ANATOMIC FEATURES
The laboratory mouse has several unique characteristics, and there are vast differences in normal anatomy, physiology, and behavior among different strains of mice, many of which represent abnormalities arising from homozygosity of recessive or mutant traits in inbred mice.
Integumentary System
The history of the laboratory mouse is steeped in selective breeding for variation in coat color and consistency, with many defined mutants. Hair growth occurs in cyclic waves, beginning cranially and progressing caudally. Examination of mouse skin mandates awareness of the growth cycle and location examined. Melanin pigment is restricted to the hair follicular epithelium and hair shaft, with minimal pigmentation of the interfol- licular epidermis. Thus, newborn mice, regardless of their ultimate coat color, are uniformly pink until hair growth begins.
Hematology and Hematopoeitic System
Mouse hematology has been recently reviewed (Everds 2007). Strain-specific data and comparisons among inbred mouse strains are available through the Mouse Phenome Database. Recommended approaches to
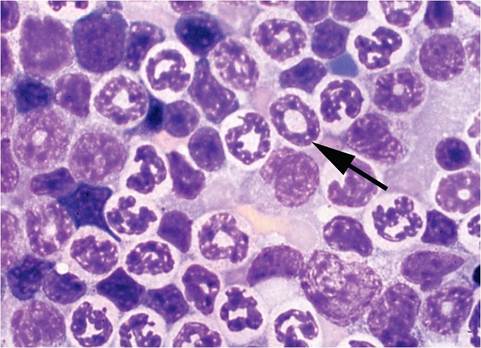
FIG. 1.1. Ring-shaped nuclei (arrow) of myeloid progenitor cells in the bone marrow of a normal mouse.
evaluation of GEMs with hematological phenotypes are also available (Car and Eng 2001). Mouse erythrocytes are small, with a high reticulocyte count, moderate polychromasia, and anisocytosis. Lymphocytes are the predominant circulating leukocyte and constitute approximately three-fourths of the total differential count. Mature male mice have significantly higher granulocyte counts than do female mice. Peripheral blood granulocytes tend to be hypersegmented, and band cells are rare, except when mice have chronic suppurative infections. Granulocytes in tissues and bone marrow often have ring-shaped nuclei (Fig.
1.1). Ring-shaped nuclei can be visualized as early as the progranulocyte stage in bone marrow, spleen, and liver, and only rarely can be found in peripheral blood. They also occur in cells of the monocytic lineage. Mice have circulating basophils, but they are extremely rare. Mice possess a very large platelet mass, due to high platelet numbers and relatively low mean volume, although some platelets can be as large as erythrocytes. The spleen is a major hematopoietic organ throughout life in the mouse, and hematopoiesis is found in the liver up to weaning age but may return in adults during disease states. Hepatic hematopoiesis can be misconstrued as inflammation. Hematopoiesis remains active in long bones throughout life.Respiratory System
Cross sections of the nose reveal prominent vomeronasal organs, which are important in pheromone sensing and are frequent targets of viral attack. Virus- associated vomeronasal and olfactory rhinitis in neonatal mice can result in failure to suckle. Respiratory epithelium may contain eosinophilic secretory inclusions (hyalinosis), which are especially obvious in B6 and 129 mice. The lungs have a single left lobe and 4 right lobes. Cartilage surrounds only the extrapulmo- nary airways in mice, rats, and hamsters. Thus, primary bronchi are extrapulmonary. Respiratory bronchioles are short or nonexistent. Cardiac muscle surrounds major branches of pulmonary veins and should not be misconstrued as medial hypertrophy. Bronchus-associated lymphoid tissue is normally present only at the hilus of the lung, except in hamsters. Lymphoid accumulations are present on the visceral pleura of mice, within interlobar clefts. These are organized lymphoid structures that are contiguous with the underlying lung tissue and are similar to "milkspots" in the peritoneum. Although not a normal finding, focal intra-alveolar hemorrhage is a consistent agonal finding in lungs of mice, regardless of the means of euthanasia. As in other species, focal subpleural accumulation of alveolar macrophages (alveolar histiocytosis) is common (see Rat chapter 2, "alveolar histiocytosis").
