Autonomic nervous system
Overview
Many activities of an animal are under conscious control. Such activities include voluntary motor movement and generally are not immediately necessary to sustain life.
These activities are controlled by the somatic nervous system. In contrast, most physiological and endocrine activities of the body require no conscious activity but are instead controlled automatically. These activities are generally essential to sustain life and include such activities as blood pressure, heart rate maintenance, and control of blood chemistry. These life-sustaining activities are controlled by the autonomic nervous system, which coordinates and integrates the visceral functions of the body. As originally proposed by Claude Bernard and further developed by Walter Cannon, the concept of homeostasis is largely controlled by the autonomic nervous system.The somatic nervous system, or somatic motor system, is under voluntary control and regulates the contraction of skeletal muscle. In contrast, the autonomic nervous system, or visceral motor system, is generally not under voluntary control and controls visceral effectors, including smooth muscle, cardiac muscle, glandular tissue, and visceral reflexes.
There are some fundamental differences in the arrangement of these two branches of the nervous system (Fig. 10.24). Efferent fibers in the somatic nervous system begin in the primary motor cortex and travel either to cranial nerve nuclei located in the brain or to α-motor neuron cell bodies in the anterior horn of the spinal cord. From there, motor neurons exit the CNS and travel to skeletal muscles. Therefore, in the somatic nervous system, the cell bodies of the motor neurons are located within the CNS.
Efferent fibers of the autonomic nervous system generally originate in the hypothalamus and travel to either autonomic nerve nuclei in the brain or preganglionic neurons located in the anterior horn of the spinal cord.
The preganglionic neurons then leave the spinal cord and synapse on postganglionic neurons located in autonomic ganglia located in the periphery.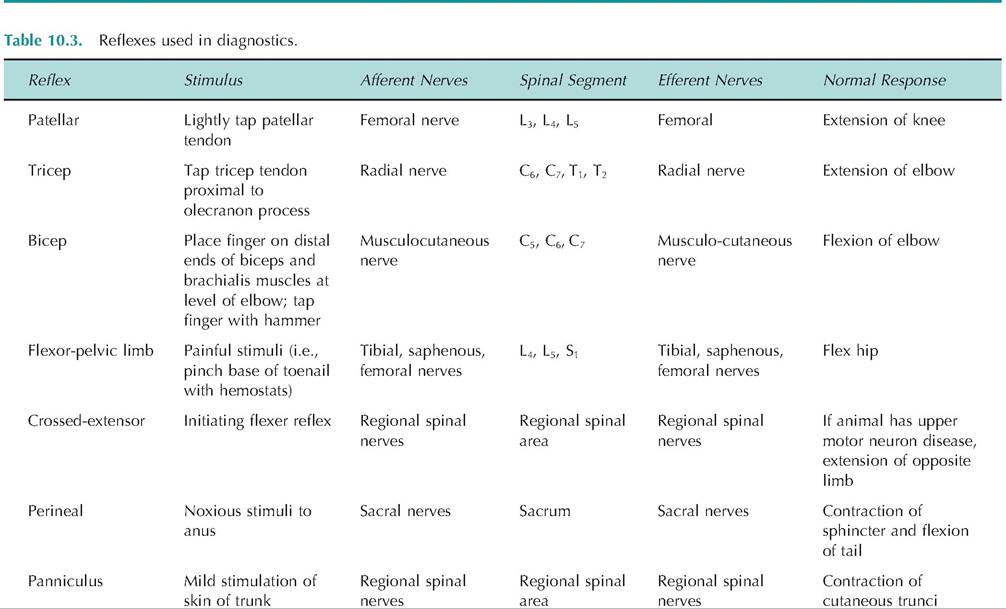
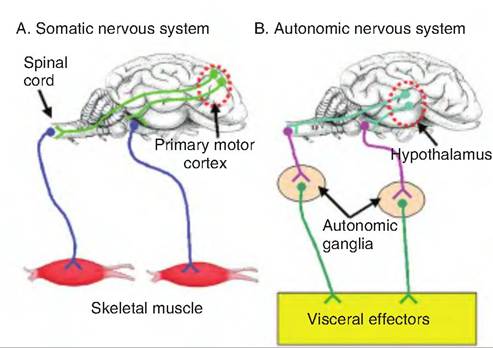
Fig. 10.24. Organization of the somatic and autonomic nervous systems. (A) Motor commands for the somatic nervous system originate in the primary motor cortex. These efferent fibers then synapse either within the brain on nuclei for cranial nerves, or on α-motor neurons within the spinal cord. The α-motor neurons, whose cell bodies are within the central nervous system, exit and travel to the skeletal muscle. (B) Motor commands for the autonomic nervous system originate in the hypothalamus. These efferent signals then travel to autonomic nuclei either in the brain or spinal cord where they synapse on preganglionic fibers. These preganglionic fibers leave the central nervous system and travel to autonomic ganglia located in the periphery where they synapse on postganglionic fibers. The postganglionic fibers travel to the visceral effectors such as smooth muscle, cardiac muscle, and glands.
