AUJESZKY’S DISEASE, OR PSEUDORABIES
FRANCISCO RUIZ-FONS1
1Instituto de Investigacion en Recursos Cinegeticos IREC (CSIC-UCLM-JCCM), Ciudad Real, Spain
Aujeszky’s disease (AD), pseudorabies or ‘mad itch’ is a neurological/respiratory disorder that affects a wide range of animals, except humans and some primates.
It is caused by porcine or suid herpesvirus type 1, also known as pseudorabies virus or Aujeszky’s disease virus (ADV), which belongs to the family Herpesviridae in the genus Varicellovirus.AETIOLOGY
ADV is a 150—180 nm virion composed of a 145 Kb linear double-stranded DNA genome within an enveloped
TABLE 1.1 Important mammalian herpesviruses for European wildlife. Viruses are presented according to their taxonomic distribution within the three subfamilies of the order Herpesvirales.
| Name | Acronym | Common name | |
| Subfamily Genus | Alphaherpesvirinae Simplexvirus | ||
| Species in the genus | Bovine herpesvirus 2 | BoHV2 | Bovine mammilitis virus |
| Human herpesvirus 1 | HHV1 | Herpes simplex virus type 1 | |
| Genus | Macacine herpesvirus 1 Varicellovirus | McHV1 | Herpes simian B-virus |
| Species in the genus | Bovine herpesvirus 1 | BoHV1 | Infectious bovine rhinotracheitis virus |
| Bubaline herpesvirus 1 | BuHV1 | Water buffalo herpesvirus*a | |
| Canid herpesvirus 1 | CaHV1 | Canine herpesvirus | |
| Caprine herpesvirus 1 | CpHV1 | Goat herpesvirus | |
| Cervid herpesvirus 1 | CvHV1 | Red deer herpesvirus | |
| Cervid herpesvirus 2 | CvHV2 | Reindeer herpesvirus | |
| Felid herpesvirus 1 | FeHV1 | Feline rhinotracheitis virus | |
| Phocid herpesvirus 1 | PhoHV1 | Harbour seal herpesvirus | |
| Suid herpesvirus 1 | SuHV1 | Pseudorabies virus | |
| Unclassified in the subfamily | n/a | n/a | Bottlenose dolphin herpesvirus |
| Subfamily Genus | n/a Betaherpesvirinae Cytomegalo virus | n/a | Tursiops truncatus alphaherpesvirus |
| Species in the genus | Macacine herpesvirus 3 | McHV3 | Rhesus macaques cytomegalovirus*b |
| Unclassified in the subfamily | Suid herpesvirus 2 | SuHV2 | Porcine cytomegalovirus |
| Subfamily Genus | n/a Gammaherpesvirinae Lymphocryptovirus | n/a | Bat betaherpesvirus |
| Genus | Human herpesvirus 4 Macavirus | HHV4 | Epstein-Barr virus*c |
| Alcelaphine herpesvirus 1 | AlHV1 | Malignant catarrhal fever virus*d | |
| Alcelaphine herpesvirus 2 | AlHV2 | Hartebeest malignant catarrhal fever virus*d | |
| Caprine herpesvirus 2 | CpHV2 | Caprine herpesvirus 2 | |
| Genus | Ovine herpesvirus 2 Percavirus | OvHV2 | Sheep-associated malignant catarrhal fever virus |
| Mustelid herpesvirus 1 | MusHV1 | Badger herpesvirus | |
| Unclassified in the genus Genus | Phocid herpesvirus 2 Rhadinovirus | PhoHV2 | Phocid herpesvirus 2 |
| Unclassified in the genus | Leporid herpesvirus 2 | LeHV2 | Herpesvirus cuniculi |
| Unclassified in the subfamily | n/a | n/a | Rupicapra rupicapra gammaherpesvirus 1 |
| Unclassified in the family | Erinaceid herpesvirus 1 | ErHV1 | European hedgehog herpesvirus |
| Sciurid herpesvirus 1 | ScHV1 | Ground squirrel cytomegalovirus | |
| Sciurid herpesvirus 2 | ScHV2 | Ground squirrel herpesvirus |
n/a — not available
*a Most buffalos in Europe are semi-domesticated
*b Mostly only at zoos, only monkey wild population in Europe living in Gibraltar is free from McHV3 *c Shown experimentally to infect dog cells and also found in seroscreenings of canids
*dPresent in Europe only in zoos but represent the type species in the genus
nucleocapsid.
