AVIAN TUBERCULOSIS
RUTH CROMIE
Wildfowl & Wetlands Trust (WWT), Slimbridge, Gloucester, UK
Avian tuberculosis (synonyms: avian mycobacteriosis, M. avium intracellulare disease, M. avium intracellulare complex disease or M.
avium complex disease) is an insidious, chronic, bacterial disease of birds, most commonly reported in waterbirds, gregarious birds, raptors or those associated with agricultural premises. Mycobacterium avium is the principle cause of the disease in birds.AETIOLOGY
As both saprophyte and opportunist facultative pathogen, M. avium is a ubiquitous, slow-growing, environmental bacterium favouring acidic conditions rich in organic matter. Much of the organism’s ecological success, both in host and environment, is due to the lipid-rich, thick, waxy cell wall, which helps to protect it from desiccation, freezing and ultraviolet radiation, and accounts for its resistance to both antibiotics and disinfectants. In the environment, the hydrophobic cell wall is responsible for the organisms being found at the air-water interface, leading to the reference to the mycobacteria as ‘the ducks of the microbial world’(18). Mycobacterium avium’s ability to grow at temperatures between 20 and 45°C allows it to exploit birds as a host.
The classification of M. avium and the means by which this can be done are as complex as the organism itself. It has a large number of biochemically identical stable serotypes distinguished by the numbers of polar glycopepti- dolipids present in the cell wall. Using classical agglutination serotyping, M. avium consists of serotypes 1-3 (the most common avian pathogens), with M. intra- cellulare including serotypes 4-24. To this end M. avium is sometimes known as the M. avium complex (MAC), M. avium intracellulare (MAI) or the M. avium intracellulare complex (MAIC). Within this complex lies the unusual mycobactin-dependent strain responsible for the disease in wood pigeons (Columba palumbus), often referred to as the wood pigeon bacillus(19).
This mycobactin dependence is shared by the closely related M. avium subsp. paratuberculosis responsible for Johne’s disease. Avian disease caused by these different serotypes within the MAC are indistinguishable. Molecular techniques have been highly valuable for diagnosis and epidemiological investigations and have overturned many epidemiological assumptions. They have also revealed insertion sequences linked to avian virulence: IS1245 and the closely related IS901(20).EPIDEMIOLOGY
GEOGRAPHICAL DISTRIBUTION AND HOSTS
The disease has a global distribution, with the majority of wild cases being reported in the northern temperate zone. Cases have been reported across Europe and the absence of disease in a region probably reflects lack of reporting. Europe sits within a number of major migratory bird flyways, and for such a chronic slowly progressing disease, cases in Africa and Asia and, to a lesser extent north America, may still have some importance in the epidemiology of the disease in Europe and vice versa.
Mycobacterium avium is the principle cause of avian tuberculosis in wild, domestic and captive birds and also causes infections in an extremely wide vertebrate host range, including free-living European mammals such as: mustelids, e.g. Eurasian otters (Lutra lutra)(21); rodents, e.g. European beavers (Castor fiber)(22) and yellow-necked mice (Apodemus flavicollis)2i; cervids, e.g. red deer (Cervus elaphus)(24); suids, e.g. wild boar (Sus scrofa)14i; and members of the Insectivora, e.g. European hedgehogs (Eri- naceuseuropaeus)124’ and common shrews (Sorexaraneus)(23). In captive situations the host range is even broader, including marsupials, mustelids, primates, a wide range of ungulates and even fish and reptiles.
The disease has been described in a wide range of bird species. This is probably in part because of differences in genetic/inherent susceptibility.
This has been suggested from captive studies in which, even when exposed to infection, the majority of birds did not succumb to disease. The genetic basis of susceptibility to mycobacterial infections has been largely determined for laboratory mice and is likely to be similar for birds. This is important for some highly susceptible threatened species in captivity that suffer from founder effects and inbreeding. A high prevalence of avian tuberculosis in free-living threatened whooping cranes (Grus americana) in the USA (39% for one population) has negatively affected this conservation pro- gramme(27), and genetic factors may be at play.Of equally great importance in susceptibility and risk of infection are life history, ecology and behaviour. In the wild, the disease is most commonly reported in:
• waterbirds, with a ‘usual’ prevalence of 1—6% of mortalities
• birds associating with agricultural premises, where prevalence in e.g. starlings (Sturnus vulgaris) and sparrows (Passer domesticus) has been reported between 5 and 40%(28); for this group there are two main risk factors, namely contact with potentially infected poultry and high population densities due to abundance of food supply and nest sites; however, it should be recognized that they are also the subject of greater investigation and thus reporting than other bird groups
• gregarious birds: this group incorporates many of the first two categories, but additionally a gregarious lifestyle puts birds at risk due to the density dependence of the disease; as an example, the disease has been diagnosed in domestic and free-living feral pigeons (Columba livia domestica) with a prevalence of 1%(29), and in 3% of crows(30)
• raptors and scavengers: for this taxon, population density is less of an issue but cases reflect consumed infected prey or secondary infections of wounds; reports in raptors may reflect the higher rate of discovery of carcasses or sick/injured birds and greater likelihood of investigation given the greater interest frequently shown for this family of birds; European reports estimate between 1 and 4% prevalence of avian tuberculosis1-31’32’.