Gastrointestinal System
Mice are coprophagic, with approximately one-third of their dietary intake being feces. Stomach contents will reflect this behavior. Incisive foramina, located posterior to the upper incisors, communicate between the roof of the mouth and the anterior nasal cavity. Incisors grow continuously, but cheek teeth are rooted. Mice have no deciduous teeth, and their incisors are pigmented due to deposition of iron beneath the enamel layer. One of several sexual dimorphisms in the mouse is found in the salivary glands. The submandibular salivary glands in sexually mature males are nearly twice the size as females and parotid salivary glands are also larger. Male submandibular glands have increased secretory granules in the cytoplasm of serous cells (Fig. 1.2). These glands undergo similar masculinization in pregnant and lactating females. The intestine is simple. Gut-associated lymphoid tissue (Peyer's patches) is present in both the
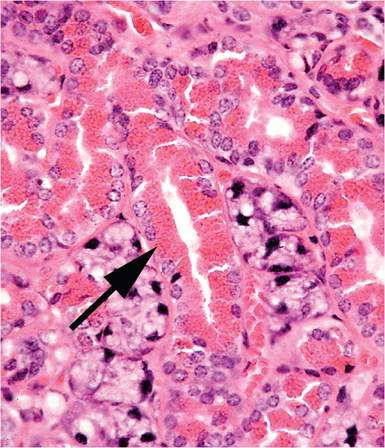
FIG. 1.2. Submandibular (submaxillary) salivary gland from an adult male mouse. Note the prominent secretory granules (arrow) in the cytoplasm of epithelial cells.
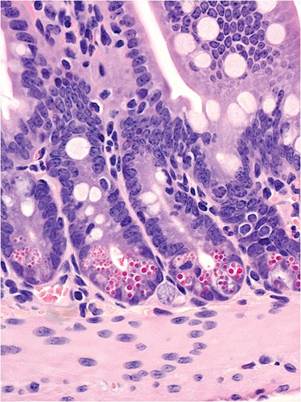
FIG. 1.3. Ileal mucosa of a mouse illustrating the distinct cytoplasmic granules within Paneth cells at the base of the crypts.
small and large intestine. Paneth cells occupy crypt bases in the small intestine. These specialized enterocytes have prominent eosinophilic cytoplasmic granules (Fig. 1.3), which are larger in mice than in other laboratory rodents. Pregnant and lactating mice have noticeably thickened bowel walls due to physiological mucosal hyperplasia. Mice have a very short (1-2 mm) rectum, which is the terminal portion of the large bowel that is not enveloped in serosa. Because of this feature, mice are prone to rectal prolapse, especially if they have colitis.
The intestine of neonatal mice has several unique features. Neonatal small intestinal enterocytes are vacuolated and may contain eosinophilic inclusions due to the presence of the apical-tubular system, which is involved in uptake of macromolecules (Fig. 1.4). It
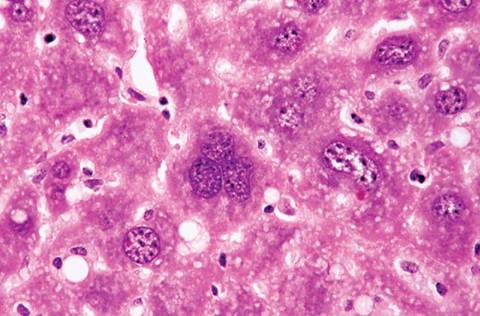
FIG. 1.5. Polykarya and megalokarya, indicative of polyploidy, are commonly found in the liver and increase with age and disease states.
disappears as the intestine undergoes maturation. The neonatal mouse bowel has very shallow crypts of Leiberkuhn populated with mitotically inactive stem cells and very long villi that are populated with terminally differentiated, absorptive epithelium. Intestinal cell turnover kinetics are slow in the neonate, making neonates highly vulnerable to acute cytolytic viruses. Turnover kinetics accelerate with acquisition of microflora and dietary stimuli.
The liver of mice has variable lobation. Polyploidy is common in mouse liver cells. Hepatocytes frequently display cytomegaly, anisokaryosis, polykarya, and kar- yomegaly (Fig. 1.5). Cytoplasmic invagination into the nucleus is frequent, giving the appearance of nuclear inclusion bodies (Fig. 1.6). Hematopoiesis normally occurs in the infant liver (Fig. 1.7) but wanes by weaning age, although islands of myelopoiesis or erythropoiesis can be found in hepatic sinusoids of older mice,
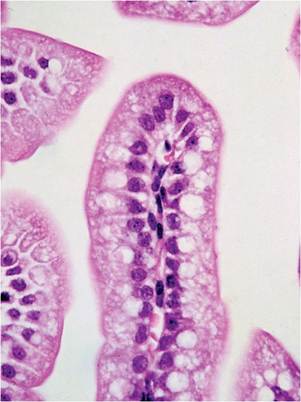
FIG. 1.4. Enteric mucosa of a neonatal mouse, illustrating the vacuolated appearance of villus enterocytes.