The 105—110 nm wide nucleocapsid is formed by different structural proteins and its envelope is a lipidic membrane composed of nine different enclosed glycoproteins used in the life cycle of the virus, immune modulation and pathogenicity.EPIDEMIOLOGY
ADV is widely distributed in European wild boar populations (Figure 1.1)(3). Some countries, where AD has not been identified in wild boar populations, have reported
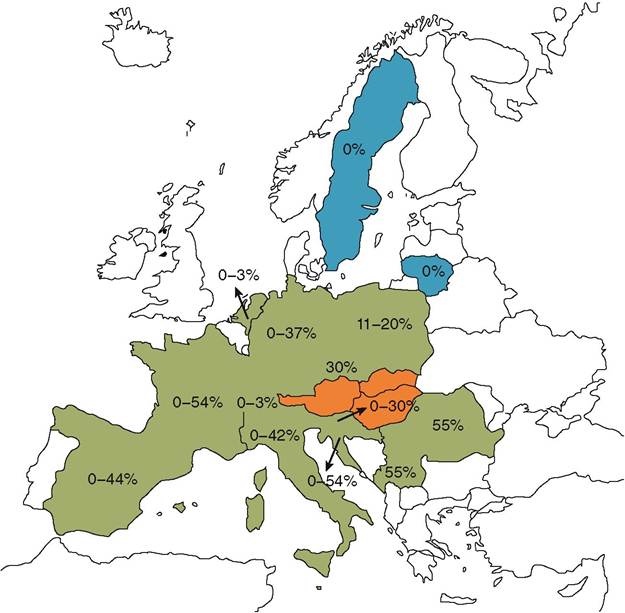
FIGURE 1.1 European countries where reports of ADV surveillance in wild boar populations have been published in the scientific literature® or reported in national wildlife surveillance programmes between 1987 and 2011. Countries shaded in green represent ADV-positive wild boar populations, whereas countries shaded in blue represent ADV-negative surveyed wild boar populations. Countries shaded in orange have reported pseudorabies outbreaks in hunting dogs associated with wild boar hunting, but where the status of wild boar populations is unknown. Seroprevalence/ prevalence reported range of the within-country surveyed European wild boar populations are shown.
pseudorabies outbreaks in dogs used in boar hunting, i.e. Austria1-4’5), Belgium)6), Hungary and Slovakia)7). Several European countries where ADV has not been reported or where it was eradicated from domestic pigs have not assessed the status of ADV in their wild boar populations (e.g. Denmark, Norway, Finland or the UK). Thus, the current known distribution of ADV in European wild boar populations may not be accurate.
ADV is able to infect a wide range of mammals, including ungulates, carnivores, lagomorphs, rats and mice. Infection in mammals is usually fatal; however, in some species subclinical infection is possible)8). In suids, the only natural host species for ADV, the infection may cause disease or be subclinical.
Many European wild boar populations have had laboratory assessments for the presence of ADV or anti-ADV antibodies (Figure 1.1); however, the basic understanding of ADV epidemiology in boar is poor. Pan-European serological studies on ADV in wild boar have shown that the probability of contact with the virus increases with age. ADV causes life-long latent infection in suids and naturally infected animals remain seropositive, and potentially infective, for life. A similar viral exposure risk occurs for males and females; however, sex-related differences, with higher exposure of females to ADV, is seen in some European and North African wild boar populations (9). This may be related to behavioural differences between the sexes. Intra-group transmission is higher in all-female groups of wild boar, whereas males tend to be solitary. The probability of wild boar acquiring ADV in endemic areas also seems to be dependent on population density and the extent to which the animals aggregate)10), both of which are highly variable factors across Europe, and this gives rise to regional/local variations in prevalence. Additionally, wild boar population structure, female group size, management or predation may influence the rate of transmission of ADV within and between groups. This could be the reason for the similar viral infection risk of males and females observed in many wild boar populations in Europe. Movement of individuals between infected and susceptible wild boar groups or populations is likely to be important for virus spread.
