The Reproductive System
Thomas Colville
OUTLINE
INTRODUCTION, 465
MEIOSIS, 466
Chromosomes, 466
Spermatogenesis, 468
Oogenesis, 469
MALE REPRODUCTIVE SYSTEM, 469
Testes, 469
Vas Deferens, 474
Urethra, 474
Accessory Reproductive Glands, 474
Penis, 475
FEMALE REPRODUCTIVE SYSTEM, 477
Ligaments, 477
Ovaries, 478
Oviducts, 482
Uterus, 482
Cervix, 482
Vagina, 483
Vulva, 483
The Estrous Cycle, 483
LEARNING OBJECTIVES
When you have completed this chapter you will be able to:
1.
Differentiate between meiosis and mitosis.2. Describe the process of spermatogenesis.
3. Describe the process of oogenesis.
4. Describe the structure of spermatozoa.
5. List the components of the male reproductive system and describe the functions of each.
6. List the characteristics and functions of the accessory structures of the male reproductive system.
VOCABULARY FUNDAMENTALS
Acrosome ahck-rδ-sδm
Afterbirth ahf-tar-barth
Ampulla ahm-pyool-uh
Androgen ahn-drδ-jehn
Anestrus ahn-ehs-truhs
Antrum ahn-truhm
Body of the penis boh-de of the pe-nihs
Broad ligament brawd lihg-ah-mehnt
Bulb of the glans buhlb of the glahnz
Bulbourethral gland buhl-bδ-u-re-thruhl glahnd
Cervix sar-vihx
Chromosome krδ-muh-sδm
Clitoris klih-tohr-ihs
Common vaginal tunic kohm-uhn vahj-ihn-ahl too-nihck
Cornification kohr-nih-fih-ka-shuhn
Corona glandis kuh-rδ-nuh glahn-dihs
Corona radiata kuh-ro-nuh ad-e-aht-ah
7. List the components of the female reproductive system and describe the functions of each.
8. Describe the events that occur during the ovarian cycle.
9. List the stages of the estrous cycle and describe the events that occur during each stage.
10. Differentiate among monoestrous, diestrous, polyestrous, and seasonally polyestrous cycles.
Corpus cavernosum penis kohr-puhs kahv-ar-nδ-suhm pe-nihs
Corpus cavernosum urethrae kohr-puhs kahv-ar-nδ- suhm u-re-thre
Corpus hemorrhagicum kohr-puhs hehm-uh-rahj -uh-kuhm
Corpus luteum kohr-puhs lu-te-uhm
Cremaster muscle kre-mahs-tar muhs-uhl
Crura krar-uh
Cryptorchidism krihpt-ohr-kihd-ihsm
Cumulus oophorus kyoo-myuh-luhs δ-δ-far-uhs
Diestrous dι-ehs-truhs
Diestrus dι-ehs-truhs
Diploid chromosome number dihp-loyd krδ-muh-sδm nuhm-bar
Efferent ducts of the testes e-far-ehnt duhcktz of the tehs-tes
Ejaculation eh-j ahck-u-la-shuhn
Epididymis ehp-ih-dihd-eh-mihs
Erectile tissue eh-rehck-tihl tihsh-yoo
Erection eh-rehck-shuhn
Estrogen ehs-trδ-jehn
Estrous ehs-truhs
Estrous cycle ehs-truhs sι-kuhl
Estrus ehs-truhs
Fimbriae fihm-bre-e
Follicle fohl-ih-kuhl
Follicular atresia fuh-lihck-yuh-lar a-tre-zhuh
Follicular cell fuh-lihck-yuh-lar sehl
Fossa glandis fohs-ah glahn-dihs
Gamete gahm-et
Glans of the penis glahnz of the pe-nihs
Glycoprotein glι-kδ-prδ-ten
Gonad gδ-nahd
eGlrl anulosa c grahn-u-lδ-hslah se
Gubernaculum goo-bar-nahck-yuh-luhm
Haploid chromosome number hahp-loyd krδ-muh-sδm
nuhm-bar
Homologous hδ-mohl-δ-guhs
Hydraulic pressure hι-draw-lihck prehsh-ar
Infundibulum ihn-fuhn-dihb-yuh-luhm
Inguinal ring ihn-gwih-nahl rihng
Interstitial cell ihn-tar-stihsh-ahl sehl
Labia la-be-uh
Ligate lι-gat
Mature f^‰Ze maht-char fohl-ih-kuhl
Meiosis mι-δ-sihs
Metestrus meht-ehs-truhs
Mitosis mι-tδ-sihs
Monoestrous mohn-δ-ehs-truhs
Multiparous muhl-tihp-ear-uhs
Oocyte δ-δ-sιt
Oogenesis δ-δ-jehn-eh-sihs
Orchiectomy ohr-kihd-ehck-tδ-me
Os pen is ohs pe-nihs
Oviarian follicle δ-vear-e-uhn fohl-ih-kuhl
Ovary δ-var-e
Oviduct δ-vih-duhckt
Ovulation ohv-yoo-la-shuhn
Ovum δ-vuhm
Pampiniform plexus pahm-pihn-eh-fohrm plehck-suhs
Parietal vaginal tunic pah-rι-eh-tahl vahj-ihn-ahl too-nihck
Penis pe-nihs
Placenta plah-sehn-tah
dPyo lar bo pδ-larohb-d e
Polyestrous pohl-e-ehs-truhs
Prepuce pre-pyoos
Proestrus prδ-ehs-truhs
Progestin prδ-jehs-tihn
Proper vaginal tunic praw-par vahj-ihn-ahl too-nihck
Prostate^ιd prah-stat gladnd
Pseudocyesis soo-dδ-sι-e-sihs
Pseudopregnancy soo-dδ-prehg-nuhn-se
Retractor penis muscle rih-trahck-tar pe-nihs muhs-uhl Roots of the penis rootz of the pe-nihs
Round ligament of the uterus round lihg-ah-mehnt of the u-tar-uhs
Scrotum skrδ-tuhm
Seasonally polyestrous se-zuhn-ahl-e pohl-e-ehs-truhs eSseimclien al v sehm-ihn-uhl vehs-ih-kuhl Seminiferous tubule sehm-uh-nihf-ar-uhs too-bul Sertoli cell sar-tδ-le sehl
Sex chsomosome sehx krδ-muh-sδm
Xigmoidflrnre sihg-moyd flehck-shar
Spermatic cord spar-maht-ihck kohrd
Spermatogenesis spar-mah-tδ-jehn-eh-sihs Spermatozoon spar-maht-uh-zδ-uhn
Sphincter sfihnk-tar
Suspensory ligament of the ovary suh-spehn-suh-re lihg -ah-mehnt of the δ-var-e
Testes tehs-tes
Tie tι
Uniparous u-nihp-ear-uhs
Urethra u-re-thrah
Urethral process u-re-thruhl proh-sehs
Uterus u-tar-uhs
Vagina vah-jι-nah
Vaginal tonic vahj-ihn-ahl too-nihck
Vae deferens vahs dehf-ar-ehns
Vestibule of the vulva vehs-tuh-byool of the vuhl-vah Visceral vaginal tunic vih-sar-ahl vahj-ihn-ahl too-nihck Vulva vuhl-vah
INTRODUCTION
To say the least, the reproductive system is unusual.
The rest of the body's systems work to ensure the survival of the individual animal they are part of, whereas the reproductive system works to ensure the survival of the species of animal. The reproductive system does interact with other body systems, but purely reproductive structures are not essential to the life of an animal. Think about it: The testes, ovaries, and other reproductive organs of male and female animals are commonly removed surgically to prevent reproduction and influence behavior. If no complications result from the surgery, the rest of the body systems continue to go merrily on their way, and the animal is none the worse for its loss. It simply cannot reproduce. Of course it is a different story with structures that the reproductive system shares with other systems. For example, the urethra in the male has both urinary and reproductive functions. It is essential to life and must not be damaged or removed.Something else unique about the reproductive system is that it requires a second animal (of the opposite sex) to fully carry out its function, which is to produce a brand new animal (the offspring). What we refer to as the reproductive system in an individual animal is really only half a system. A complete reproductive system is made up of all the male reproductive organs and structures in one animal and all the female reproductive organs and structures in another. Both are necessary for an offspring to be produced.
The basic reproductive process starts with fertilization: the head of a spermatozoon must penetrate into the cytoplasm of an ovum. The fertilized ovum must then be provided with a hospitable place to grow and develop until the offspring is born. After birth, the offspring must be fed and cared for until it can fend for itself. For these basic processes to occur, the physical characteristics and behavior of two different animals must be synchronized and coordinated with delicate precision. This elegant process of developing and bringing together a male animal and a female animal at the appropriate time and in appropriate states of readiness to produce a viable offspring is the amazing role of the reproductive system.
For simplicity's sake, we discuss the male and female reproductive systems as separate physical entities. Remember, though, that a whole reproductive system is made up of both the male and female systems working in concert with each other.
/ TEST YOURSELF 19-1
1. How does the reproductive system differ from other body systems?
2. Why is the reproductive system of an individual animal considered only half a system?
MEIOSIS
Our discussion of reproduction begins at the most basic, cellular level. Gametes (reproductive cells—ova and spermatozoa) are produced by the unique process of cell division called meiosis. This is the first step in the process by which nature “shuffles the genetic cards” each time a new animal is conceived. Meiosis ensures that the genetic makeup of each new animal is unique. Each animal that enters the world has characteristics similar to others of its species and breed, but its precise genetic makeup is unique. Except for identical twins or clones, it is virtually impossible for two animals to have exactly the same genetic makeup.
CLINICAL APPLICATION
Unique Patients
The fact that each animal has an individual, unique genetic composition is not just hypothetical or interesting in an abstract sense. This uniqueness is an important concept for us to keep in mind as we examine and treat patients. It can significantly affect things such as behavior, clinical signs of disease, doses of drugs, and response to treatments. Even though all Golden Retrievers look a lot alike, a particular Golden Retriever we are working with may respond differently to our drugs and treatments than others of its breed we have worked with before. This is an important concept to grasp if we are to be effective in our work in clinical veterinary medicine. Each of our patients is an absolutely unique, original individual. It is reasonable to expect more similarities than differences among animals of a particular species and breed, but always keep in the back of your mind the possibility that this animal may not react exactly the same as others you have worked with in the past.
Chromosomes
DIPLOID CHROMOSOME NUMBER
The genetic material in the body's cells is contained in the nuclei in the form of coiled masses of DNA called chromosomes. This genetic material contains a complete “blueprint” for all of the structures and functions in the animal's body. For the body to function efficiently, each cell must contain the same genetic information. So each of the cells in an animal's body, with the exception of the gametes (spermatozoa or ova), contains identical chromosomes.
Another member of the same species has a slightly different genetic code in its chromosomes (the cause of that uniqueness business), but the total number of chromosomes in the nucleus of each of its body cells (except for the gametes) is the same. The total number of chromosomes in the nucleus of each cell is called the diploid chromosome number, which is always an even number, because the chromosomes occur in pairs (di means “double,” and ploid refers to chromosomes). Therefore all cattle have the same number of chromosomes in the nuclei of their cells. The same goes for all dogs, horses, humans, and even corn and tomato plants. Table 19-1 contains the diploid chromosome numbers of some selected species. The diploid chromosome number is sometimes expressed generically with the abbreviation 2n. The n is a mathematical expression that represents a number, and the 2 indicates that the number is doubled (diploid).