These neurons are called pre- and postganglionic neurons because they synapse on one another within a ganglion located in the peripheral nervous system. In contrast to the somatic nervous system, the cell bodies of motor neurons of the autonomic nervous system are located outside of the CNS within autonomic ganglia.
Innervation of target tissue by the autonomic nervous system differs from that of the somatic nervous system. Whereas somatic neurons innervating skeletal muscle have specialized synapses called motor end plates containing specialized presynaptic structures such as an active zone, nerve endings of autonomic neurons have only swellings called varicosities and vesicles containing neurotransmitters (Fig. 10.25). Autonomic neurons can be highly branched with synaptic transmission occurring at multiple sites along an axon terminal.
Overlap of somatic and autonomic functions
Both somatic and visceral sensory information can initiate a visceral reflex (Fig. 10.26). In other words, the two systems work together to maintain homeostasis. For example, if an animal suddenly feels cold, it may activate the somatic nervous system to move to a
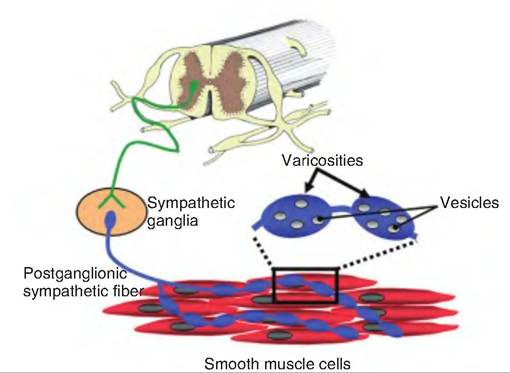
Fig- 10.25. Sympathetic varicosities. Rather than embedding within the smooth muscle cells, postganglionic sympathetic fibers running over smooth muscle cells have swellings along their length called varicosities. Within the varicosities are vesicles containing norepinephrine.
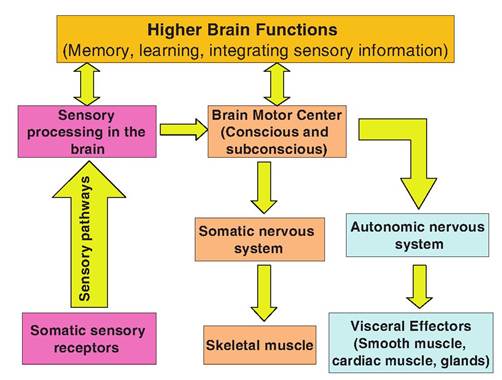
Fig. 10.26. Neural integration of the somatic and autonomic nervous systems. The somatic and autonomic nervous systems can simultaneously elicit various responses as a result of sensory information.
warmer location but may also activate a visceral reflex that causes vasoconstriction in the skin and piloerec- tion in order to reduce heat loss. If an animal is running hard, not only are neural inputs necessary to tell the skeletal muscles to contract, but visceral reflexes are necessary in order to increase blood flow to the skeletal muscles.
Divisions of the autonomic nervous system
The autonomic nervous system is subdivided into the sympathetic and parasympathetic subdivisions. These subdivisions have opposing effects on most functions. The sympathetic division, sometimes called the "flight- or-fight" division, generally causes excitation and results in catabolism. This division is activated during periods of stress and exertion. The parasympathetic division is responsible for rest, digestion, and anabolism (i.e., building phase of metabolism). Walter Cannon originally suggested that these two divisions work in opposition and function at opposite times; in fact, they are generally both active and work in conjunction to control motor systems.
Included under the autonomic nervous system is the enteric nervous system, sometimes called the "little brain." Located within the walls of the digestive tract, this extensive network of neurons is a largely self-contained system consisting of the myenteric (or Auerbach's) plexus and the submucosal (or Meissner's) plexus that controls digestive functions. While under normal circumstances the other branches of the autonomic nervous system influence the enteric nervous system, if the digestive tract is deinnervated, such as occurs during severe spinal injuries, the enteric nervous system is able to maintain digestive functions. The enteric nervous system is discussed in more detail in Chapter 17.
Sympathetic division
The sympathetic division of the autonomic nervous system allows that body to respond to emergency situations resulting from sudden changes in the internal or external environment. It mediates an increase in alertness, heart rate, blood pressure, metabolism, respiration rate, sweating, piloerection, and mobilization of energy within the body. Simultaneously, it decreases activity of the digestive, urinary, and immune systems. It causes an increase in blood flow to the skeletal muscles while decreasing blood flow to the visceral organs.