ADV survival rate in the environment is low. Transmission by the aerosol route is also low in hot and dry weather conditions, which are unfavourable for the virus, but is enhanced if weather conditions are cool and wet.
The European wild boar is currently considered as a true ADV reservoir, because the virus can infect, replicate and be excreted in this species, which is sufficiently abundant to be a wild reservoir. Other mammalian species are deadend hosts in which death occurs before viral excretion.
In the USA, some experimentally infected raccoons (Procyon lotor) have been found to behave as short-term reservoirs ofADV when infected at low doses(8), which would suggest a transient reservoir role.Currently the main routes of ADV transmission in the European wild boar are not known; however, they are suspected to be by direct contact between individuals. There is little information as to whether aerosol infection is an efficient transmission pathway between wild boar. The oronasal route is suspected to be the usual means of ADV transmission between European wild boar, but the precise importance of aerosol transmission even over short distances is not known.
Venereal transmission is considered of primary importance for ADV transmission in American feral pig popula- tions(11), and it may be an important route in European wild boar as well. An increase in seroprevalence after the mating season was found in wild boar in Spain(9), which, apart from suggesting an increasing contact rate between individuals, may perhaps also reflect the occurrence of venereal transmission. Additionally, ingestion of infected meat via cannibalism is considered a possible route of transmission.
Wild boar females usually live in groups with their offspring and juvenile animals. This may give rise to closer contact within female groups, and oronasal transmission is thought to predominate in these groups. Wild boar males are usually solitary for most of the year except during the mating season, when they make contact with female groups. Venereal transmission could be linked with reactivation of latent infections due to mating stress. Behavioural patterns of wild boar depend to a large extent on the availability of food resources, and it is believed that these food-based behaviours may be an important influence in determining ADV prevalence. The threshold infective dose for ADV in wild boar may vary according to the virulence of the circulating strain and the immune status of the infected animal, as occurs in the domestic pig.
ADV is excreted in suids by nasal exudates, saliva, vaginal mucus, sperm, milk, faeces and occasionally urine. Different routes of infection by ADV are potentially possible because there is some, unquantified, survival of the virus in the environment, particularly in organic material, and some persistence in aerosols. Wild carnivores acquire infection after consumption of ADV- infected wild boar meat, as may happen to dogs that eat or bite infected wild boar during hunting. Direct contact with ADV-excreting boar or indirect contact with infected fomites or aerosols are assumed to be the main ways of infection for wild ungulates.
PATHOGENESIS, PATHOLOGY AND IMMUNITY
Following primary infection, viral replication of ADV takes place in the nasal or genital mucosa and in the ton- silar epithelium. Later, ADV invades the nervous system via the nerve endings present in the genital, oral and nasal mucosae and progresses by moving along the nerves into the central nervous system (CNS). At this stage of infection, ADV can be detected in oropharyngeal tonsils, nasal cavity, genital mucosa, sacral ganglia or trigeminal ganglia. At this stage the virulence of the ADV strain and the immune status of the host (in the case of true reservoirs) determine whether there is invasion of the CNS, or establishment of a latent infection in the trigeminal or sacral ganglia. Infection progresses rapidly into the CNS in deadend hosts. The virus can be detected in association with blood cells after infection but peripheral blood mononuclear cells do not carry ADV in latent infections. The virus may also replicate in lung and pharyngeal respiratory epithelia and in endothelium.
Very little is known about natural disease development in wild boar, but recent natural AD cases in wild boar piglets in Germany(12) show similarities with domestic pigs. Clinical disease in the domestic pig ranges from fatal nervous disease usually seen in piglets, respiratory problems in post-weaning pigs and respiratory and reproductive manifestations in adults.