SEX CHROMOSOMES
One of the pairs of chromosomes that make up the diploid chromosome number are the sex chromosomes. They determine the genetic gender (male or female) of the animal. They are designated as being either X chromosomes or Y chromosomes. If both of the sex chromosomes are X chromosomes (XX), the individual is genetically female. If one is an X and the other is a Y (XY), the individual is genetically male. The YY combination is not possible, for reasons that we will discuss. Many other factors, including the influence of hormones, help determine the overall maleness or femaleness of an individual, but the makeup of the sex chromosomes is a basic determining factor.
(Table 19-2 lists common female and male gender terms.) So the full diploid chromosome makeup of a female animal is often abbreviated as 2n,XX and that of a male animal as 2n,XY.| TABLE 19-1 | Diploid Chromosome Numbers of Some Selected Species |
| SPECIES | DIPLOID CHROMOSOME NUMBER |
| African Hedgehog 90 | |
| Cat | 38 |
| Cattle | 60 |
| Corn | 20 |
| Dog | 78 |
| Donkey | 62 |
| Fox | 34 |
| Goat | 60 |
| Gray Fox | 66 |
| Horse | 64 |
| Human | 46 |
| Mosquito | 6 |
| Mouse | 40 |
| Pig | 38 |
| Rabbit | 44 |
| Rat | 42 |
| Red Panda | 36 |
| Sheep | 54 |
| Tomato | 24 |
HAPLOID CHROMOSOME NUMBER
Some basic mathematics reveals why the gametes cannot have the diploid number of chromosomes. If the ovum of a goat and the spermatozoon that fertilizes it each contained the full caprine diploid chromosome number of 60, the fertilized ovum would contain a total of 120 chromosomes! That is way too many. The number of chromosomes in the reproductive cells must be reduced to half of the diploid number for things to work out, so when the spermatozoon avnudmthe o come together, the total number of chromosomes in the fertilized ovum equals the diploid number.
eTdhuecred number of chromosomes in the gametes is called the haploid chromosome number (hapl- means “single”) and is abbreviated n,X or n,Y, depending on which sex chromosome is present. The haploid chromosome number in the reproductive cells results from a “reduction division” of cells called meiosis. IMsi^is is referred to as a reduction division because as a cell divides by tlnis process, the tuomtablenr of chromosomes in each of the daughter cells is reduced to one half the number in the parent cell. One of feach pair o diploid chromosomes from the parent cell goes to each of the daughter cells. Each daughter cell then has the haploid chromosome number.
MEIOSIS versus mitosis
You reca11 that when most of the body's cells divide, they do it through a process called mitosis. eVhen a cell divides by mitosis, each of its chromosomes first produces a duplicate copy of itself. When the two daughter cells pull apart, half thhreo mc osomes go to one cell and half go to the other.
enEladl csh c e up with an identical, full diploid set of chro- hmeosomes. T genetic makeup of the two daughter cells is
| TABLE 19-2 | Selected Common Gender Terms | |
| SPECIES | FEMALE | MALE |
| Ass/Donkey | Jenny | Jack |
| Bison | Cow | Bull |
| Camel | Cow | Bull |
| Cat | Queen | Tom |
| Cattle | Cow | Bull |
| Chicken | Hen | Rooster |
| Deer | Doe | Buck |
| Dinosaur | Cow | Bull |
| Dog | Bitch | Dog |
| Dolphin | Cow | Bull |
| Elephant | Cow | Bull |
| Ferret | Jill | Hob |
| Fox | Vixen | Dog |
| Gerbil | Doe | Buck |
| Guinea Pig | Sow | Boar |
| Goat | Doe | Buck |
| Hamster | Doe | Buck |
| Hedgehog | Sow | Boar |
| Horse | Mare | Stallion |
| Jellyfish | Sow | Boar |
| Leopard | Leopardess | Leopard |
| Llama | Cow | Bull |
| Mouse | Doe | Buck |
| Mule | Molly | John |
| Otter | Sow | Boar |
| Parrot | Hen | Cock |
| Pig | Sow | Boar |
| Porcupine | Sow | Boar |
| Porpoise | Cow | Bull |
| Rabbit | Doe | Buck |
| Rat | Doe | Buck |
| Red Panda | Sow | Boar |
| Sheep | Ewe | Ram |
| Squid | Hen | Cock |
| Tiger | Tigress | Tiger |
| Turkey | Hen | Tom |
| Whale | Cow | Bull |
| Wolf | Bitch | Dog |
| Zebra | Mare | Stallion |
exactly the same, and identical to that of the parent cell. This ensures that the genetic information in all of the body's cells, eoxrcept f the gametes, stays exactly the same.
aWmheetnesthe g are produced by meiosis, the chromo- soomes d not produce duplicate copies of themselves before the daughter cells pull apart. Half of the total number hf chromoeomes (one from each diploid chromosome lpuadirin),ginc one sex chromosome, goes to each daughter cheicllh. W chromosomes go to which daughter cell is entirely. ^om. The chromosomes merely pull apart into their new cells. In this way, the genetic material of the reproductive cells gets “shuffled,” resulting in genetically unique offspring.
CLINICAL APPLICATION
Chromosome Combinations
If you want to do some interesting mathematical calculations, determine how many different combinations of chromosomes could result for different species by randomly assigning one of each pair of diploid chromosomes to each reproductive cell. Hint: Because the diploid chromosomes occur in pairs, you can start with the number 2 and multiply it times itself as many times as there are pairs of chromosomes. For example, a diploid chromosome number of 8 would have four pairs of chromosomes, so the number of possible haploid chromosome combinations would be 24, or 16 possible combinations. Try using that calculation for some of the diploid chromosome numbers listed in Table 19-1. (Divide the diploid chromosome number in half and multiply 2 by itself that many times.) You are going to get some very large numbers. Those large numbers represent the number of possible combinations of chromosomes that could occur in each spermatozoon or ovum from an animal of that species.
If you want to calculate the number of chromosome combinations possible in an offspring of that species, you need to take the number of possible chromosome combinations for a spermatozoon and multiply it by the number of possible chromosome combinations for an ovum: you are going to come up with some ginormous numbers.
In a human, for example, the diploid chromosome number is 46, so the number of possible combinations of chromosomes in each spermatozoon or ovum is 223, or 8,388,608. The number of possible combinations of chromosomes in a human infant would then be 223 ? 223, or 70,368,744,177,664. Barring an identical twin, the chance of your parents producing a brother or sister with the exact same chromosome combination as you is 1 in 70,368,744,177,664!
SPERMATOGENESIS
Spermatozoa, the male gametes, are produced in the seminiferous tubules of the testes through a process called spermatogenesis (genesis means “creation”). Spermatozoa are produced continuously and in very large numbers in an effort to ensure that one spermatozoon will successfully reach and fertilize the ovum when breeding occurs. So the process of spermatogenesis is designed to produce huge numbers of spermatozoa continuously (Figure 19-1).
Spermatogenesis begins at the periphery of the seminiferous tubule with a cell called a primary spermatocyte. This cell has the normal diploid chromosome number for the species. The chromosome complement of the cell is abbreviated as 2n,XY. Keep in mind that the XY refers to the sex chromosomes, which in a male are always an X and a Y. The primary spermatocyte divides by meiosis into two secondary spermatocytes, which are pushed inward toward the tubule lumen. These now have haploid chromosome numbers. One has an n,X chromosome complement, and the other has an n,Y complement. The two secondary spermatocytes divide by mitosis (so that the chromosome number does not get reduced further) into four spermatids. Two of the spermatids have n,X chromosome complements, and two have n,Y. As a result of the cell divisions that produced them, the spermatids are now located near the center of the tubule lumen. They do not undergo any further cell divisions, but they grow tails and undergo other physical changes that convert them to spermatozoa. When the spermatozoa are fully developed, they detach and are carried to an organ called the epididymis for storage before ejaculation. This is discussed in more detail below.
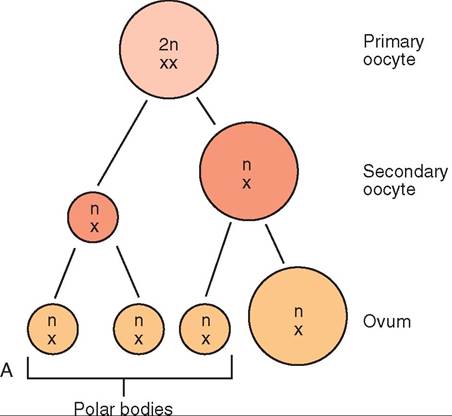
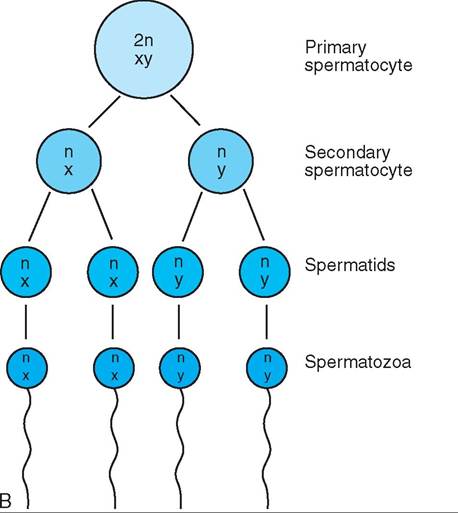
FIGURE 19-1 Meiosis. This process reduces the diploid chromosome number to the haploid number in reproductive cells. A, Oogenesis. Note that only one functional ovum is produced from the original primary oocyte. B, Spermatogenesis. Note that four spermatozoa are produced from the original primary spermatocyte.
Half of the spermatozoa produced have an X sex chromosome, and half have a Y sex chromosome. This is the means by which the genetic sex of the offspring will be determined. If an X-bearing spermatozoon fertilizes the ovum (which always contains an X sex chromosome), the offspring will have XX sex chromosomes and be genetically female. If a Y-bearing spermatozoon is successful, the offspring will have XY sex chromosomes and will be genetically male. This ensures sexual equality of numbers in that about the same number of females and males will be conceived.
OOGENESIS
Ova, the female gametes, are produced in follicles in the ovaries through a process called oogenesis. They are not produced continuously, unlike spermatozoa. At or soon after birth, a female has a fixed number of primary oocytes (which are the precursor cells to ova) already formed in her ovaries. That is the total number available in her lifetime. They remain in a quiet, immature state until they are recruited as part of an ovarian cycle. Each ovarian cycle produces one or more mature ova, depending on the species. Because spermatozoa come to the ovum to fertilize it, large numbers of ova are not needed. Therefore the process of oogenesis is designed to produce small numbers of ova at one time (see Figure 19-1).
Oogenesis takes place in the developing ovarian follicle. The primary oocyte is the immature cell that waits quietly in the ovary for the signal to start developing. It has the normal diploid chromosome number for the species. The chromosome complement of the cell is abbreviated as 2n,XX. When it becomes activated, the primary oocyte divides by meiosis into a large, secondary oocyte and a small polar body. Each has the haploid chromosome number n,X. Polar bodies are essentially garbage cans for excess chromosomes. They will not develop into ova. The secondary oocyte and the first polar body divide by mitosis (to preserve the haploid chromosome number) into an ovum and three polar bodies. So from one primary oocyte, one mature ovum will result.
MALE REPRODUCTIVE SYSTEM
The tasks for the male component of the reproductive system sound pretty simple: produce male sex hormones, develop
TEST YOURSELF 19-2
1. What is the difference between the diploid chromosome number and the haploid chromosome number? In which body cells is each found?
2. Which gamete, the ovum or the spermatozoon, determines the genetic sex of the offspring when fertilization occurs?
3. How does cell division by meiosis differ from cell division by mitosis?
4. How does spermatogenesis differ from oogenesis? Why are the basic processes so different?
male gametes (spermatozoa), and deliver the spermatozoa to the female system at the appropriate time. However, a rather complicated system of organs, glands, structures, and tubes is required to carry out these tasks. Figure 19-2 illustrates the reproductive system of the bull, and Figure 19-3 illustrates that of the dog. Let us start our examination of the male reproductive system with the basic organs of male reproduction: the testes.