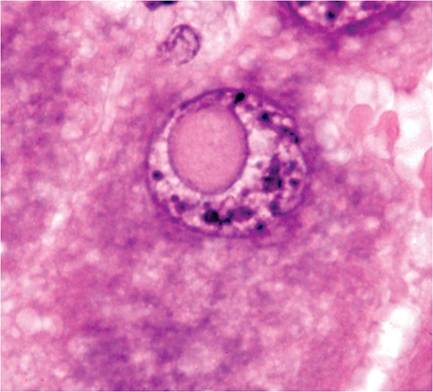
FIG. 1.6. Cytoplasmic invagination into the nucleus of a hepatocyte, a common finding in rodents that has been misinterpreted as viral inclusions.
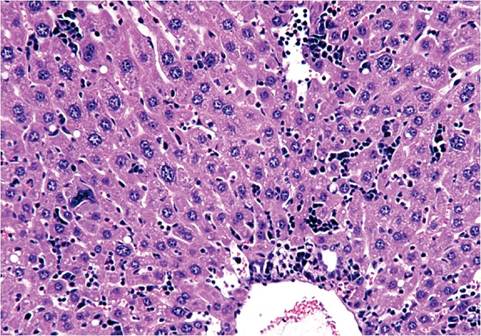
FIG.
1.7. Liver from a newborn mouse. There are numerous hematopoietic cells in the sinusoidal regions.particularly in disease states (Fig. 1.8). Hepatocytes frequently contain cytoplasmic fat vacuoles. Some strains, such as BALB mice, normally have diffuse hepatocellular fatty change, resulting in grossly pallid livers, compared with the mahogany-colored livers of other mouse strains.
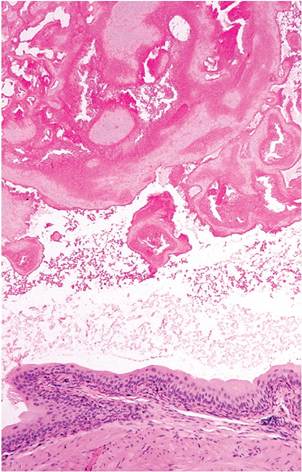
FIG. 1.9. Copulatory plug in the urinary bladder of a male mouse. The presence of ejaculate coagulum is common in the urethra and bladder as an agonal finding, although antemortem ejaculation may result in urinary obstruction.
Genitourinary System
Female mice have a large clitoris, or genital papillus, with the urethral opening near its tip, which is located anterior to the vaginal orifice. Females that develop in utero between male fetuses are somewhat masculinized, reflected by an increased anogenital distance and behavior. Tissues of the adult uterine wall are normally infiltrated with eosinophils, which wax and wane cyclically and disappear during pregnancy. Eosinophils increase in number in response to semen. Mice have hemochorial placentation. Males have large redundant testes that readily retract into the abdominal cavity through open inguinal canals, particularly when the mice are picked up by the tail. Both sexes have well-developed preputial (or clitoral) glands, and males have
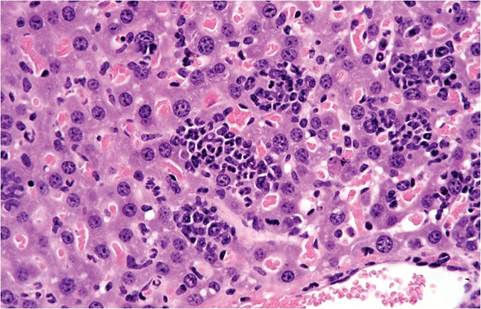
FIG. 1.8. Liver from an adult mouse with suppurative pyelonephritis, illustrating marked hepatic myelopoiesis.
conspicuous accessory sex glands, including large seminal vesicles, coagulating glands, and prostate. Ejaculation results in formation of a coagulum, or copulatory plug. This frequently occurs agonally. Coagulum can be found in urinary bladder or urethra as a normal incidental finding at necropsy (Fig.
