop. Most of the anterior pituitary hormones are involved in the process either directly or indirectly. Prolactin and growth hormone directly encourage mammary gland denelopment. Follicle-stimulating hormone (FSH) and luteinizing hormone (LH) stimulate the ovaries to rporgoednuce est and progesterone during each heat cycle. The estrogen and progesterone encourage the alveoli and duct s^ms of the mammary glands to develop. Thyroidstimulating hormone and adrenocorticotropic hormone (ACTH) influence the process indirectly through their target organs (the thyroid gland and the adrenal cortex). The levels aorfiothuesse v hormones must be balanced precisely for complete mammary gland development to take place. Abnormally high levels of any hormones, such as might hocecnur w thyroid hormone or corticosteroid drugs are administered to a young animal, can actually inhibit normal lmanadmmary g development.
LACTATION
hTyhseicapl growth and development of the mammary glands are the first steps in making them able to produce ehmsetilk. T r takes place during pregnancy.
Lactation is the process of milk production. It begins toward the end of pregnancy and is obvious at the time of parturition. Prolactin and growth hormone from the ante- ruiiotarrpyit gland and hormones from the adrenal cortex avroelviend in getting lactation started.
COLOSTRUM
Before it starts to produce milk, the mammary gland produces a sort of premilk secretion called colostrum. Colostrum has a different appearance and a different composition trhmaanl no milk. It contains larger amounts of proteins, lipids, and amino acids than milk and also contains high levels of various essential vitamins. It supplies important onutrients t the newborn and has a laxative effect that helps clear the dark, sticky meconium from the newborn's intestinal tract.
Probably the most critical of colostrum's roles is the transfer of what is called passive immunity from the dam to the newborn. Among the proteins in colostrum are high levels of immunoglobulins, also called antibodies, that are an important part of the body's defense against infection. The antibodies in the colostrum are specific for disease- causinm organisms that the dam has been exposed to or vaccinated against. If the newborn drinks sufficient colostrum ⅛'ng the first few hours after birth, the large antibody imllolecules w be absorbed intact into its bloodstream. This provides it with important passive immunity, in which com- rpelfeoter,mped antibodies help protect it against diseasecausing microorganisms until its own immune system umfafitcuierenstlys to protect it. If colostrum is not consumed within the first few hours, the lining of the newborn's intestine can no longer absorb the large antibody molecules ihnetyact. T will be broken down by the digestive process, and passive immunity will not be transmitted. Youngsters tohat d not get sufficient colostrum in that critical early neonatal period are much more prone to diseases. Even if tohey d not die of early infections, they are typically weaker aond d not grow as rapidly as animals that consumed colostrum at the appropriate time.
CLINICAL APPLICATION
Passive Immunity and Vaccinations
The passive immunity that newborns acquire from drinking colostrum is important to their early survival. However, it ean interfere with early vaccinations, particularly in puppies and kittens. If a puppy or kitten is vaccinated while it still has a high level of antibodies from its mother in its bloodstream, the vaccine will be inactivated and will not stimulate the animal's immune system to manufacture new antibodies. So why not just wait until the passive immunity from mom wears off? He difficulty is that passive immunity lasts dif- fnegrtihnsg le of time in different animals. If you vaccinate ltyo,o ear the vaccine is wasted. If you wait too long to vaccinate, the passive immunity may have worn off, leaving the animal susceptible to infection. The level of passive antibod- ioeusning the y animal's bloodstream can be tested, but this is expensive. The most practical solution is to give the young raineismal a se of vaccinations spread out over the period when protection from passive immunity is most likely to end. Specific vaccination protocols differ, but the first vaccination is usually given somewhere around 6 to 8 weeks of age, with subsequent vaccinations given every 2 to 3 weeks until the animal is 14 to 16 weeks old.
MAINTENANCE OF LACTATION
Once it has begun, lactation will continue as long as the lmanadmmary g is emptied regularly by nursing or milking. The key is continued physical stimulation of the teat or nipple, combined with regular removal of milk from the gland. These activities send sensory nerve impulses to the brain. From there nerve pathways lead to the hypothalamus, which stimulates the anterior pituitary gland to continue its production o0 the hormones that keep lactation going. When nursing or milking stops, the flow of essential hormones stops also. The lframcko noalho stimulation combined with increased pres
sure in the gland (because it is no longer being emptied) causes lactation gradually to cease. The mammary gland “dries up.” This is called involution of the mammary gland.
MILK LET-DOWN
Milk let-down is the immediate effect of nursing or milking. When milk is produced, it accumulates in the outer reaches of the mammary gland, in the alveoli and small ducts. It does vneot mo into the larger ducts and sinuses, where it is accessible for nursing or milking, until milk let-down occurs. uWrshienng n or milking begins, sensory nerve impulses are sroeanint.t the b As we've seen, this causes the hypothalamus itmo sutlate the anterior pituitary gland to produce the hor
mones necessary to maintain lactation. However, it also has another major effect. It also causes the hypothalamus to release the hormone oxytocin from the posterior pituitary ghleand. T oxytocin travels to the mammary gland and cuasuclseelsikme myoepithelial cellsoaurnd the alveoli and small ducts to contract. This squeezes milk down into the luacrtgse d and sinuses, where it can be removed by nursing or milking. The process of milk let-down takes from a few soeconds t a minute or more to produce results, so there is hoftten a slig delay from the time a newborn starts to nurse thoe nw the milk starts to flow freely.
TEST YOURSELF 20-4
1. Why does mastitis inone quarter of adairy cow's udder mot necessarily spread to the other three quarters?
2. Describe the suspensory apparatus of the udder.
d. Why don'ttha Siamrnury glands of male animals usually develop or secrete milk?
4. Describe the importatae of colostrum to the health of a mewborm animal.
5. Describe how nursing or milking causes milk let-dowm amd also helps sustaim lactatiom.