TESTES
CHARACTERISTICS
The testes are the male gonads—the organs where the male reproductive cells are formed (Figures 19-4 and 19-5). They are generally oval, and their size varies considerably among species. In common domestic animals, they are located outside the abdomen in the inguinal (groin) region, housed in a sac of skin called the scrotum.
FUNCTIONS
The testes have two main functions: spermatogenesis and hormone production. Spermatogenesis is the process by which spermatozoa, the male gametes, are produced in the seminiferous tubules of the testes. Between the seminiferous tubules, cells called interstitial cells (interstitial means between) produce male sex hormones, or androgens. The principal androgen they produce is testosterone, which is responsible for the development of male secondary sex characteristics (such as the male body shape) and the male libido (sex drive). It also has a general anabolic (protein-building) effect on the body, which results in the enhanced muscle and bone development that gives male animals their size and bulk.
SPERMATOZOA
Spermatozoa are long, thin cells with three main parts: an enlarged head, a midpiece, and a long, narrow tail (Figure 19-6). The head contains the payload of the cell, the nucleus, and is covered by a caplike structure called the acrosome. The acrosome contains digestive enzymes that are released once the spermatozoon is inside the female reproductive tract. It helps the spermatozoon penetrate through the layers surrounding the ovum to accomplish fertilization. The midpiece is the “power plant” of the cell. It contains a large concentration of energy-producing mitochondria arranged
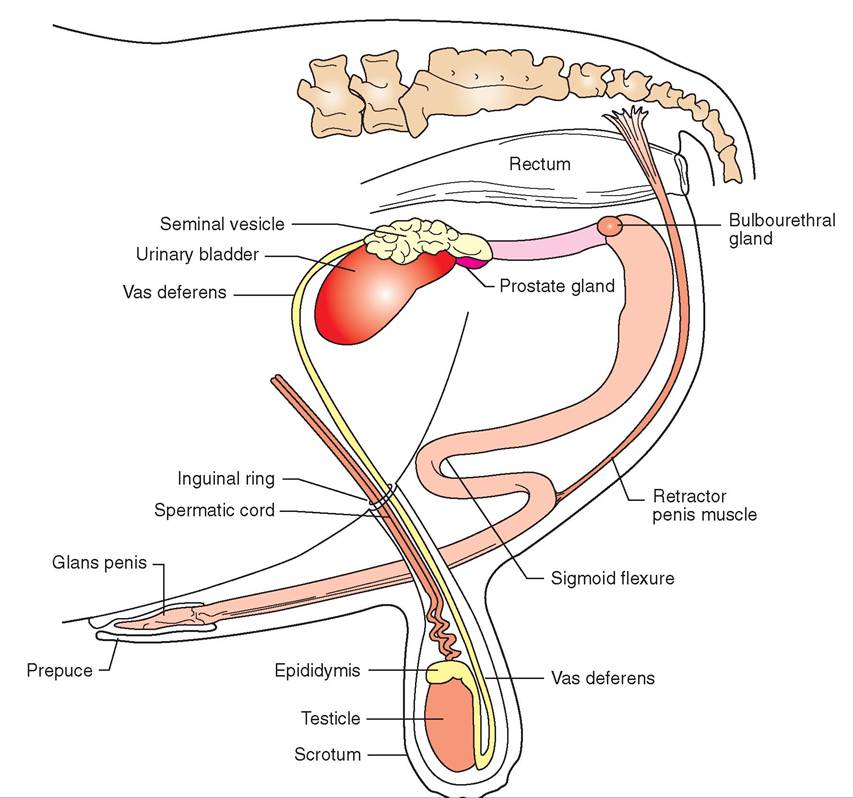
FIGURE 19-2 Reproductive system of the bull. Lateral view.
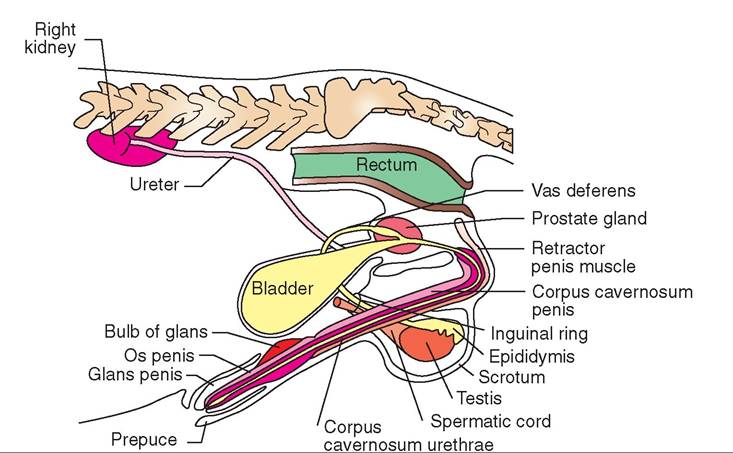
FIGURE 19-3 Male Urinaryandreproductive Ofgans ofthedoa∙ Latera I view.
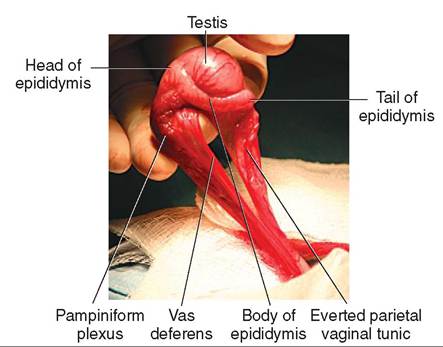
FIGURE 19-4 Canine testis. Lateral view.
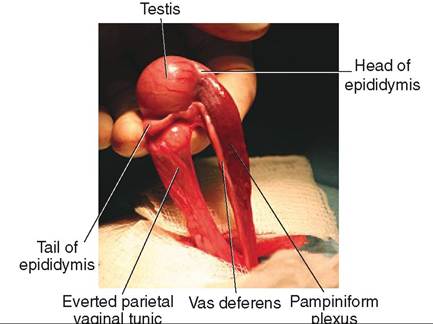
FIGURE 19-5 Canine testis. Medial view.
in a spiral pattern. The long, thin tail resembles the flagellum that many single-celled organisms use for propulsion. It contains musclelike contractile fibrils that produce a whip-like movement of the tail and propel the cell forward once it is activated.
DEVELOPMENT AND LOCATION
Before birth, the testes begin development in the abdominal cavity near the kidneys. They are attached to their eventual permanent home in the scrotum by a band of connective tissue called the gubernaculum. As the tiny embryo grows, the gubernaculum remains fairly constant in length. This results in the testes gradually being pulled caudally and ventrally. What is actually happening is that the fetus is growing longer and larger while the testes are being held in place. At or soon after birth, the testes are normally pulled down through the inguinal rings into the scrotum. The inguinal rings, located in the groin region, are two slitlike openings in the abdominal muscles. This process is referred to as the descent of the testes and is illustrated in Figure 19-7.
SCROTUM
The scrotum is the sac of skin that houses the testes. Besides providing a home for the testes, the scrotum also helps
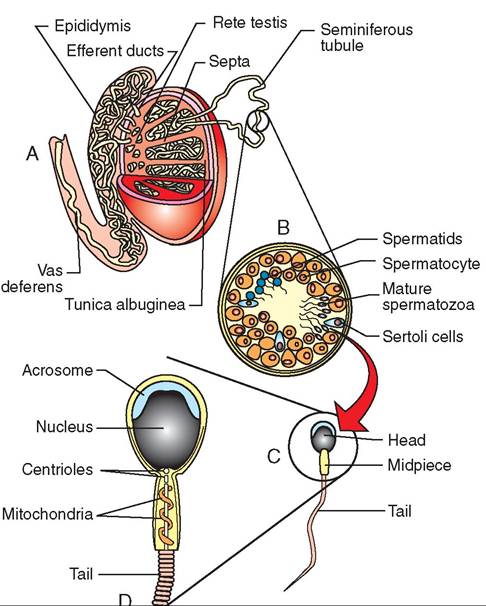
FIGURE 19-6 Structure of a testis. A, Testis and epididymis. B, Enlarged cross section of seminiferous tubule. C, Mature spermatozoon. D, Head area detail of mature spermatozoon.
regulate their temperature. The testes have to be kept slightly cooler than body temperature to produce spermatozoa. A bandlike muscle, the cremaster muscle, passes down through the inguinal ring and attaches to the scrotum. It can adjust the position of the testes relative to the body. In warm conditions, the cremaster muscle relaxes, and the testes hang down away from the warm body. This position helps to reduce their temperature. In cold conditions, however, the cremaster muscle pulls the testes up tight against the body wall, which helps warm them.
SPERMATIC CORD
The spermatic cords link the testes with the rest of the body. They are tubelike, connective tissue structures that contain blood vessels, nerves, lymphatic vessels, and the vas deferens. The arrangement of blood vessels in the spermatic cord is particularly interesting. They form a heat-exchange mechanism that helps keep the temperature of the testes slightly lower than that of the rest of the body. As Figures 19-8 and 19-9 show, the single testicular artery carries blood down to the testis. Surrounding the artery is a structure called the pampiniform plexus, which is an intricate meshwork of tiny veins derived from the testicular veins. As body-temperature blood passes down to the testis through the testicular artery, it is cooled by the blood returning from the testis in the pampiniform plexus. At the same time, the blood in the pampiniform plexus is warmed by the blood in the testicular
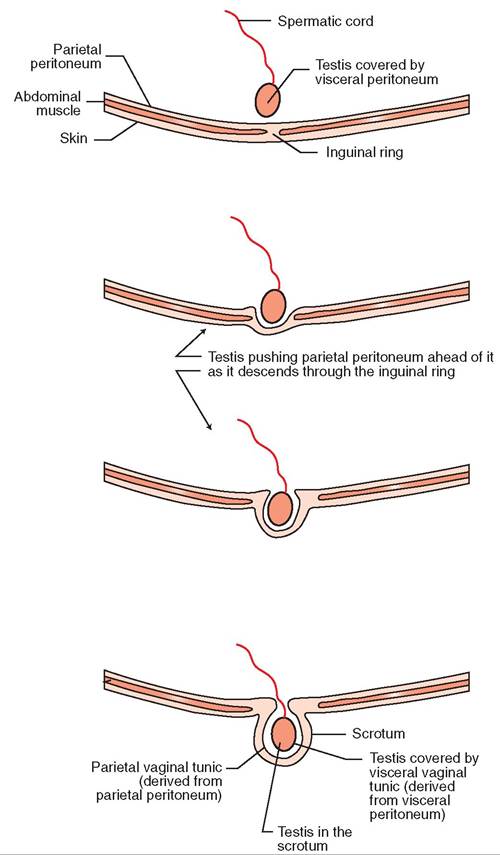
FIGURE 19-7 Descent of the testes and derivation of the vaginal tunics.
artery. This heat-exchange mechanism helps maintain the testes at a temperature slightly lower than body temperature but keeps the body as a whole from losing heat by warming the blood from the testes back to body temperature before it returns to the abdomen.