In other words, the sympathetic nervous system activates those systems an animal needs in order to fight while inhibiting those systems not needed for fighting. If an animal is engaged in a fight, it needs increased blood flow to the skeletal muscles for increased muscular activity. Such blood flow comes at the expense of blood flow to the viscera. If an animal is shunting resources to fight, it does not need to be simultaneously digesting a meal; therefore, blood flow is shifted from the digestive tract to the skeletal muscle.
Parasympathetic division
The parasympathetic nervous system stimulates restful activities while inhibiting stress responses. Therefore, it is active during nonstressful conditions.
The parasympathetic nervous system promotes activities such as digestion while simultaneously conserving energy and decreasing blood pressure, heart rate, and respiration rate. Metabolic rate is decreased by the parasympathetic nervous system, and the pupils are constricted while the lens is allowed to become more convex in order to accommodate for close vision.Anatomy of the autonomic nervous system
Sympathetic division
The sympathetic division, also called the thoracolumbar division, exits the CNS from the thoracic and lumbar vertebrae. The preganglionic fibers have cell bodies in the Intermediolateral horn, sometimes called the lateral horns or visceral motor zones, of the spinal cord (Fig. 10.27). After leaving the spinal cord through the ventral root, the preganglionic sympathetic fibers enter the spinal nerve along with somatic motor fibers. Shortly thereafter, the sympathetic preganglionic fibers separate from the spinal nerve and pass through the white rami to enter the sympathetic chains (paravertebral chain) lying on either side of the spinal cord. Therefore, the preganglionic fibers are short, and the postganglionic fibers are long.
Upon entering the sympathetic chain, the myelinated preganglionic fibers can either synapse at that level, or they can travel rostrally or caudally before synapsing. The unmyelinated postganglionic fibers exit the sympathetic chain via the gray rami.
The sympathetic preganglionic fibers innervating the head exit the spinal cord from the first four tho-
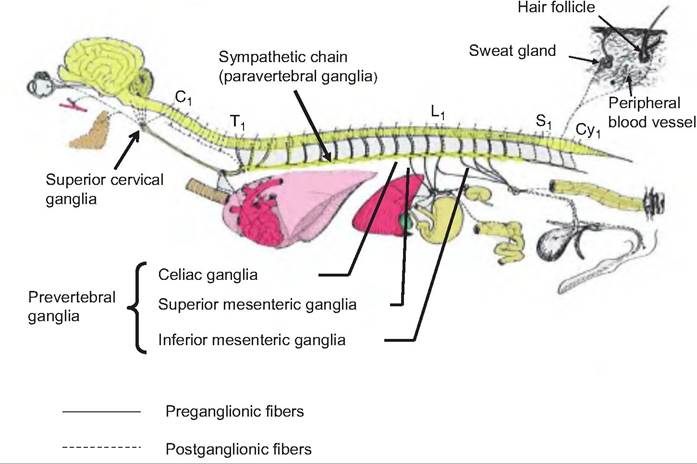
Fig. 10.28. Sympathetic (thoracolumbar) division of the autonomic nervous system. The preganglionic fibers of the sympathetic nervous system originate in the lateral horns of the thoracic and lumbar vertebrae. The fibers exit and travel as shown in Figure 10.2 to outlying ganglia. The postganglionic fibers originate in the outlying ganglia and travel to the target organs.
C1, T1, L1, S1, and Cy1 represent first cervical, thoracic, lumbar, sacral, and coccygeal vertebrae, respectively. (Figure modified from Getty, 1964.)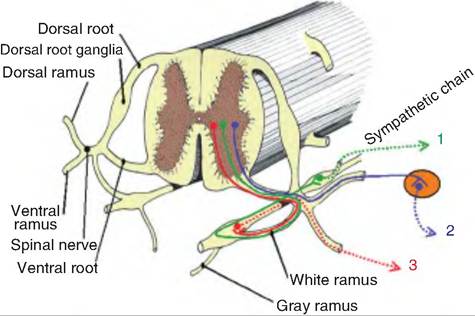
Fig. 10.27. Sympathetic chain and pathways. Preganglionic sympathetic fibers originate in the iIitermediolateral horn of the thoracic and lumbar vertebrae. These fibers can follow three paths: 1. As shown in green, the fiber can enter the sympathetic chain via the white ramus and continue to a different level within the chain where it then synapses on a postganglionic fiber that leaves the chain via the gray ramus. 2. As shown in blue, the preganglionic fiber can enter the spinal nerve and synapse on a postganglionic fiber in an outlying ganglion (prevertebral ganglion). 3. As shown in red, the preganglionic fiber can enter the sympathetic chain and synapse on a postganglionic fiber at the same level. (Figure modified from Getty, 1 964.)
racic segments (Tl-T4), enter the sympathetic chain, and ascend to the superior cervical ganglion, the most rostral extension of the sympathetic chain (Fig. 10.28). These fibers stimulate the dilator muscles for the irises of the eye, inhibit nasal and salivary glands, and innervate the muscle that lifts the eyelids. These responses are all typical of those observed during stress. Other fibers from the superior cervical ganglia innervate skin and blood vessels in the head, as well as sending branches to the heart.