In captivity, avian tuberculosis is typically a disease associated with older birds, and given the slow progress of the disease this is likely to be the case for wild birds. The relatively low age structure of many wild populations may assist in keeping prevalence low.
There is no apparent sex predilection for disease susceptibility. The role of stress should not be underestimated, and periods of nutritional or other stress are certain to have an adverse impact on latent infection, leading to conversion to clinical infection or reducing the host’s protective responses during the prolonged immunological battle with the host.
ENVIRONMENTAL FACTORS
Appreciating the ecology of M. avium allows an understanding of why birds in damp or wet environments are at particular risk. High densities of hosts have the impact of both increasing possible stresses but also increasing the rate of encountering infectious individuals. Contact with potentially infected domestic or captive birds is an obvious and important risk factor. Access to infected prey or food materials is a clear risk factor that is reflected in the numerous reports in predatory and scavenging birds.
The role of exposure to environmental mycobacteria in sensitization of the immune system and subsequent disease outcome is well recognized in humans(18) and thought to occur in captive birds. It is probably of critical importance to the disease outcome in wild birds but too complex to evaluate or test.
EPIDEMIOLOGICAL ROLE OF WILD ANIMALS
Outcomes of infection are difficult to determine in wild birds. It is assumed that birds either: overcome initial infection (the degree to which this protects from further infection is not known); develop latent infection, which may reactivate e.g. during a later period of stress; or develop progressive disease, during which time they may be infectious and shed bacilli.
Infected and infectious birds play two roles in the epidemiology of the disease, firstly as direct shedders into the environment, and secondly through being eaten and infecting predators and scavengers directly.
The debilitating nature of the disease would probably allow disproportionate numbers of tuberculous prey species to be caught by predatory species.Migratory birds, particularly those with periods of their life cycle in which population densities are high, e.g. wintering or staging waterbirds, or communally winterroosting species, have the greatest potential for becoming infected and spreading infection across great distances. Raptors, even if migratory, are less likely to be involved in significant onward spread of infection.
TRANSMISSION
Mycobacteria are shed in faeces from lesions in the intestinal tract, and thus the principle source of pathogenic avian tubercle bacilli is a faecal-contaminated environment. Oral ingestion accounts for the majority of cases, and how a bird interacts with its environment during feeding may be key in determining ingestion rates — for example, dabbling waterfowl sifting food from muddy areas at air-water interfaces are likely to ingest more bacilli than selectively grazing geese. Inhalation of aerosols, either from a contaminated environment or directly from infected birds with lesions in the respiratory tract have been suggested as the cause of primary pulmonary infections in domestic or captive birds at least(33).
The role of the egg in transmission is intriguing. It has long been assumed that vertical transmission does not occur, as infected poultry give rise to apparently uninfected young. However, there are a small number of reports of M. avium-infected eggs(34). It is unknown whether or not these would have gone on to develop into healthy embryos, and it is speculated that they may have accounted ultimately for embryo death, but the role of the egg can be viewed as relatively unimportant in the epidemiology of this disease.
Contamination of wounds has been recorded in raptors, and other possible, if less likely, sources or modes of transmission have been suggested, including ticks, rodents, beetle and fly larvae, and coitus.
PATHOGENESIS, PATHOLOGY AND IMMUNITY
Most often M. avium enters the body through ingestion of bacteria while feeding. What constitutes an infectious dose has not been defined for wild birds, but it is speculated that infection is probably acquired over time following repeated ingestion of pathogenic strains of M. avium from a contaminated environment.
Following either ingestion or inhalation, the lipid-rich cell walls help protect against initial physical and chemical innate defence processes within the host. It is then assumed that bacteria bind to enterocytes in the intestinal mucosa, with eventual haematogenous spread to the most frequently involved organs: the liver and spleen.