STRUCTURE
TUNICS. Two layers of connective tissue called the vaginal tunics surround the testes in the scrotum and the spermatic cord. The “vaginal” part of the name refers to the fact that the tunics form sheathlike covering layers. They are derived from layers of peritoneum that were pushed ahead of the testes as they descended through the inguinal rings (see Figure 19-7). The thin, inner layer, the visceral vaginal tunic (also known as the proper vaginal tunic), is derived from the visceral layer of peritoneum that coated the testes as they developed in the abdomen. It is tightly adherent to the surface of the testes and the structures of the spermatic
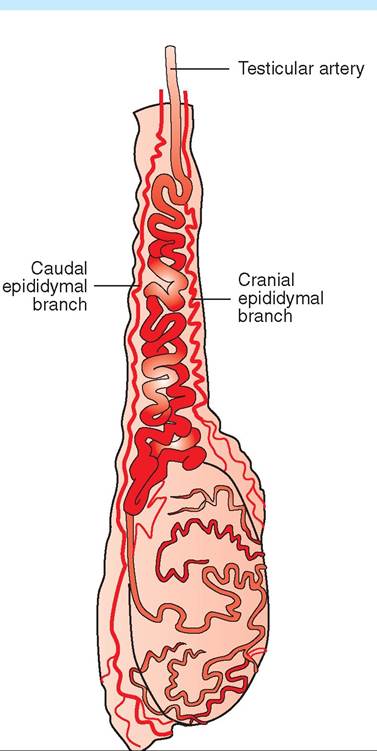
FIGURE 19-8 Arterial blood supply to testis and epididymis of a bull. The testicular artery passes down the center of the spermatic cord.
cords. It is so thin and transparent that it is invisible to the naked eye. The thick, outer layer, the parietal vaginal tunic (also known as the common vaginal tunic), is derived from the parietal layer of peritoneum that lines the abdominal cavity. It forms a visible fibrous sac around each testis and spermatic cord.
The tunics become significant when an animal is to undergo a castration (orchiectomy). An incision into the scrotum will reveal the tunic-covered testis. In one common method of castration, an incision is made in the outer, parietal vaginal tunic, and the testis is then everted through the incision in the tunic, thereby exposing the blood vessels in the spermatic cord. The blood vessels can then be ligated (tied off) or clamped for a time before the testis is removed. Figure 19-10 shows the initial incision through the parietal vaginal tunic during a dog castration. Note the testis, covered in the visceral vaginal tunic, bulging up through the incision in the parietal tunic.
CAPSULE. Beneath the tunics, each testis is enclosed by a heavy, fibrous connective tissue capsule called the tunica albuginea (see Figure 19-6). This capsule protects and supports the soft contents of the testis. Many small partitions,
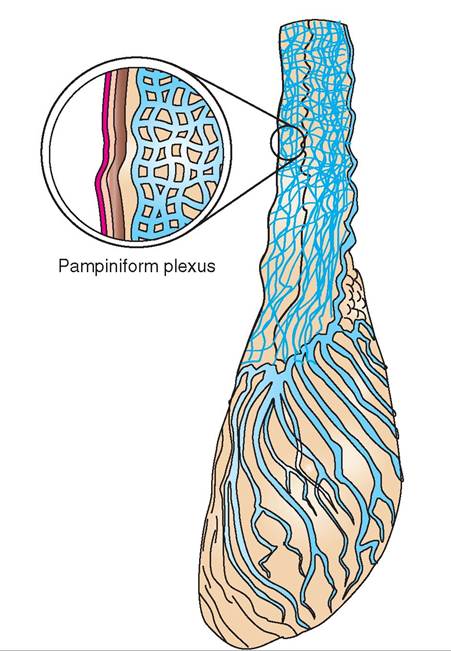
FIGURE 19-9 Pampiniform plexus. Formed from testicular veins visible on the surface of the testis, it surrounds the testicular artery and helps cool the blood passing down to the testis.
androgens, the male sex hormones, under the influence of luteinizing hormone (LH) from the anterior pituitary gland. LH is sometimes referred to as interstitial cell-stimulating hormone (ICSH) in the male.
Inside the seminiferous tubules (see Figure 19-6), spermatozoa are produced by the process of meiosis (see Figure 19-1). The “parent” cells stay out toward the periphery of the tubule and push the “daughter” cells toward the center of the tubule as they divide. Primary spermatocytes divide to produce secondary spermatocytes, which divide to produce spermatids. The spermatids undergo physical changes to become spermatozoa. While they are undergoing these transformations, the spermatids are attached to large “nurse” cells called Sertoli cells.
In addition to providing mechanical and nutritional support, the Sertoli cells also help shield the developing spermatozoa from the body's immune system. Proteins on the surfaces of the genetically unique spermatozoa would stimulate the immune system to produce antibodies against them if they were not shielded. This would cause an autoimmune reaction, meaning the immune system would attack part of its own body—the spermatozoa. The Sertoli cells are also known to produce small amounts of estrogens under the stimulation of follicle-stimulating hormone (FSH) from the anterior pituitary gland.
CLINICAL APPLICATION
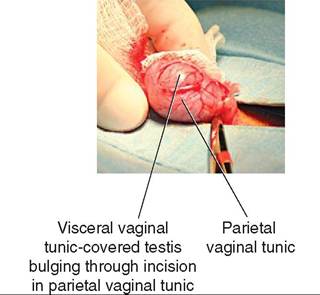
FIGURE 19-10 Tunics of the canine testis.
Cryptorchidism
Sometimes one or both of the testes do not descend into the scrotum. This is referred to as cryptorchidism. This condition can be either unilateral, when one testis has failed to descend, or bilateral, when both testes have failed to descend. An undescended testis can be located anywhere along the normal path of descent, from the region of the kidney to an area just outside the inguinal ring but not completely down in the scrotum (called a “high flanker”). Testes retained in the abdominal cavity are usually sterile (cannot produce spermatozoa) because spermatogenesis requires a temperature slightly lower than body temperature. The interior of the abdomen is too hot for spermatozoa to be produced. However, testosterone continues to be produced, so a bilaterally cryptorchid animal has all the characteristics of a male animal but typically cannot reproduce.
called septa, extend into the testis from the capsule. The septa divide each testis into tiny lobules that contain the seminiferous tubules, as well as other cells and structures.
SEMINIFEROUS TUBULES. The seminiferous tubules are where spermatogenesis takes place. Each tubule is shaped like a long, convoluted “U” that is attached at both ends to a complex system of ducts called the rete testis (a rete is a complex network of something in the body). Between the seminiferous tubules are the endocrine cells of the testis known as the interstitial cells. The interstitial cells produce DUCT SYSTEM. When spermatozoa complete their physical development in the seminiferous tubules, they are transported to a storage site where they will be prepared for ejaculation. After they detach from their protective Sertoli cells, the spermatozoa enter the complex of ducts that make up the rete testis. From there, they flow through the efferent ducts of the testes to their storage site—the epididymis.
Grossly, the epididymis is a flat, ribbonlike structure that lies along the surface of the testis (see Figures 19-4 and 19-5). It is actually a single, long, very convoluted tube that connects the efferent ducts of the testis with the vas deferens. If the tube that makes up the human epididymis were stretched out, it would be about 6 m (20 feet) in length. The epididymis is divided into three regions: (1) the head of the epididymis is where the spermatozoa enter from the efferent ducts, (2) the body is the main portion that lies along the surface of the testis, and (3) the tail continues on as the vas deferens.
The epididymis functions as a storage site for spermatozoa and a place for them to mature before they are expelled by ejaculation. Spermatozoa are immature when they leave the seminiferous tubules. They must mature in the epididymis for a week or more before they can fertilize ova. If the spermatozoa are never ejaculated (e.g., in an individual who has had a vasectomy), they live out their lives in the epididymis, die, and are broken down and absorbed by the tubular lining cells.
Clinical application
Sertoli Cell Tumor
On occasion, Sertoli cells can multiply out of control, particularly in dogs. This produces what is termed a Sertoli cell tumor in the testis. Because normal Sertoli cells produce small amounts of estrogens, this action can become exaggerated as the cells multiply out of control. The level of estrogens in the animal’s bloodstream rises rapidly, producing feminization of the male dog. The prepuce often becomes pendulous, and the mammary glands may become enlarged (gynecomastia) and can even produce milk. The penis and the opposite testis atrophy, and other male dogs may be attracted to the affected animal as if he were a female in heat. The signs of feminization are often the primary complaints when affected animals are presented to the veterinary clinic. The primary treatment for a Sertoli cell tumor is usually castration of the animal.
TEST YOURSELF 19-3
1. What are the two main functions of the testis? Where in the organ does each take place?
2. What are the three main parts of a spermatozoon? What is the main purpose or function of each?
3. Why is a bilaterally cryptorchid animal usually sterile?
4. Would a bilaterally cryptorchid animal exhibit normal male behavior? Why or why not?
5. What is important about the scrotum's ability to adjust the position of the testes relative to the body?
6. What are the main components of the spermatic cord?
7. From what are the visceral and parietal vaginal tunics that cover the testes derived?
8. Where are spermatozoa stored before ejaculation?
VAS DEFERENS
Something has to move the spermatozoa from the epididymis (located way down in the scrotum) up to the urethra (within the pelvic cavity) when ejaculation occurs. This is the job of the vas deferens. Also known as the ductus deferens, the vas deferens is a muscular tube that connects the tail of the epididymis with the pelvic portion of the urethra. Thick layers of smooth muscle in its wall give it a very solid, cordlike texture. The vas deferens passes up through the inguinal ring as a part of the spermatic cord. Once inside the abdomen, each vas deferens separates from the rest of the spermatic cord, loops back caudally, and connects with the urethra just caudal to the neck of the urinary bladder (see Figures 19-2 through 19-5, and Figure 19-11). In many species, the vas deferens enlarges just before joining the urethra. This enlargement, if present, is called the ampulla, which may contain glands that contribute material to semen.
The job of the vas deferens is to propel spermatozoa and the fluid they are suspended in quickly from the epididymis to the urethra when ejaculation occurs. Once they are in the urethra, the spermatozoa are mixed with secretions from accessory reproductive glands to form semen, which is pumped out into the female reproductive tract.
URETHRA
The urethra of the male has two functions. Most of the time it carries urine from the urinary bladder out of the body. This is its urinary function. However, when ejaculation occurs, urine flow is temporarily blocked. Spermatozoa from the vas deferens and secretions from the accessory reproductive glands enter the urethra and are pumped out as semen. Ejaculation is its reproductive function.
The urethra is spoken of as having two portions. The pelvic portion, within the pelvic cavity, is where the vas deferens and accessory reproductive glands enter. The remainder of the urethra, which runs down the length of the penis, is known as the penile portion of the urethra.
ACCESSORY REPRODUCTIVE GLANDS
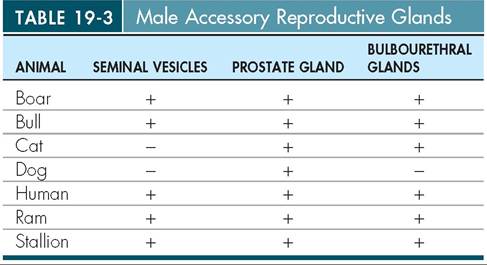
Spermatozoa make up only a small portion of the total volume of semen. The majority is made up of secretions from the various accessory reproductive glands. The ducts of all the accessory reproductive glands enter the pelvic portion of the urethra (see Figure 19-11). Different species have different combinations of accessory reproductive glands. Table 19-3 provides a summary of which common animals have which glands.
The accessory reproductive glands produce alkaline fluid containing substances such as electrolytes, fructose, and prostaglandins. The alkalinity of the fluid helps counteract the acidity of the female reproductive tract. This helps more spermatozoa survive to reach the oviducts, where they hope to meet up with an ovum. Fructose is a simple sugar that acts as an energy source for the very active spermatozoa. Some of the prostaglandins are thought to stimulate the contractions of the female reproductive tract which help move the spermatozoa up to the oviducts.