Prevertebral ganglia
The preganglionic fibers from T5 caudally pass through the sympathetic chain before synapsing in preverte- bral ganglia. Prior to reaching the prevertebral ganglia, these fibers form nerves called splanchnic nerves (splanchnic = viscera) that include such nerves as the thoracic greater, lesser, and least splanchnic nerves; lumbar splanchnics; and sacral Splanchnics. The prevertebral ganglia include the celiac ganglion and the superior and inferior mesenteric ganglia. These ganglia differ from the paravertebral chain in that they lie anterior to the vertebral column, are unpaired, and occur only in the abdomen and pelvis. The postganglionic fibers from the prevertebral ganglia innervate the GI tract and its accessory organs, including the pancreas and liver. These fibers also innervate the kidneys, bladder, and genitalia.
Pathways to the adrenal medulla
Embryologically, the adrenal medulla and sympathetic ganglia arise from the same tissue. Therefore, the adrenal medulla is unique in that it consists of postganglionic cells of the sympathetic nervous system. The preganglionic fibers are found in the thoracic splanchnic nerves that pass through the celiac ganglion without synapsing before reaching the adrenal medulla. The postganglionic cells making up the adrenal medulla secrete norepinephrine and epinephrine when stimulated.
Parasympathetic division
The parasympathetic division, or craniosacral division, originates from brain stem nuclei and S2-S4 of the sacrum (Fig. 10.29). The preganglionic fibers synapse on postganglionic fibers either on or near the target organ (intramural ganglia). Therefore, the preganglionic fibers are long, and the postganglionic fibers are short.
The cranial outflow of the parasympathetic nervous system comes from four cranial nerves. These fibers originate in the Edinger-Westphal nucleus, the oculomotor nerve (III), the superior salivary nuclei associated with the facial nerve (VII), the inferior salivary nuclei associated with the glossopharyngeal nerve (IX), and the dorsal vagal nucleus and nucleus ambiguous, both associated with the vagus nerve (X). The oculomotor nerves are responsible for pupil constriction and bulging of the lens as occurs during accommodation when objects are moved closer in the field of vision. The facial nerves stimulate secretory activity from glands in the head such as the lacrimal and nasal glands and mandibular and sublingual salivary glands. The glossopharyngeal nerves activate the parotid salivary glands.
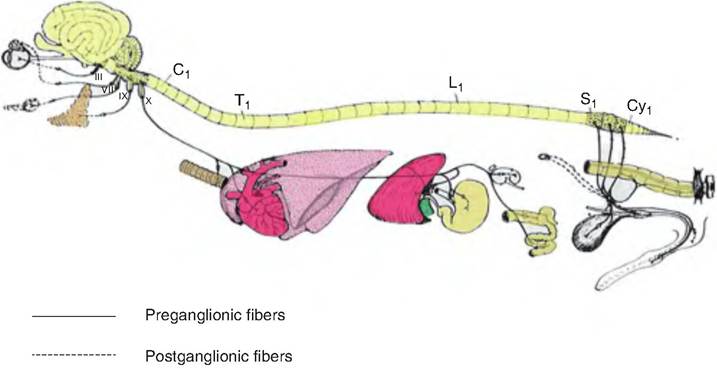
Fig. 10.29. Parasympathetic (craniosacral) division of the autonomic nervous system. The preganglionic fibers of the parasympathetic nervous system originate in the brain stem and sacral vertebrae. The fibers exit either as cranial nerves from the brain stem or spinal nerves from the sacrum and project to outlying ganglia located on or near the target organ. The postganglionic fibers originate in the outlying (intramural) ganglia and innervate the target organs. (Figure modified from Getty, 1964.)
The vagus nerves account for 90% of all preganglionic parasympathetic fibers. They provide input to the neck and all viscera in the thoracic and abdominal cavities. The vagus nerve mostly arises from the dorsal motor nuclei in the medulla, with its fibers synapsing on ganglia located on the target organ walls. These ganglia are not named, but instead collectively are called intramural ganglia, that is, "ganglia within the walls." After passing into the thorax, the vagus sends branches to various nerve plexuses, including the cardiac plexuses, the pulmonary plexuses, and the esophageal plexuses supplying the heart, lungs, and bronchi, and esophagus, respectively. Upon reaching the esophagus, fibers from the two vagus nerves intermingle and form the anterior and posterior vagal trunks.