1.9) and must not be misconstrued as a calculus or obstruction. However, copulatory plugs can and do cause obstructive uropathy. Sexual maturity in males results in several sexual dimorphic features, including larger kidneys, larger renal cortices, larger cells in proximal convoluted tubules, larger renal corpuscles, and cuboidal epithelium lining the parietal layer of Bowman's capsule, resembling tubular epithelium (Fig. 1.10). This is not absolute, since some glomeruli of male mice are surrounded by flat epithelium and some glomeruli of female mice are surrounded by cuboidal epithelium. Mice are endowed with relatively large numbers of glomeruli per unit area, compared with other species, such as the rat. Mice have a single, long renal papilla that extends into the upper ureter. Proteinuria is also normal in mice, with highest levels in sexually mature male mice. Major contributors to proteinuria in male mice are “mouse urinary proteins,” which function as pheromones. In particular, MUP-1 is highly antigenic and a major cause of occupational allergies among animal handlers.Endocrine System
The mouse adrenal gland has several notable features. The adrenals of male mice tend to be smaller and have less lipid than those of females. Accessory adrenals, either partial or complete, are very common in the
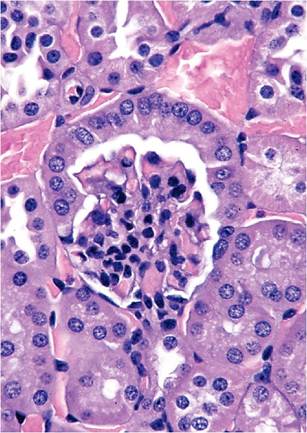
FIG. 1.10. Renal cortex from an adult male mouse, illustrating the cuboidal epithelium lining the parietal surface of Bowman's capsule.
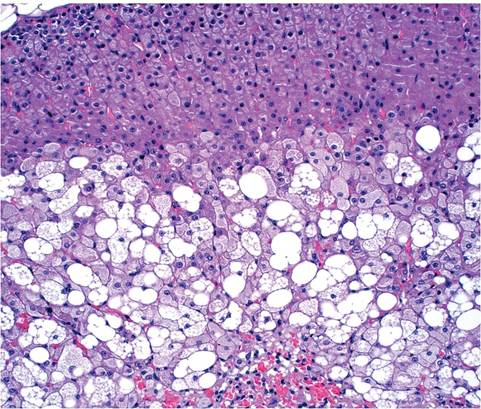
FIG. 1.12. Vacuolating degeneration of the involuting X zone at the corticomedullary junction of the adrenal gland of an adult female mouse.
adrenal capsule or surrounding connective tissue. The zona reticularis of the adrenal cortex is not discernible from the zona fasciculata. Proliferation of subcortical spindle cells, with displacement of the cortex, is common in mice of all ages (Fig. 1.11). The function of these cells is not known. A unique feature of the mouse adrenal is the X zone of the cortex, which surrounds the medulla. The X zone is composed of basophilic cells and appears in mice around 10 days of age. When males reach sexual maturity and females undergo their first pregnancy, the X zone disappears. The X zone disappears gradually in virgin females. During involution, the X zone undergoes marked vacuolation in females
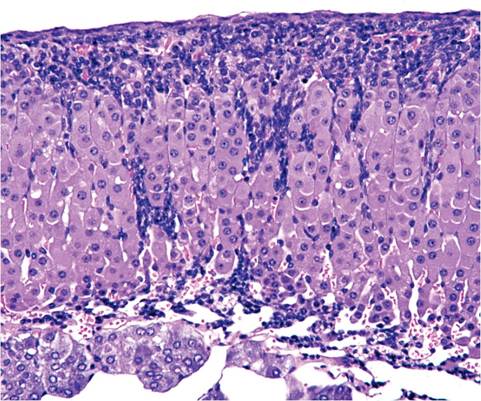
FIG. 1.11. Subcapsular spindle cell proliferation in the adrenal gland of a normal adult mouse. This change is common, but its significance is not known.
(Fig. 1.12) but not in males. Residual cells accumulate ceroid. Pancreatic islets are highly variable in size, including giant islets that can be confused with hyperplasia or adenomas.
Skeletal System
Bones of mice, like those of rats and hamsters, do not have Haversian systems, and ossification of physeal plates with age is variable and incomplete, depending upon mouse genotype.