SEMINAL VESICLES
Ducts from the two seminal vesicles, sometimes called vesicular glands, enter the pelvic urethra in the same area as the
+, Indicates the presence of a gland; -, indicates the gland is absent.
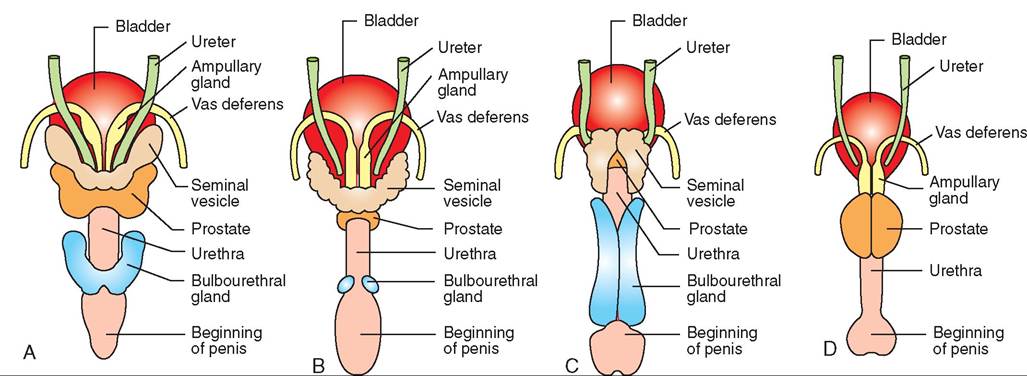
FIGURE 19-11 Accessory reproductive glands. Comparisons among some common domestic species (dorsal views). A, Stallion. B, Bull. C, Boar. D, Dog.
CLINICAL APPLICATION
Vasectomy
If a section of each vas deferens is removed surgically from an animal, the animal is rendered sterile (it cannot reproduce). This procedure is known as a vasectomy. It prevents spermatozoa from reaching the urethra during ejaculation. The othen components of semen are ejaculated, but no spermatozoa are included. The individual is “firing blanks,” so to shpeeak. T spermatozoa live out their lives in the epididymis, edie, and ar absorbed by the lining cells.
Vasectomy is a common contraceptive surgery to prevent pregnancy in humans. It is performed less commonly for that reason in nonhuman animals but is sometimes per- fnormed o bulls to create “teaser” animals. Teaser bulls are loupsed t he identify cows in heat so that they can be bred, tyoiffitecnialb ar insemination, to other males.
vas deferens. They are present in all common domestic animals except for cats and dogs.
PROSTATE GLAND
The prostate gland is a singe structure that more or less completely surrounds the urethra. Multiple ducts carry its
secretions into the urethra. It is present in all common domestic animals. In dogs, it is particularly large because it icnsclyethsseoory a reproductive gland they have.
BULBOURETHRAL GLANDS
The two bulbourethral glands, also known as the Cowper’s glands, are located further caudally than the other accessory reproductive glands. Their ducts enter the urethra back near the caudal border of the pelvis. They secrete a mucinous (mucus-containing) fluid just before ejaculation that clears and lubsicates the urethra for the passage of semen. All coommesmtiocn d animals have bulbourethral glands eacept foogr. the d
PENIS
The penis is the male breeding organ. It is made up mainly of muscle, erectile tissue, and connective tissue, with the urethra running down its center. It has a very large blood supply and many sensory nerve endings. When the male is sufficiently aroused and stimulated, the erectile tissue bnegcoormgeeds e with blood, causing the penis to enlarge
iafnfedn.st This allows it to be inserted into the vagina of teohmreafle f breeding.
The three main parts of the penis are the roots, the body, and the glans.
ROOTS
The roots of the penis attach it to the brim of the pelvis. They consist primarily of two bands of connective tissue, called the crura, corered by the ischiocavernosus muscles.
BODY
The body of the penis is its largest part. It is mainly made up o0 two bundles of erectile tissue. Erectile tissue is com- fposed o a spongy network of fibrous connective tissue and many tlny, blood-filled spaces called sinuses. Ier the nonerect state, the amount of blood flowing into the erectile tissue is the same as the amount flowing out. When more blood flows into erectile tissue than leaves it, however, the sinuses engorge with blood and try to enlarge. The network of connective tissue surrounding the engorged sinuses prevents them from swelling up like a bunch of water balloons. Instead, the relatively inelastic connective tissue around the blood sinuses generates enough hydraulic pressure in the engorged erectile tissue to cause the penis to become a little larger and a lot stiffer. This is called erection of the penis.
The two erectile tissue structures that make up the body of the penis are the smaller corpus cavernosum urethrae and the larger corpus cavernosum penis. The corpus cavernosum urethrae, also called the corpus spongiosum, forms a sleeve around the urethra as it passes through the body of the penis. The larger body of erectile tissue dorsal to the urethra is the corpus cavernosum penis.
CLINICAL APPLICATION
Canine Prostate Problems
The prostate gland in dogs is large—about the size of a walnut in a medium-size dog. Many conditions can cause it to become even larger, such as infection, tumors, or just normal aging. Because the urethra runs through the center of the gland, significant enlargement of the prostate squeezes the urethra. This can partially or completely block the passage of urine, leading to difficulty urinating. Instead of urine passing in a large stream, it dribbles out or may stop entirely. The cause of prostate gland enlargement must be determined to formulate an effective treatment strategy. Some conditions can be managed with medication, but others require surgery.
GLANS
The glans of the penis is its tip, or the distal, free end of the organ. Its structure and appearance vary considerably among species. Horses have a well-defined glans that contains a considerable amount of erectile tissue. In ruminants, the glans is small and not well defined. Cats have short spines covering their glans. The penis of the dog is a special case that is described more completely below. Regardless of the species, the surface of the glans has a rich supply of sensory nerve endings that make it very sensitive to physical stimulation.
PREPUCE
The prepuce is the sheath of skin that encloses the penis when it is not erect. The outer part of the prepuce is normal skin, but the inner portion that is in contact with the penis is a smooth, moist, mucous membrane. The prepuce of the boar also contains a small preputial pouch in which urine and cellular debris accumulate. Decomposition of the material in this pouch gives boars their typical ripe odor. (This is considered very sexy to sows—a clear case of different strokes for different folks!)
PENIS OF THE DOG
The penis of the dog and other canine species is unusual (see Figure 19-3). It includes a bone and an erectile structure that causes the male and female to remain stuck together after breeding is completed. The bone in the penis of the dog is called the os penis (see Figure 7-37). The urethra runs through a groove in the ventral surface of this bone. No other domestic animals have a similar bone, but several wildlife species do, including raccoons, beaver, and walruses. It is rarely of clinical significance unless it is fractured (very rare) or unless urinary stones (calculi) lodge in the urethra, where it enters the groove in the os penis, and obstruct urine flow.
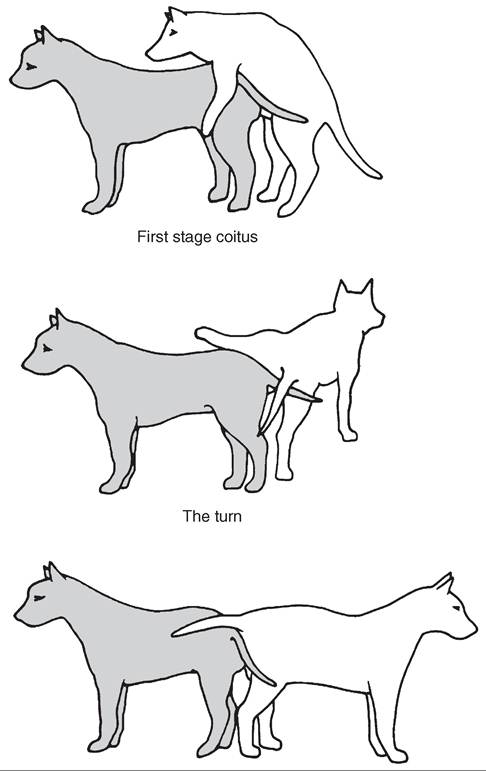
The other unique structure in the penis of the dog is an enlargement toward the rear of the glans called the bulb of the glans. It is made up of erectile tissue that is derived from the corpus cavernosum urethrae. During breeding, the bulb becomes engorged with blood more slowly than the other erectile structures. It usually does not reach full size until after ejaculation has taken place. Once it enlarges, however, the swollen bulb is tightly clamped in place by contractions of the muscles surrounding the vagina and vulva of the female. This makes it impossible for the male to withdraw the penis. The male then typically dismounts from the female and turns so that he faces the opposite direction from her. The two stand in a tail-to-tail position looking for all the world like the fictional “Pushme-Pullyou” from the Dr. Doolittle story. This is known as the tie and seems to be important for conception to occur in the dog. The tie typically lasts 15 to 20 minutes and does not seem uncomfortable for either animal. Once the erection of the bulb subsides, the animals can separate. Forcing the animals apart prematurely can cause injury. So tell well-meaning people not to throw cold water on a pair of tied dogs in an effort to separate them. Just let nature take its course. The positions of dogs during copulation are shown in Figure 19-12.
PENIS OF THE HORSE
The penis of the horse contains a large component of erectile tissue, and the glans at the distal end flairs into a distinctive mushroom shape, the widest part of which is called the corona glandis (Figure 19-13). The distal end of the urethra, the urethral process, extends from a central depression, the fossa glandis, at the tip of the penis. Because of the large amount of erectile tissue it contains, the penis of the horse enlarges considerably in both length and circumference when erect.
SIGMOID FLEXURE
The nonerect penis of the bull, ram, and boar is normally bent into an “S” shape. This is called the sigmoid flexure (see Figure 19-2); sigmoid means S-shaped. The penises of these animals also have a higher proportion of connective tissue to erectile tissue than other species, and so the penis does not enlarge much when erection occurs. Rather, the main mechanism of erection in these species is a straightening of the sigmoid flexure resulting from internal hydraulic pressure. The basic principle is the same as in a garden hose that wants to straighten out when the water is turned on. This causes the penis to protrude from the prepuce for breeding. A long, thin, cordlike muscle, the retractor penis muscle,
Second stage coitus
FIGURE 19-12 Illustration of the three most obvious stages of breeding in dogs. (From Feldman E, Nelson R: Canine and feline endocrinology & reproduction, ed 3, Philadelphia, 2004, WB Saunders Company.)
originates near the base of the tail and attaches to the bend of the sigmoid flexure. It functions like a small bungee cord: When erection straightens out the sigmoid flexure, the retractor penis muscle stretches. When the erection subsides, the retractor penis muscle pulls the penis back into its nonerect, S shape.
REPRODUCTIVE FUNCTIONS
ERECTION. Erection enlarges and stiffens the penis to prepare it for breeding. See Chapter 20 for information about the processes of erection.
EJACULATION. Ejaculation is the process by which semen is expelled from the penis. See Chapter 20 for more information about ejaculation.
FEMALE REPRODUCTIVE SYSTEM
The female part of the reproductive system is a bit more complex than the male part, because it has more jobs to do.
TEST YOURSELF 19-4
1. Will the volume or gross appearance of the semen of an animal that has had a vasectomy be significantly different from that of a normal animal? Why or why not?
2. How does the function of the bulbourethral glands differ from those of the other accessory reproductive glands?
3. What are the three main parts of the penis?
4. What structures do spermatozoa pass through from their site of production to their eventual deposition in the female reproductive tract?
Some are similar to those of the male, such as the production of sex hormones and development of gametes. But, in addition, the female system receives the male gametes, furnishes a site for them to fertilize the ovum (the oviduct), provides a hospitable environment for the embryo to grow and develop, carries it for the entire period of pregnancy, and then pushes the offspring out into the world when it is fully developed. All these jobs do not stop at birth: In the period immediately after, the mammary glands provide nutrition to the newborn. This is a lot of work for one system to do.