The prevertebral fibers from the sacral spinal cord leave via the ventral roots and travel in the pelvic nerve to the pelvic ganglion plexus. These fibers innervate the descending colon, bladder, and external genitalia.
Physiology of the autonomic nervous system
Neurotransmitters and receptors
The major neurotransmitters in the autonomic nervous system are ACh and norepinephrine. Postganglionic neurons of the parasympathetic nervous system release ACh and are thus called cholinergic neurons; those of the sympathetic nervous system generally release norepinephrine and are called adrenergic fibers. The exception is sympathetic fibers innervating sweat glands, which release ACh.
Both sympathetic and parasympathetic preganglionic fibers generally release ACh that acts at nicotinic receptors to induce fast excitatory postsynaptic potentials in postsynaptic cells.
Cholinergic fibers
Acetylcholine can bind to two types of receptors called nicotinic and muscarinic. These receptors were named for pharmacological compounds that activate these receptors and are thus called agonists. Nicotine, the active ingredient in cigarettes, is the agonist for the nicotinic receptor; muscarine, which is found in poisonous mushrooms, is the agonist at muscarinic receptors.
Nicotinic receptors are found at skeletal muscle end plates, and in autonomic ganglionic neurons in both the parasympathetic and sympathetic nervous systems. Therefore, somatic neurons and all preganglionic neurons release ACh that acts at nicotinic receptors. The binding of ACh to nicotinic receptors always causes excitation of the effector cell. In autonomic ganglion, stimulation of the nicotinic receptors evokes a fast EPSP, usually triggering an action potential in the postsynaptic neuron.
All cholinergic postganglionic fibers act at muscarinic receptors. These include all parasympathetic fibers as well as those sympathetic fibers innervating sweat glands.
Adrenergic fibers
Postganglionic sympathetic fibers, except those innervating sweat glands, release norepinephrine. The sympathetic postganglionic fibers in the adrenal medulla release both norepinephrine and epinephrine. There are two major classes of adrenergic receptors: alpha (α) and beta (β). Norepinephrine and epinephrine act at both types of receptors. Target organs can have one or both types of receptors. While, in general, activating «-adrenergic receptors causes excitation and activating β-adrenergic receptors causes inhibition, there are notable exceptions to these rules because there are various subclasses of these classes of receptors. A summary of the actions of these receptors is shown in Table 10.4.
Note that norepinephrine can have opposite effects on vasomotor tone, depending on the dose of neurotransmitter (Fig. 10.30). When the dose of norepinephrine or epinephrine is relatively low, it causes vasodilation in skeletal muscle. Such a response is seen when an animal is alarmed or is experiencing a flight-or-fight response in which there needs to be increased blood flow to skeletal muscle to support its increased metabolic activity. This effect is mediated by the neurotransmitters acting at β2-adrenergic receptors. In contrast, the same neurotransmitters can cause vasoconstriction when these neurotransmitters are administered at high doses, and the effect is mediated by α i-adrenergic receptors. This latter response is observed only at doses of neurotransmitters above those present in vivo.
Other neurotransmitters
Acetylcholine can also evoke slow EPSPs and inhibitory postsynaptic potentials (IPSPs) in postganglionic neurons. These effects are mediated by muscarinic receptors. The slow EPSPs occur when ACh causes the opening of Na+ and Ca++ channels and the closure of M-type K+ channels. Since these K+ channels are normally open at resting membrane potentials, their closure results in depolarization.