Encephalitis has been found in wild boar naturally infected with ADV(12,13) and in animals that have been experimentally infected with ADV of moderate virulence(14). The pathological outcome of ADV infection depends on the virulence of the strain. It is hence probable that low-virulence strains present in European wild boar populations may cause no lesions in this species. The histopathological findings consist of nonsuppurative meningoencephalitis and ganglioneuritis with neuronal degeneration, focal gliosis, perivascular mononuclear cuffing and lymphocytic inflammation. Intranuclear inclusion bodies may be observed in neurons of the CNS or in ganglionar neurons. The viral tropism for epithelial tissues in the respiratory tract leads to alveolar, bronchiolar and bronchial epithelial degeneration and mononuclear cell infiltration. Degeneration and necrosis, often with intranuclear inclusion bodies, may occur in the liver, spleen, kidneys, pancreas, adrenal gland, thymus, lymph nodes, tonsils and intestinal epithelium. Oedema and haemorrhages are frequently observed.Disease in dead- end hosts progresses rapidly, usually with a fatal outcome within 24 to 72 hours following infection. The tropism of ADV for endothelial cells leads to extravasations and oedema in the lungs, nasal and oral cavities. ADV pathogenesis is broadly similar for different dead- end host species, except for mink, in which vasculopathy is predominant to neuropathy1-15). Gross and microscopic lesions of AD in dead-end hosts and domestic pigs reflect the neurotropic nature of this herpesvirus. Many of the affected dead-end hosts may show no gross lesions because of the rapidly fatal outcome of infection, or they may show skin lesions caused by self-trauma due to the intense pruritus (see the Clinical Signs section below). Fibrinoid vasculitis, with haemorrhages and myocardial necrosis, is inconsistently described but appears to be typical in farmed mink(16). Cardiac alterations in dogs may cause sudden death due to arrhythmias. Lesions in abdominal organs have been also found in different species of North American carnivores such as bears, coyotes ( Canis latrans) and a Florida panther (Puma concolor coryz).
ADV infection evokes both humoral and cell-mediated immune responses in suids, but the immune response is unable to completely clear infection, and reinfection and activation of latent infections may occur. The cellular immune response to ADV has been the subject of little research in wild boar. Outer envelope ADV glycoproteins stimulate the production of neutralising antibodies, particularly those directed against glycoproteins (g) C and D (gC and gD). During the early stages of infection, neutralizing antibodies block virus attachment and invasion of cells. Infection of wild boar with low-virulence ADV strains (those circulating in European wild boar populations) induces a long- lasting active humoral immunity, which can be passed on to the offspring and confer protection to boar piglets during the first 15 weeks of life(17).
A characteristic of herpesviruses is their ability to evade the host immune response by producing long-term latent infections in specific tissues. Subsequent immunosuppression in the host may allow the infection to reactivate with viral replication and then dissemination throughout the body. Virus can then be excreted in high titre and is able to infect other susceptible individuals. ADV mainly establishes latency in neuronal cells, such as the trigeminal or sacral nervous ganglia. Reactivation of latent infections does not usually lead to overt clinical disease. Reactivation of latent infections should be carefully considered when planning ADV eradication from domestic pigs. It should also be considered when studying ADV epidemiology in wild boar populations.
CLINICAL SIGNS
ADV strains circulating in some European wild boar populations are attenuated and as a result have low virulence. The majority of the wild boar infected with ADV show no clinical signs. Experimental infection of wild boar with virulent strains has resulted in fatal disease(14), similar to that following experimental infection in domestic pigs. Experimental infection of immune-compromised wild boar with ADV strains of wild boar origin has resulted in clinical disease(14). An outbreak of AD was reported in European wild boar in Spain, where juveniles and adults showed nervous clinical signs and the mortality was 14%(13). Two wild boar with signs of neurological disturbance have been diagnosed with AD in Germany(12). These findings indicate that clinical disease cases in free- living boar in Europe may occur but are infrequently observed.
Mild clinical signs including mild pyrexia, sneezing, nasal discharge and conjunctivitis were observed in wild boar experimentally infected with an ADV isolate from wild boar origin(14); however, following steroid-induced immunosuppression, when these animals were reinfected using the same strain, they developed severe clinical disease with pneumonia and death.
I n wild dead- end host species the clinical outcome of AD is usually fatal, resulting in death within a few days after infection. The first signs are appetite loss and diminished activity, but later the animal develops mild nervous signs. A sero-mucoid nasal discharge may appear, as well as respiratory distress and fever. The affected animals often develop pruritus, which may lead to self-mutilation. Later excitement and hyperaesthesia become greater and convulsions can occur before the animal collapses and dies. In some cases the clinical course is very short and death is rapid, with only minimal clinical signs observed.