Nearly all of the female reproductive system is internal and is located within the abdominal and pelvic cavities. Figures 19-14 and 19-15 illustrate the female reproductive system of the bitch, and Figure 19-16 compares the female reproductive systems of several species; Figures 19-17 and 19-18 show reproductive structures of the cat.
LIGAMENTS
The main reproductive organs—the ovaries, oviducts, and uterus—hang by broad sheets of peritoneum from the dorsal part of the abdominal cavity. The sheets of peritoneum are collectively called the left and right broad ligaments (see Figure 19-17). Each broad ligament has segments that are named according to the organ they directly support, although the divisions are not clearly apparent at a quick glance. The mesovarium supports the ovary, the mesosalpinx supports the oviduct, and the mesometrium supports the uterus. (The term meso refers to a sheet of tissue that attaches an organ to the body wall.) The broad ligaments contain blood vessels and nerve fibers that supply the ovaries, oviducts, and uterus; they also contain fat.
The ovarian end of the broad ligament extends cranially and attaches to the body wall in the area of the last rib. This forms the suspensory ligament of the ovary (see Figure 19-18). During surgical removal of the ovaries and uterus (ovariohysterectomy) in dogs and cats, the suspensory ligament must often be stretched or divided to allow the ovary to be elevated into the incision, so the ovarian blood vessels can be tied off.
A cord of fibrous tissue and smooth muscle called the round ligament of the uterus is contained in the free edge of a lateral fold of the broad ligament on each side
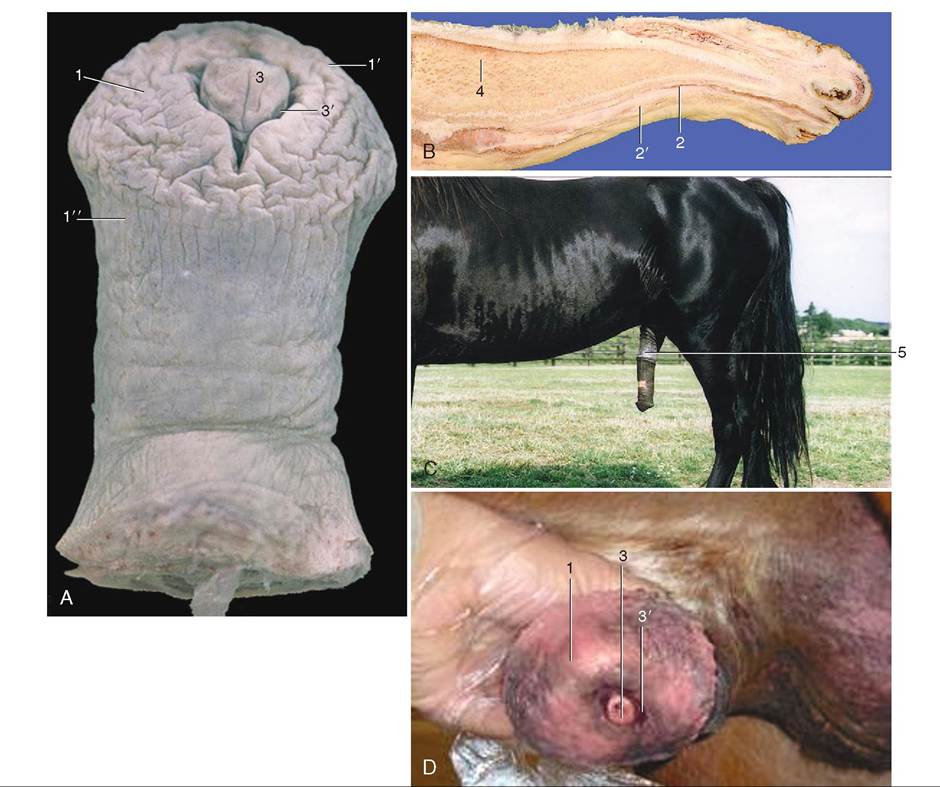
FIGURE 19-13 Penis of the horse. A, Distal portion of penis; B, median section; C, penis just after copulation; D, distal tip of glans. 1, Glans; 1', corona; 2, urethra; 2' corpus cavernosum urethrae; 3, urethral process within 3' fossa glandis; 4, corpus cavernosum penis; 5, prepuce. (From Dyce KM, Sack WO, Wensing CJG: Textbook of veterinary anatomy, ed 4, St Louis, 2010, W.B. Saunders Company.)
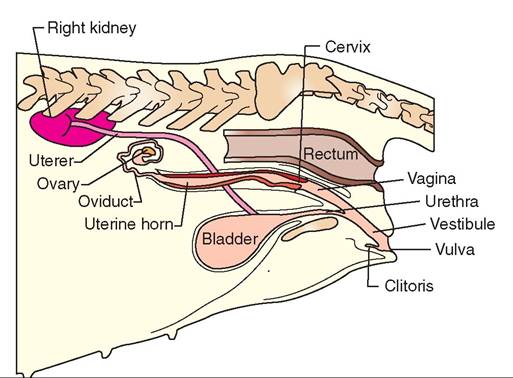
FIGURE 19-14 Urinary and reproductive organs of bitch. Lateral view.
(see Figure 19-17). The round ligament extends from the tip of the uterine horn caudally and to the inguinal ring area ventrally. During ovariohysterectomy in dogs and cats, the round ligament is transected (cut) along with the broad ligament to allow the uterine horns to be removed.
OVARIES
CHARACTERISTICS
The ovaries are the female gonads; they are the female equivalent of the male testes. They are located in the dorsal part of the abdominal cavity near the kidneys. Their shape varies among species; in most, they are somewhat almond shaped, but the ovaries of the horse are indented, making them beanlike in appearance. The ovaries of sows, because their large litter sizes require large numbers of follicles, often look like clusters of grapes.
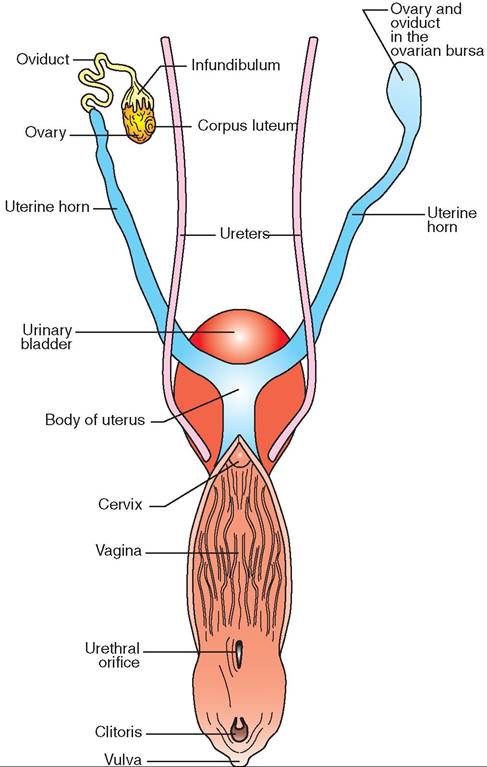
FIGURE 19-15 Reproductive system of bitch. Dorsal view.
CLINICAL APPLICATION
Ovariohysterectomy
Ovariohysterectomy is a surgical procedure in which the ovaries, oviducts, and uterus are removed from an animal. It is commonly known as spaying an animal. Despite the frequency Wlr which it is performed, ovariohysterectomy is a major surgical procedure that involves opening the abdominal cavity. To remove the reproductive organs safely, the blood vessels supplying them must first be tied off (ligated). The ebsloseolsd v to each ovary are usually ligated along with the portion of the broad ligament (mesovarium) that contains them. This portion is referred to surgically as the ovarian pedicle or ovarian stump. Tlie ovaries then can be severed sraofmely f their ligated blood vessels. The remainder of the ibgraomadenlt is usually either ligated en masse or cut back thoeward t body of the uterus. The blood vessels in this part rohofeatd b ligament are usually small. The body of the uterus and its accompanying blood vessels are ligated and transected (ehcpeurot)d. uTctiver organs then can be removed safely
from file abdominal cavity and the incision sutured closed.
FUNCTIONS
Like the testes, the ovaries have two main functions: producing gametes and producing hormones. Oogenesis is the yrocess by which ova (the female gametes) are produced in follicles in the ovaries. Unlike spermatozoa, ova are not con- rtantly produced during the reproductive life of the animal. At Ot soon after birth, the ovaries are seeded with tens of thousands of immature reproductive cells called oocytes. Some of these oocytes will mature into ova, the mature feepmroadleucrtive cells, through the activities of the
ovarian cycle. The rest will either degenerate or never begin development. No more oocytes are produced during the
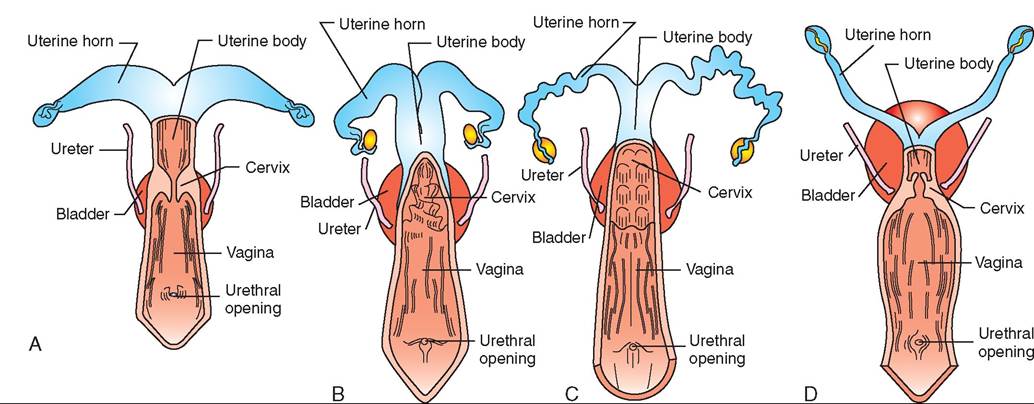
FIGURE 19-16 Femdle reproacctive tract. Comaarisons omongsome common dpmestic soθcies (dorsal views). A, Mare. B, Cow. C, Sow. D, Bitch.
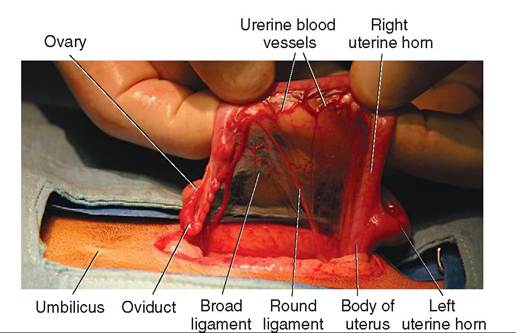
FIGURE 19-17 Reproductive structures of female cat. Lateral view.
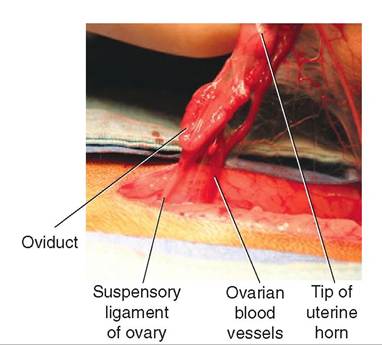
FIGURE 19-18 Reproductive structures of female cat. Lateral view.
animal's life, however. The number of oocytes in the ovaries soon after birth is the maximum number that will be available to that animal during her lifetime.