Lymphoid System
Rodents do not have tonsils, but have nasal-associated lymphoid tissue (NALT). Germinal centers are not well defined in lymph nodes. The thymus does not involute in adults. Hassall's corpuscles are indistinct. Islands of ectopic parathyroid tissue may be encountered in the septal or surface connective tissue of the thymus, and, conversely, thymic tissue may occur in thyroid and parathyroid glands. Epithelial-lined cysts are also common. The splenic red pulp is an active hematopoietic site throughout life (Fig. 1.13). During disease states and pregnancy, increased hematopoiesis can result in splenomegaly. Lymphocytes tend to accumulate around renal interlobular arteries, salivary gland ducts, urinary bladder submucosa, and other sites, increasing with age. These sites are often involved in generalized lymphoproliferative disorders. Melanosis of the splenic capsule and trabeculae is common in melanotic strains of mice (Fig. 1.14). This must be differentiated from iron (hemosiderin) pigment (Fig. 1.15), which tends to accumulate in the red pulp as mice age, particularly in multiparous females. Mast cells can be frequent in the spleen of some mouse strains, such as A strain mice.
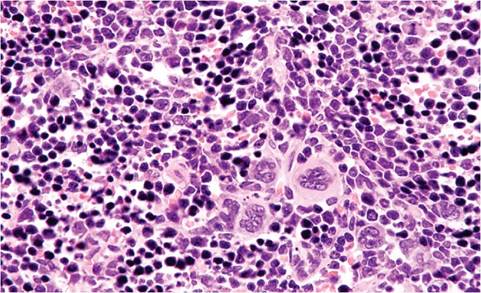
FIG. 1.13. Spleen from an adult mouse, illustrating the large numbers of hematopoietic cells, including megakaryocytes, in the sinusoids, a common finding throughout life.
Other Anatomic Features
The brain and spinal cord are larger in mature male mice compared to females. Melanosis occurs in the antero- ventral meninges of the olfactory bulbs, optic nerves, parathyroid glands, heart valves, and spleens of melanotic mouse strains, such as B6 mice. Foci of cartilage or bone can be found within the base of the aorta. These foci are not an os cordis but rather occur within the wall of the aorta. Mice have 3 pectoral and 2 inguinal pairs of mammary glands, with mammary tissue enveloping much of the subcutis, including the neck. Mammary tissue can be found immediately adjacent to salivary glands, which is especially apparent during lactation. Nipple development is hormonally regulated in mice, and nipples are quite small in males. Mammary tissue of males totally involutes during development. Remarkably, virgin female mice can be induced to lactate by the presence of other females nursing litters. Mammary glands normally involute between pregnancies, but they do not involute in multiparous FVB mice, due to a tendency to develop hyperplasia of prolactin-producing cells and pituitary adenomas. Brown fat is prominent as a subcutaneous fat pad over the shoulders and is also present in the neck, axillae, and peritoneal tissue.
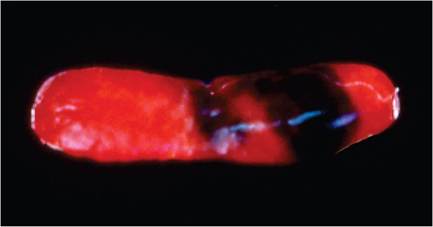
FIG. 1.14. Splenic melanosis in a melantotic (C57BL) mouse. Note the patches of pigmented capsule.
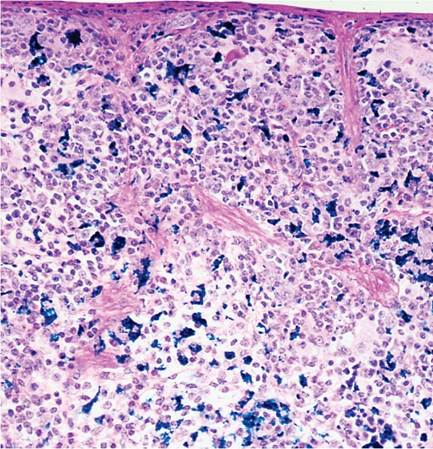
FIG. 1.15. Iron pigment (hemosiderin) in the spleen of an adult female mouse (Perl's stain).