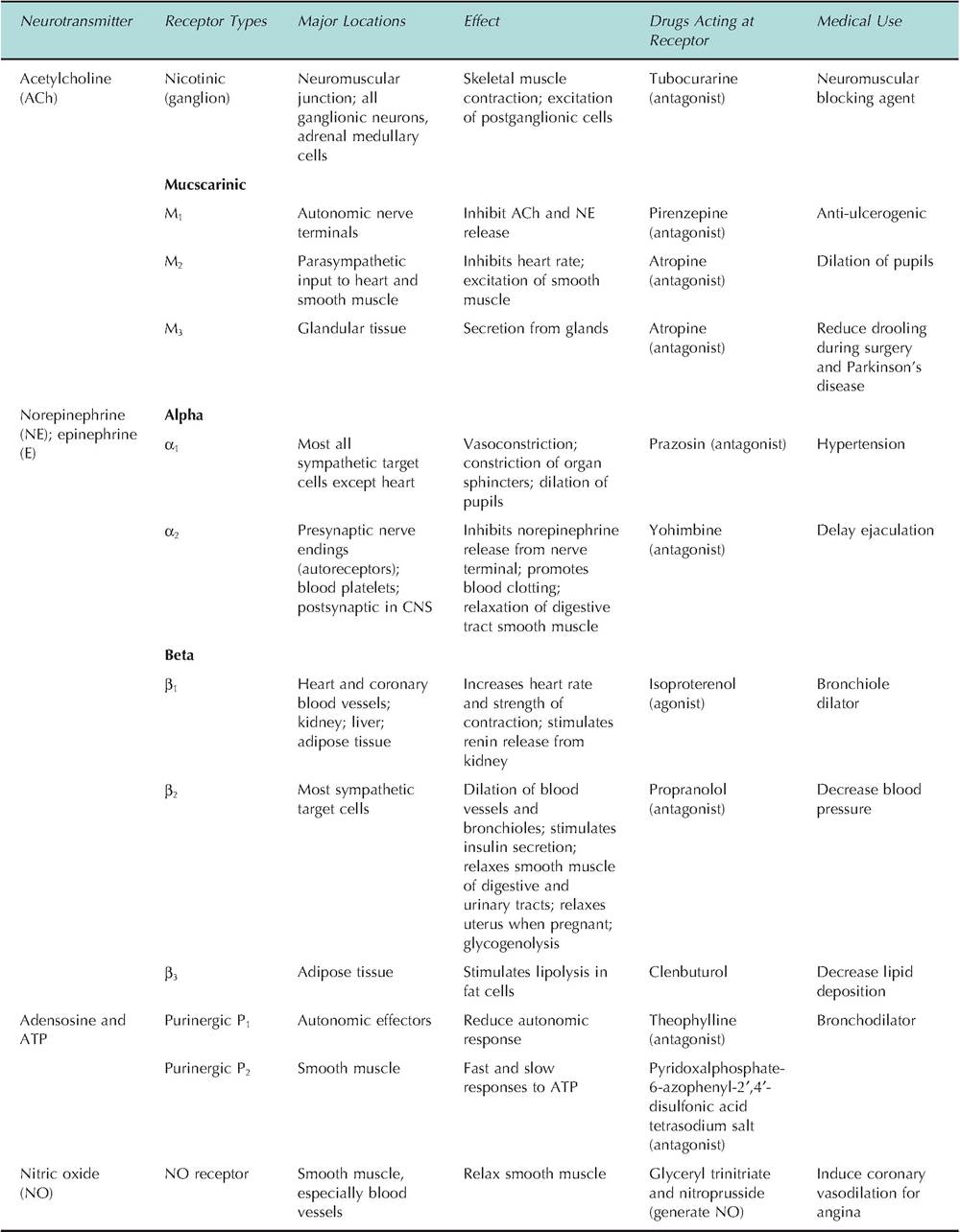
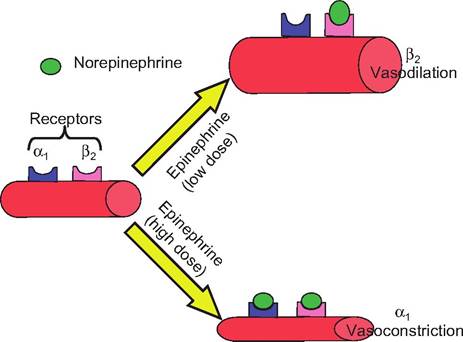
Fig. 10.30. Adrenergic effect on blood vessels. Blood vessels have both α1 and β2 receptors on their surface. Epinephrine and norepinephrine have differential effect on blood vessel tone depending on the concentration of the neurotransmitter. When the concentration of these neurotransmitters is low, they stimulate the β2-adrenergic receptors on the blood vessel walls, causing vasodilation. When the concentration is high, the primary effect is due to stimulation of the α1-adrenergic receptors, which causes vasoconstriction.
The slow IPSPs result from opening K+ channels resulting in hyperpolarization.
Adenosine triphosphate (ATP) is frequently coreleased with norepinephrine at many postganglionic sympathetic neurons. ATP can act at ATP-gated ion channels (P2 purinergic receptors) causing excitation. In addition, adenosine, which is produced by the hydrolysis of ATP, can act both pre- and postsynapti- cally at P2 receptors. Adenosine reduces the release of norepinephrine and ATP from nerve terminals, particularly after intense sympathetic activity.