The hormones produced in the ovaries fall into two categories: estrogens and progestins. Estrogens are produced by the cells of the developing ovarian follicles and are responsible for the physical and behavioral changes that prepare the animal for breeding and pregnancy. Progestins, principally progesterone, are produced by the corpus luteum that develops from the empty follicle after ovulation. Progestins help prepare the uterus for implantation of a fertilized ovum. They are also necessary for pregnancy to be maintained once implantation occurs.
OVARIAN CYCLE
Ova are not constantly produced in the ovaries. Their production involves a complex sequence of events carried out in a cyclic (repeating) fashion under the influence of two hormones from the anterior pituitary gland: folliclestimulating hormone (FSH) and luteinizing hormone (LH). Each cycle includes the development of an ovum within a follicle, its release from the follicle (called ovulation), formation of a corpus luteum, and the degeneration of unripened follicles and, eventually, the corpus luteum. Figure 19-19
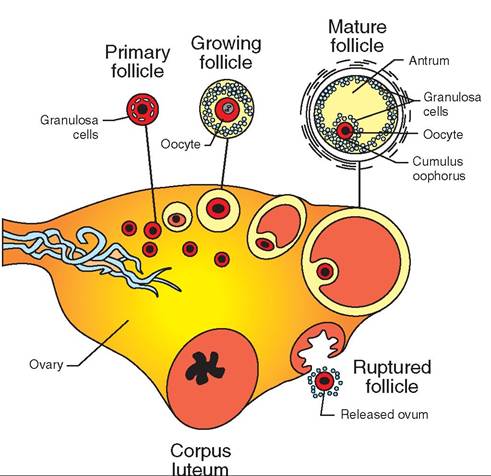
FIGURE 19-19 Ovarian cycle. Schematic representation of the sequence of events in the ovarian cycle, starting with the primary follicle and proceeding (clockwise) to the mature follicle, ovulation, and formation of the corpus luteum. These stages would not all be present at one time in an ovary.
illustrates the main sequence of events in this process, and Figure 19-20 shows steps in follicular development in cattle.
The number of follicles involved in each cycle depends on the species. Uniparous species, such as horses, cattle, and humans, normally give birth to only one offspring at a time. Their ovaries produce one mature ovum per cycle. Multiparous species, such as cats, dogs, and sows, give birth to litters of young. Their ovaries produce multiple ova per cycle.
The beginning stage of follicle development in the ovary is the primordial, sometimes called the primary, follicle. The thousands of immature oocytes in the ovaries of newborn animals reside in this stage until they become activated later in life and begin to develop further. The primordial follicle consists of the immature gamete (the oocyte) surrounded by a single layer of flattened follicular cells. When FSH is released from the anterior pituitary, the whole ovary is bathed in it. Something, however, causes just a few of the thousands of primordial follicles to begin developing. This is known as follicular recruitment or follicular activation.
Once a primordial follicle has become activated, it is referred to as a growing follicle. The oocyte starts to grow in size, the glycoprotein zona pellucida layer forms just outside the cell membrane, and the follicular cells enlarge into cuboidal shapes and begin to multiply. Multiple layers of follicular cells form around the zona pellucida of the developing oocyte. At this stage, the follicular cells are called granulosa cells. As the granulosa cells multiply, the follicle starts to grow rapidly in size. The granulosa cells do more than just physically surround the developing oocyte. They also produce estrogen hormones that begin preparing the animal for breeding and pregnancy. The larger the follicle
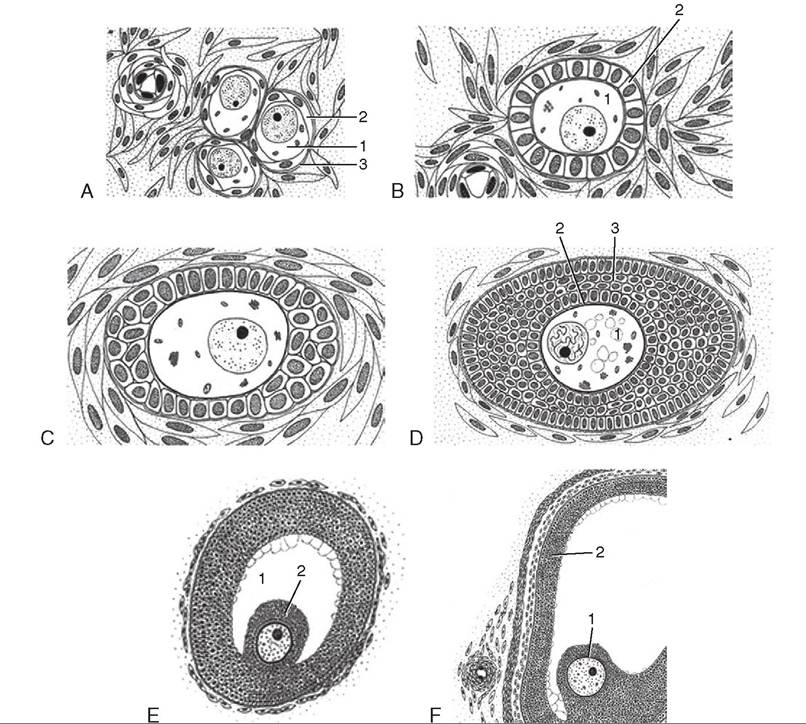
FIGURE 19-20 Folliculcir devel cpmentin cαttle.(Not to scale.) A, Primordial follicle. The oocyte (1) is surrounded by flat follicular cells (2) resting on a basement membrane (3). B, Primary follicle. The oocyte (1) is surrounded by cuboidal granulosa cells (2). C, Early growing follicle. D, Slightly larger early growing follicle. The oocyte (1) is surrounded by the developing zona pellucida (2) and a multilayered granulosa cell compartment (3). E, Larger growing follicle showing the antrum (1) and the oocyte located in the cumulus oophorus (2). F, Large growing follicle presenting the oocyte in the cumulus oophorus (1) and the granulosa cell layer (2) surrounding the antrum. (Courtesy Sinowatz and Russe (2007). From Hyttel P, Sinowatz, F, Vejlsted M: Essentials of domestic animal embryology, St Louis, 2010, Saunders.)
becomes, the greater the amount of estrogens produced. As the follicle continues to grow, fluid-filled spaces begin to form between the granulosa cells. The spaces gradually become confluent (they join together), forming one large, fluid-filled space called the antrum.
When the follicle has reached its maximum size, it looks like a large, blisterlike structure on the surface of the ovary. At this stag, it is called a mature fofficle. Alternvfive names for the matae follicle include Graafian follicle and vesicular ovarian follicle. Inside the mature follicle, the oocyte sits on a little mound of granulosa cells called the cumulus oophorus, and its zona pellucida is surrounded by a thin layer of granulosa cel Is called the corona radiata. Most of the volume oufrethe mat follicle is made up of the fluid-filled antrum. eA,t this stag estrogen production from the follicle is at a Veak level, and the animal is ready for breeding to take place. In most animal species, ovulation (rupture of the mature follicle with release of the reproductive cell into the oviduct) oceans spontaneously as a result of the rising level of LH. eTnheirsalgly occurs regardless of whether breeding has tcaek. en pla In some species, however, such as cats, camels, rabbits, and ferrets, breeding must take place before ovulation can occur. For this reason, cats, camels, rabbits, and ferrets are called induced ovulators. (TIus is also why the heat fperiods o cats can be so prolonged if they are not bred.) Once the mature gamete is released from the follicle, its name changes. It is now called an ovum.
Ovulation is a traumatic and somewhat explosive event. uTrhfea cse of the mature follicle weakens and physically
ruudpdteunrleys, s releasing the fluid from the antrum. The
frush o fluid out of the follicle carries the ovum with it, still surrounded by its halo of granulosa cells, the corona radiata. oTmlhlpiect ylee f fills with blood that rapidly clots, forming
the corpus Iiemorrhagicum.
Under continued stimulation by the high LH level, the eglrlasnulosa c that line the corpus hemorrhagicum begin to multiply again. This time they form a solid structure, the corpus luteum, that gets about as large as the mature follicle was just before ovulation. The term corpus luteum literally means “yellow body,” because it has a pale yellow color grossly (without magnification). The corpus luteum produces hormones called progestins. The principal progestin is progesterone, which is necessary for pregnancy to be maintained if the ovum is fertilized. If the ovum has been fertilized and implants in the uterus, it sends an endocrine signal to the ovary that causes the corpus luteum to be maintained. If the ovum has not been fertilized, no “we are pregnant” endocrine signal is sent and the corpus luteum degenerates after a short period.
Not all follicles that were activated in a particular ovarian cycle fully develop and ovulate. It is as if the ovary “auditions” follicles and chooses particular ones to fully develop. The rest may degenerate at any stage of their development. This is called follicular atresia and is a normal part of each ovarian cycle.
TEST YOURSELF 19-5
1. What two main types of hormone are produced in the ovary? Where is each produced?
2. What changes does an ovarian follicle undergo as it develops from a primordial follicle to a mature follicle?
3. After ovulation has occurred, what cells in the ovary multiply to form the corpus luteum?
OVIDUCTS
The two oviducts also are known as the Fallopian tubes and uterine tubes; they are small, convoluted tubes that extend from the tips of the uterine horns (see Figures 19-17 and 19-18). Their roles are to guide ova from the ovary to the uterus and to serve as the usual site for fertilization of ova by spermatozoa. The oviducts are not attached to the ovaries at all; when ovulation takes place, they have to “catch” the ova in the funnel-like infundibulum, which is the enlarged opening at the ovarian end of each oviduct. At the time of ovulation, it more or less surrounds the area of the ovar y where follicles have formed. Muscular, fingerlike projections called fimbriae form the margin of the infundibulum. They “feel” along the surface of the ovary and position the infundibulum where the follicles are located. This helps ensure that the infundibulum is properly positioned to catch the ova when ovulation occurs. If ova miss the opening of the oviduct, they fall into the abdominal cavity, where they usually just disintegrate after a time.
When examined closely, the complex physical structure of the oviducts becomes apparent. They contain many smooth muscle fibers in their walls. Their linings are very intricate and folded, and the lining cells are covered with countless, movable cilia. When an ovum enters the oviduct after ovulation, delicate muscle contractions and gentle movements of the cilia begin slowly and gently moving it toward the uterus. The greatly folded lining of the oviduct keeps it in continuous, gentle contact with the ovum. If breeding has taken place, spermatozoa are already up in the oviducts when ovulation occurs. The oviducts are the usual place where the ovum and some lucky spermatozoon come together. This is called fertilization of the ovum. Once fertilization has taken place, the oviducts gently conduct the fertilized ovum down to the uterus for implantation.
UTERUS
The uterus is the womb, where the fertilized ovum implants and lives while it grows and develops into a new animal. When the fetus is fully developed, the uterus helps push it through the birth canal out into the world. Although it seems like a simple receptacle in which the fetus can develop, the roles of the uterus are actually quite complex. It has to grow along with the developing offspring and then return to its original size after birth. It forms part of the placenta, which is the life-support system that keeps the fetus alive while it develops during pregnancy. The uterus must remain quiet during pregnancy, and it must contract powerfully at the time of birth. After it has delivered the newborn and the placenta (the afterbirth), it has to contract quickly to stop bleeding from the sites where the placenta was attached to its lining. It does not have the complex, cyclic functions of the ovaries or the intricate structure of the oviducts, but the uterus is vital to the success of reproduction.
Physically, the uterus is a hollow muscular organ. In common domestic animals, it is somewhat Y-shaped, with the uterine body forming the base of the Y and the two uterine horns forming the arms. The body of the uterus extends in a caudal direction, eventually joining with the cervix at its caudal end. The two uterine horns project crani- ally (see Figure 19-16). The oviducts extend from the tips of the uterine horns. The whole organ is suspended from the dorsal part of the abdomen by the mesometrium portion of the broad ligament.