Diagnosis
Aujeszkys disease should be considered when neurological disease is seen in European wild mammals; however, some countries, such as the UK, are free of ADV. Detection of virus is by isolation in cell cultures or molecular detection of ADV genomic material in tissues. PCR testing utilizes the glycoprotein encoding genes, which are highly conserved between different ADV strains (gB/gD) and constitute the main target of polymerase chain reaction (PCR) tests.
Viral isolation and/or viral genome detection by PCR are used for the diagnosis of ADV infection in the European wild boar. The trigeminal ganglia (TG) are considered the best site to detect latent infections in domestic pigs. The attenuated nature of European wild boar ADV strains may lead to the establishment of latent infections in sacral ganglia after venereal transmission as has been recorded in feral pigs in North America. Hence, absence of ADV in TG does not exclude latent infection in European wild boar(18). In preparation for PCR testing, both sets of ganglia require dissecting out and removal from dead animals.
Serological methods for detection of anti-ADV antibodies are of limited diagnostic use in non-suid species because of the rapid course of the infection. Viral neutralization tests, western blot and enzyme-linked immunosorbent assay (ELISA) may be useful techniques for the detection of antibodies against ADV in suids. The ELISA is a sensitive and specific test in the domestic pig. Owing to its low cost, high reproducibility and rapidity of use it is also a useful tool for epidemiological studies in European wild boar. However, 45% of European wild boar with viral ADV DNA did not have antibodies detectable with ELISA(18). As a result of suspicions that the currently used ELISA may not detect all wild boar ADV antibodies, new serological tests may be necessary in particular to identify latently infected wild boar in Europe. Further research is required in Europe to ensure that diagnostic tests used for wild boar are reliable.
MANAGEMENT, CONTROL AND REGULATIONS
Management of ADV in wild boar populations first requires surveillance for the disease. Where presence of ADV is identified, management of the disease in free- living European wild boar is difficult because: i) ADV is widely distributed across European wild boar populations; ii) its prevalence is high in some wild boar populations and seems to be increasing, while the geographical range of ADV infected boar is also extending in some regions; iii) there is little relevant information on the efficacy of preventive management strategies such as vaccination, reduction of population densities (through targeted hunting) and avoiding supplementary feeding, which results in concentration of animals(19). Risk assessment is particularly important when considering ADV control in wild boar. A limited amount of work has been done in Europe on the testing of an Aujeszkys disease vaccine for wild boar; however, currently no validated vaccine is available. There is no reporting regulation of ADV in wild boar in Europe. Aujeszkys disease is notifiable to the World Organisation for Animal Health (OIE).
PUBLIC HEALTH CONCERN
ADV is considered as a non-zoonotic pathogen, but mild pruritus may appear in humans when handling the virus in the laboratory1-20).
SIGNIFICANCE AND IMPLICATIONS FOR ANIMAL HEALTH
Aujeszkys disease is common to both wild boar and domestic pigs, and it has been eradicated from the domestic pig in many European countries. Contact between wild boar and domestic pigs, especially in extensive production, may lead to outbreaks in the domestic pig as a consequence of ADV circulating in wild boar populations.
The effect of ADV on the population dynamics of wild boar appears to be limited to reduced reproductive output and reduced survival of neonates, with little measurable effect in reducing the overall numbers of animals in boar populations. However, there is insufficient information on the disease in wild boar populations to properly assess the effects of ADV.
There are several reported incidents of Aujeszkys disease causing deaths in dogs used in boar hunting across Europe. Carnivores, including threatened species, that consume wild boar are at risk of acquiring ADV. Wolves are active predators of European wild boar and may be at a high risk of contracting Aujeszkys disease; however, there is little published evaluation of the effects of the disease on wild animals other than boar. The seroprevalence of ADV in wild boar has increased substantially in the remaining habitat for the IUCN critically endangered Iberian lynx (Lynx pardinus) in Spain, and the disease poses a risk to reintroduction programmes in these areas.