Many neuropeptides are also coreleased with norepinephrine and ACh from autonomic neurons. Cholinergic preganglionic fibers may contain enkephalins, neurotensin, somatostatin, or substance P Cholinergic postganglionic fibers can also contain calcitonin gene-related peptide and vasoactive intestinal peptide (VIP). Corelease of VIP may enhance the effect of ACh since VIP causes vasodilation. For example, when ACh causes salivary gland secretion, VIP enhances blood flow to support the secretory response.
Noradrenergic postganglionic fibers may also release neuropeptide Y. At synapses where the nerve ending is more than 60 nm from the target tissue, neuropeptide Y enhances both the purinergic and adrenergic component of the tissue response. At synapses where the nerve ending is within 20 nm of the target, neuropeptide Y acts presynaptically to inhibit ATP and norepinephrine release. Sympathetic neurons can also contain galanin and dynorphin along with neuropeptide Y
Interactions of the sympathetic and parasympathetic divisions
Both divisions of the autonomic nervous system innervate most organs, with the two divisions having opposing effects. While both divisions are normally active, the effect on the organ is dependent on the relative activity of each division (Table 10.5).
Central nervous system control of the autonomic nervous system
The two divisions of the autonomic nervous system are highly coordinated at the level of the CNS. The nucleus of the solitary tract serves an important role in this regard. It receives visceral input from cranial nerves VII, IX, and X, and can then modulate autonomic functions in two ways. First, this nucleus projects to neurons in the brain stem and spinal cord, forming circuits to control autonomic function. For example, visceral sensory information transmitted through this nucleus is used to regulate vagal motor control to the stomach and heart. Second, this nucleus integrates autonomic functions with endocrine and behavioral responses largely through its interaction with the hypothalamus.
The hypothalamus was once termed the "head ganglion" of the autonomic nervous system. Early studies on brain function found that stimulating or Iesion- ing the hypothalamus altered hypothalamic function. Recent studies have revealed that these effects were due to interruption of ascending and descending pathways that course through the hypothalamus traveling between the cerebral cortex and basal forebrain.
The hypothalamus serves a role in regulating five autonomic functions:
1. It regulates blood pressure and electrolyte composition by controlling fluid and salt intake, thus maintaining blood osmolality and volume.
2. It regulates body temperature by controlling the set point for body temperature and activating either heat loss or heat production pathways.
3. It regulates energy metabolism by controlling food intake, digestion, and metabolic rate.
| Table 10.5. Parasympathetic and sympathetic effects. | ||
| Target Organ | Parasympathetic Effect | Sympathetic Effects |
| Eye (Iris) | Constricts eye pupils | Dilates eye pupils (α1) |
| Eye (ciliary muscle) | Makes lens more convex for accommodation | No effect |
| Glands (nasal, lacrimal, salivary, gastric, pancreas) | Stimulates secretion | Inhibits secretion; causes vasoconstriction of blood vessels to glands |
| Sweat glands | No effect | Stimulates secretion via cholinergic fibers |
| Adrenal medulla | No effect | Stimulates NE and E secretion |
| Arrector pili muscles | No effect | Contraction causing erection of hairs and goosebumps |
| Heart muscle | Decreases heat rate and strength of contraction | Increases heart rate and strength of contractions |
| Coronary blood vessel | Vasoconstriction | Vasodilation |
| BIadderAirethra | Contraction of bladder wall; relaxation of urethra sphincter; void urine | Relaxation of bladder wall; constriction of urethra sphincter; inhibits urine release |
| Lungs | Constricts bronchioles | Dilates bronchioles |
| Digestive tract | Increases peristalsis and secretion of digestive juices; relaxes sphincters | Decreases digestive motility and secretion |
| Liver | No effect | Epinephrine stimulates glycogenolysis |
| Gallbladder | Stimulates contraction | Inhibits contraction |
| Kidney | No effect | Vasoconstriction; decreased urine output; increases renin formation |
| Penis | Erection via vasodilation | Ejaculation |
| VaginaArIitoris | Erection via vasodilation of clitoris | Reverse peristalsis or contraction of vagina |
| Blood vessels | Vasodilation via production of nitric oxide | Constricts most vessels increasing blood pressure; dilates vessels in skeletal muscle, heart, and bronchioles |
| Blood coagulation | No effect | Increases coagulation |
| Cellular metabolism | No effect | Increases metabolic rate |
| Mental activity | No effect | Increases alertness |
| Adipose tissue | No effect | Stimulates Iypolysis |
4. It controls an animal's response to stress by regulating adrenal function, blood flow to muscles, and immunological responses.