Immunologic Idiosyncracies
Neonatal mice are globally immunodeficient. Different components of the innate and acquired immune response subsequently evolve at differing rates, depending upon genetic background. Although mice are generally immunocompetent at weaning, they are not fully so until 6-12 weeks of age. Neonates depend upon acquisition of maternal antibody to protect them during early life. Maternal IgG is transferred in utero through Fc yolk sac receptors, and postnatally through IgG receptors in the small intestine, which actively acquire immunoglobulin up to 2 weeks of age. Milk-borne IgA is also important in protecting suckling mice, but neither IgA nor IgM are absorbed. Passive immunity is a critical component in understanding the outcome of viral infections in mouse populations. Epizootic infections can be devastating in naive populations of neonates, but once the infection becomes enzootic within a population, maternal antibody protects suckling mice during their period of age-related vulnerability. Maternal antibody generally persists in the serum of pups for about 6 weeks.
The immune response can vary considerably among different strains of mice. An often-cited feature is the Th1-Th2 polarized T-cell response, in which BALB/c mice tend to respond to antigenic stimuli with a Th2 skewed response and B6 mice with Th1 skewed responses. This is far from absolute, but there seems to be truth in the concept that B6 mice deal more efficiently with viral infections. B6, B10, SJL, and NOD mice have their own unique immunoglobulin isotype, IgG2c, in lieu of, but distinct from, IgG2a. IgG2c is not an allelic variant of IgG2a, since in these strains the IgG2a gene is completely absent, and in IgG2a-positive strains, the IgG2c gene is absent. This may impact accurate measurements of humoral responses. The mouse genome possesses approximately 40 histocompatibility loci, and the major histocompatibility (MHC) loci are located on chromosome 17 within the MHC complex, known as the H-2 complex. Each inbred strain of mouse has a defined H-2 haplotype, or combinations of alleles, which are well-recognized determinants of strain-specific immune responses, including responses to infectious disease. Because of the inbred nature of laboratory mouse strains, H-2 haplotype is a singularly important strain characteristic.
Various stressors, including dehydration, hypothermia, and acute infections, may result in massive corticosteroid-induced lymphocytic apoptosis. This is accompanied by generalized lymphoid depletion and transient nonspecific alterations of immune responsiveness. This is especially apparent in the thymus and is a frequent and rapid onset lesion in “water bottle accidents,” when mice become hypothermic or dehydrated. Recently rederived and xenobiotic mice have lymphoid hypoplasia, accompanied by functional hyporesponsiveness.
Genetic engineering has given rise to many immunologic mutants of mice, and other naturally arising immune mutants have also been popularized, such as nude (T-cell-deficient), SCID (B- and T-cell-deficient), and beige (NK cell-deficient) mice. The preeminent immunodeficient mouse is the NSG mouse, discussed above. Immunodeficient mice must never be considered to be simply missing a single functional component of the immune system, since they typically have compen- satorily activated innate and acquired immune responses compared to wild-type. Homozygous immunodeficient inbred mouse mutants that are progeny of the heterozygous (immunocompetent) parental matings or through embryo transfer into immunocompetent recipients can acquire functional immunoglobulin- secreting B cells from their immunocompetent dams. They can also acquire functional B cells postnatally through foster nursing. The chimeric cells may remain functional for at least several months.
Less obvious and often overlooked immunologic idiosyncrasies also exist among common inbred strains. All strains of adult male mice manifest a sexual dimorphism in which serum levels of both C4 and C5 are higher than in females, and male SJL mice have a significantly higher level of C5 compared to males of other strains. In addition, inadvertent consequences have arisen from inbreeding and selection for other characteristics. One such common defect is a 2 base pair gene deletion in the 5th component of complement (C5). This mutation results in C5 deficiency in many inbred strains of mice, including AKR, SWR, DBA/2J, A/J, A/HeJ, NOD, and RF, among others. SJL mice are NK cell-deficient. NOD mice have multiple immune defects (see above). Substrain divergence due to spontaneous acquisition of mutations can give rise to novel new substrain phenotypes, such as the LPS unresponsiveness of C3H/HeJ and C57BL/10ScN mice, which is attributed to a mutation of toll-like receptor 4 (TLR4). All strains of mice lack functional TLR10 due to genetic disruption by a retroviral insertion. CBA/CaN (CBA/N), but not other CBA mice, have an X-linked defect in humoral immunity, with impaired maturation of B cells, diminished immunoglobulin production, and impaired T-independent immune responses. Thus, knowledge of specific strain and substrain characteristics greatly improves the understanding of responses to experimental variables.