The thick wall of the uterus is made up of three layers. The endometrium is composed mainly of simple columnar epithelium and simple tubular glands that secrete mucus and other substances. The thickest layer of the wall is the myometrium, which is made up of layers of smooth muscle that give the uterus the strength to push the fetus out at parturition. The outermost layer is the perimetrium, which is covered by the visceral layer of peritoneum.
The fertilized ovum implants in the uterus and begins to grow. As the offspring develops, the placenta forms around it and attaches to the lining of the uterus so that nutrients, wastes, and respiratory gases can be exchanged between the fetal bloodstream and the maternal bloodstream. (This is explained more fully in Chapter 20.) When the time comes for the offspring to be delivered, the muscular uterus provides most of the force necessary to open (dilate) the cervix so the fetus can pass through it on its way to the outside world.
CERVIX
The cervix is a muscular “valve” that seals off the uterus from the outside most of the time. It is a powerful, smooth muscle sphincter located between the body of the uterus and the vagina. It functions to control access to the lumen of the uterus from the vagina. The cervix is normally tightly closed, except at the two ends of pregnancy: estrus (the heat period) and parturition (the birth process). The cervix relaxes at estrus to admit spermatozoa during breeding. It then tightly closes again during pregnancy and does not relax again until birthing time. Uterine contractions during the first stage of labor push the newborn against the relaxed cervix and gradually pry it open (called dilation of the cervix) so that “junior” can slide down the birth canal and out into the world.
VAGINA
The vagina is the tube that receives the penis at breeding time and acts as the birth canal at parturition. It is a muscular tube that extends caudally from the cervix and connects it with the vulva. Although the lumen of the vagina is a “potential” space and is collapsed most of the time, it can stretch considerably to accommodate the penis at breeding and the newborn during the birth process. Mucous glands lining the vagina lubricate it at the time of breeding.
VULVA
The vulva is the only portion of the female reproductive system that is visible from the outside. Its main parts are the vestibule, the clitoris, and the labia. The word vestibule, in anatomic terms, means the entrance into a canal of some sort. In this case, the vestibule of the vulva is the entrance into the vagina from the outside world. It is the short space between the labia and the opening of the vagina. The urethra, the tube that carries urine out from the urinary bladder, opens on the floor (ventral portion) of the vestibule. The clitoris is also located on the floor of the vestibule a little nearer to the exterior than the urethral opening. The clitoris is the female equivalent of the penis of the male. It is homologous (equivalent in embryologic origin) to the penis and has a similar basic structure. It is attached by two roots and has a body composed of erectile tissue and a glans that is extensively supplied with sensory nerve endings. The labia (lips) form the external boundary of the vulva.
TEST YOURSELF 19-6
1. When ovulation occurs, what causes the ovum to enter the oviduct?
2. Describe the functions of the uterus relating to pregnancy and parturition.
3. Where is the urethral opening located in the female?
THE ESTROUS CYCLE
To get the ovum and some lucky spermatozoon together at the right time (when both are mature) in the right place (the oviduct), some intricate coordination has to take place between two different animals. The situation in the male is pretty straightforward. Spermatozoa are constantly produced in the testes, and the testosterone level stays fairly constant. So the male is basically always ready for breeding.
BOX 19-1 Estrous versus Estrus
Estrous is an adjective used with the noun "cycle" to refer to the entire reproductive cycle in females.
Estrus is a noun that is the name of the stage of the estrous cycle that is commonly referred to as the "heat period" when the female is sexually receptive to the male.
He just needs the appropriate signals from the female to get the process underway. Because ovum production in the ovary of the female is not continuous but occurs in a cyclic manner, the timing of breeding is controlled by the ovarian cycle of the female.
In all common domestic animals, breeding takes place only during a definite period in each reproductive cycle, when the chance of a successful pregnancy is the highest. This period when the female is receptive to the advances of the male is known as the heat period, or estrus. (Box 19-1 explains the difference between the words “estrous” and “estrus.”) It is characterized by physical and behavioral changes that communicate the “window of opportunity” for breeding to the male. The timing of breeding is critical if pregnancy is going to result. The spermatozoa and the ovum must enter the oviduct at just the right times with respect to each other.
The estrous cycle is defined as the time from the beginning of one heat period to the beginning of the next. It is controlled by the anterior pituitary hormones, FSH and LH. FSH and LH stimulate activity in the ovaries that causes one or more reproductive cells (ova) to mature and be released. They also stimulate the production of hormones by the developing follicle (estrogens) and the corpus luteum (progestins) after ovulation. The estrogens and progestins are directly responsible for the physical and behavioral changes in the female that are associated with the estrous cycle. Different animal species have different patterns of estrous cycles, although all go through the same basic stages.
ESTROUS CYCLE INTERVALS
Animals can be classified according to how and when their estrous cycles occur during the year. Polyestrous animals, such as cattle and swine, cycle continuously throughout the year if they are not pregnant. As soon as one cycle ends, another begins. Some polyestrous animals show seasonal variations in their estrous cycles. They cycle continuously at certain times of the year and not at all at others. These animals, such as horses, sheep, and cats, are called seasonally polyestrous. Diestrous animals, such as dogs, have two cycles per year, usually in the spring and fall. Monoestrous animals, such as foxes and mink, usually have only one cycle each year.
STAGES OF THE ESTROUS CYCLE
Although it is a continuous process, the estrous cycle can be divided into a series of characteristic stages. These stages reflect what is going on in the ovary as follicles develop, mature, rupture, and are replaced by corpora lutea. Keeping the events of the ovarian cycle in mind makes it easier to correlate what is going on in the ovary with the stages of the estrous cycle. The estrous cycle stages are proestrus, estrus, metestrus, and diestrus. (Another stage, anestrus, occurs in some animals between breeding seasons.)
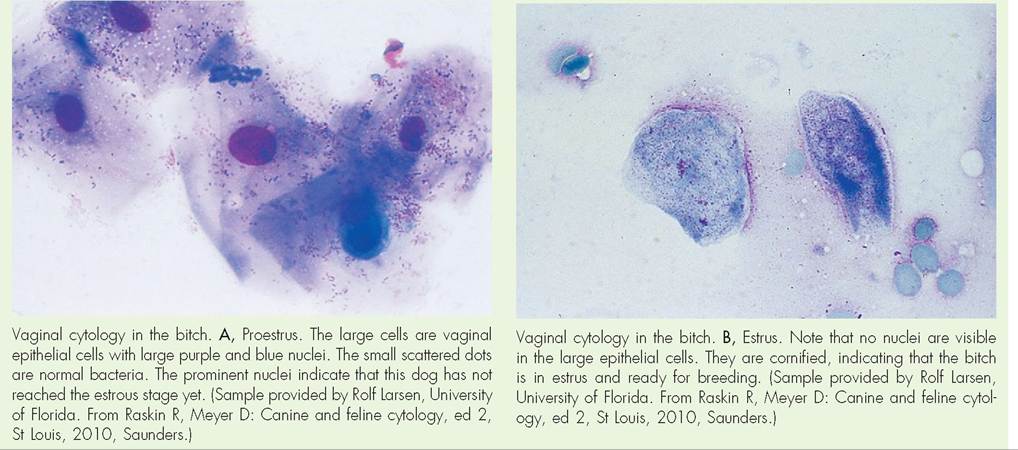
Proestrus is the period of follicular development in the ovary. During this stage, follicles begin developing and growing. As they increase in size, the follicles' output of estrogen increases accordingly, causing many physical changes that prepare the rest of the reproductive tract for ovulation and breeding. These include thickening and development of the linings of the oviduct, uterus, and vagina. The epithelial lining of the vagina also begins undergoing cornification; that is, forming a layer of tough keratin on its surface, to help protect against the physical trauma of breeding that is about to occur. These changes in the epithelial lining of the vagina can be conveniently used to detect the stage of the estrous cycle, particularly in dogs. See the Clinical Application on Vaginal Cytology.
Estrus is the heat period, or the period of sexual receptivity in the female. It occurs when the estrogen level from the mature follicles reaches its peak. This high estrogen level causes physical and behavioral changes that signal the female's willingness to breed to the male. In most species, ovulation occurs near the end of estrus. In some species, however, such as camels, cats, ferrets, and rabbits, ovulation does not occur until the animal is bred. These species, called induced ovulators, remain in a prolonged state of estrus if they are not bred. The behavioral signs of estrus in the cat— particularly the vocalizing, rolling, and rubbing—can be very annoying to owners; they may even be misinterpreted as signs of illness by novice owners.
Metestrus is the period after ovulation, when the corpus luteum develops. The granulosa cells left in the now-empty follicle begin to multiply under continued stimulation from LH. They soon produce a solid structure, the corpus luteum (yellow body), which is about the same size as the former mature follicle. The hormone progesterone, which is produced by the corpus luteum, temporarily inhibits follicular development in the ovary, causes the lining of the uterus to get very thick and “juicy” in preparation for implantation of a fertilized ovum, and causes loss of the cornified epithelial lining that developed in the vagina during proestrus and estrus. In the dog and cat there is little distinction between the metestrus and diestrus stages, so the metestrus stage is usually excluded for these species. Their estrous cycles go directly from the estrus stage to the diestrus stage.
Diestrus is the active luteal stage, when the corpus luteum has reached maximum size and exerts its maximum effect. If the animal is bred and becomes pregnant, the corpus luteum receives an endocrine signal from the developing embryo and is retained well into the pregnancy. If the animal is not pregnant, the corpus luteum degenerates at the end of diestrus. Depending on the species and the type of estrous cycle it has, the animal then either goes straight back into proestrus, or the ovary shuts down and the animal goes into anestrus. Some animals, particularly bitches, can have an exaggerated diestrous period, resulting in pseudocyesis, or what is commonly called pseudopregnancy (false pregnancy). Affected animals may act and look pregnant. The mammary glands often enlarge, the pelvis may relax, and the animal often shows maternal behavior patterns, such as nest building. In extreme cases, lactation and signs of labor may occur. Most cases of pseudopregnancy resolve spontaneously, but hormonal therapy may be required in some severe cases.
Anestrus is a period of temporary ovarian inactivity seen in seasonally polyestrous, diestrous, and monoestrous animals. It is the period between breeding cycles, when the ovary essentially shuts down temporarily. The anestrous period can be cited to dog and cat owners who are reluctant to have their animals spayed because they are afraid the animals will become fat and lazy after the procedure. In reality, dogs and cats are functionally “spayed” for a good portion of each year while they are in anestrus. Animals become fat and lazy if they are fed too much and exercised too little, regardless of whether or not they have been spayed.
TEST YOURSELF 19-7
1. What is the difference in the estrous cycle intervals of polyestrous, seasonally polyestrous, diestrous, and monoestrous animals?
2. How do the stages of the estrous cycle relate to the physical events of the ovarian cycle?
∕j CLINICAL APPLICATION
Vaginal Cytology in the Dog
Changes in the epithelial lining of the vagina during proestrus and estrus can be utilized to determine the most effective time to breed a bitch, or the most likely time she must be kept away from males to prevent unplanned breeding. The basic principle is that the vaginal lining becomes progressively thicker and the surface epithelial cells become increasingly cornified (keratinized) during the proestrus period, and this reaches a peak when the animal is in estrus and most likely to conceive if bred.
The most common technique is to insert a saline-moistened cotton swab into the vagina while avoiding contact with the lining of the vulva. The swab is gently twirled or moved back and forth, removed, and rolled onto a glass microscope slide in several rows. Then any of several different stains are applied to make the epithelial cells more visible under the microscope.
∕ j CLINICAL APPLICATION—cont'd