5. It regulates reproductive functions, including mating, pregnancy, and lactation.
The hypothalamus is well positioned to coordinate these functions since it receives sensory information from most of the body, it is able to compare that information with endogenous set points, and it can then make appropriate adjustments via negative feedback loops to maintain homeostasis. Regarding sensory information, the hypothalamus receives direct input from the visceral sensory system, the olfactory system, and the retina. Information from the retina is used by the Suprachiasmatic nucleus to synchronize the internal clock and control circadian rhythms. Pain information is carried to the hypothalamus from the spinal and trigeminal dorsal horns. In addition, the hypothalamus contains sensory neurons that monitor temperature, osmolality, glucose, and sodium levels. Signals from the periphery, such as blood leptin or angiotensin II levels, are also able to reach the hypothalamus because of specialized structures called cir- Cumventricular organs, which allow peptides to cross the blood-brain barrier at these sites.
Visceral reflexes
Although the definition of visceral reflexes was discussed earlier, here we describe specific visceral reflexes. Visceral reflexes control an array of autonomic responses. Some responses are very rapid, such as change in pupil size; others are relatively slow, such as gastric secretions.
Ocular reflexes
The diameter of the pupil and the shape of the lens are controlled by the autonomic nervous system. Sympathetic fibers originating from the superior cervical ganglia innervate the muscles controlling dilation of the pupil; parasympathetic fibers innervate circular muscles constricting the pupil. When excited, the autonomic nervous system inhibits pupillary constriction while it stimulates pupillodilator muscles.
Cardiovascular reflexes
Arterial blood pressure is controlled by both cardiac output and resistance to blood flow. The sympathetic nervous system can increase heart rate, strength of cardiac contraction, and peripheral resistance while the parasympathetic nervous system can decrease heart rate and peripheral resistance, although its effect on peripheral resistance is less than that of the sympathetic nervous system.
Sympathetic vasoconstrictor tone is controlled by adrenergic neurons originating in the ventrolateral medulla that innervate sympathetic vasoconstrictor preganglionic neurons. Stimulation of baroreceptors sends signals to the nucleus of the solitary tract, which excites interneurons in the caudal ventrolateral medulla leading to inhibition of vasomotor neurons and excitation of vagal cardiomotor neurons. This results in a decrease in blood pressure and heart rate.
Norepinephrine is released from sympathetic fibers innervating the heart and acts on the β-adrenergic receptors in the heart to increase heart rate and force of contraction. It acts by increasing the production of cyclic adenosine monophosphate (cAMP) that in turn opens long-lasting (L-type) Ca++ channels. Norepinephrine also decreases the threshold for firing of cardiac pacemaker cells (located in the sinoatrial [SA] node), leading to increased heart rate.
Acetylcholine is released from parasympathetic fibers innervating the heart and acts on muscarinic receptors in the SA and atrioventricular (AV) nodes of cardiac muscle. This causes an increase in K+ conductance in these cells, which causes hyperpolarization of the SA and AV nodes, thus slowing transmission through these sites. ACh also increases the threshold for firing the pacemaker cells in the SA node. The strength of contraction of cardiac muscle is also decreased by ACh by decreasing the intracellular cAMP, thus reducing the L-type Ca++ current.
Glandular reflexes
The parasympathetic nervous system stimulates gastrointestinal glands such as the nasal, lacrimal, and gastric glands. Glands located in the upper GI tract, particularly in the mouth and stomach, are more strongly stimulated than those in the lower GI tract. Glands in the lower part of the GI tract are mostly under the control of the enteric nervous system. Salivary glands are under both sympathetic and parasympathetic control. Sympathetic input causes a viscous secretion high in amylase; parasympathetic input causes a watery secretion of higher volume.
In general, the sympathetic nervous system decreases glandular secretions, while the parasympathetic nervous system increases glandular secretions. This is because the sympathetic nervous system generally decreases blood flow to glands. The exception is sweat glands, in which the sympathetic nervous system increases sweating. Sympathetic fibers to the sweat glands are cholinergic rather than adrenergic.
Gastrointestinal reflexes
The parasympathetic nervous system stimulates gastric acid secretion, whereas the sympathetic nervous system inhibits such function. The enteric nervous system controls peristalsis. The presence of a bolus of food in the GI tract causes stretch in the gut walls, generating sensory signals that synapse on interneurons within the gut wall. These interneurons initiate a stimulatory motor signal to the circular muscle layer rostral of the bolus while simultaneously causing inhibition of motor neurons of the circular muscle layer caudad of the bolus. In addition, the interneurons send signals resulting in stimulation of the longitudinal muscle layer caudad of the bolus while inhibiting the longitudinal muscle layer rostral of the bolus.
Urogenital reflexes
Bladder emptying is generally under autonomic control, although there can be some voluntary control. When the bladder is extended, there is a visceral sensory reflex in which parasympathetic postganglionic neurons in the pelvic ganglion plexus promote contraction of the bladder. The sympathetic nervous system promotes the bladder's smooth muscle relaxation.