Although the intestinal tract, liver and spleen are commonly affected organs, the disease can be generalized, with involvement of practically any organ. Infections in bones are not uncommon. Disease spread within the body is likely to take place over several months.
Host responses ensure that during late stages of disease coalescing granulomatous masses may replace extensive amounts of functional tissue, and death may be as a result of this loss of organ function or due to rupture of an infected organ.
Alternatively this chronic debilitating disease may render the bird more likely to predation or other infectious processes, so avian tuberculosis may not be the proximate cause of death. As an example, the mass mortality of lesser flamingos (Phoenicopterus minor) in Kenya reported in 1999(35) was due to other factors, but high levels of background avian tuberculosis were felt to have weakened the birds.
A tuberculous bird is usually in poor condition, with muscle wasting and loss of body fat. Disease may be disseminated or restricted to one tissue. Lesions range in
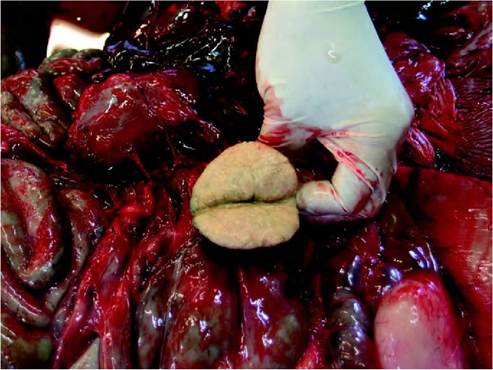
FIGURE 20.4 Multiple tuberculous lesions in the liver of a pinkfooted goose (Anser brachyrhynchus') shot and opened by hunters in Iceland. Photo: Ruth Cromie.
colour from white through pale grey, cream or yellow to tan and in size from diffuse military foci to coalescing masses, several centimetres in diameter)36). Although lesions are most typically found in the intestines, liver and spleen, either on serosal surfaces or within parenchyma, any tissue can be affected. Lesions may be discrete and encapsulated or diffuse into the surrounding tissue. Granulomas on the intestinal wall and mucosa may be ulcerated, allowing shedding of bacilli into the lumen. Lung and bone involvement are not uncommon. Infected joints are usually swollen and frequently contain caseous material. Granulomas may contain caseous material or be tumour-like in cross-section. Figure 20.4 shows extensive lesions in the liver of a pink-footed goose )Anser brachy- rhynchus) shot by hunters in Iceland.
Affected organs may be enlarged and/or uniformly pale in colour. Such findings may also be found in the nontuberculoid form of the disease, which is typified by histiocytic inflammation, or histiocytosis with diffuse, concurrent amyloidosis(37). This histiocytic form may be characteristic of early stages of the disease)38).
Microscopically, in early stages of infection there may be scattered foci of epithelioid macrophages and lymphocytes. Macrophages may be seen containing large numbers of bacilli with associated vacuolated/foamy cytoplasm. As the disease progresses, better-defined caseous granulomas may be seen. The most typical form of granuloma involves a central area of necrosis with surrounding epithelioid macrophages and lymphoctyes. Multinucleated giant cells may or may not be present and a capsule of connective tissue may or may not surround the granuloma. Alternatively, lesions may be made up of aggregations of histiocytes. Acid-fast bacteria can usually be demonstrated, either within macrophages or within the central necrotic debris.
In the non-tuberculoid form of the disease the organ enlargement is caused by diffuse infiltration of large foamy histiocytes)37).
Avian tuberculosis is typified by a protracted immunological battle with the mycobacterium’s waxy cell wall, responsible for much of the pathogenesis. Mycobacterial immunity is classically dependent on cell-mediated responses with humoral immunity playing a less important role.
Phagocytosis and killing by macrophages is key to host immunity to M. avium. Although the precise mechanisms at play within the avian macrophage are not fully understood it is assumed that the mycobacteria can survive and multiply within these cells by inhibiting phagosome— lysosome fusion, thus avoiding proteolytic enzymes and also disrupting cytokine production)37) and inactivating superoxide radicals produced by the host cell)39). Glycosides from the bacterial cell wall promote the host’s granulomatous response and enhance intracellular survival. Appropriate T-cell responses can induce killing of contained mycobacteria or the host response attempts to ensure bacteria can be ‘held at bay’ within activated macrophages within these granulomas. Host cells expressing mycobacterial antigens may be killed, resulting in tissue necrosis. Loss of protective cell-mediated responses (owing to nutritional stress, for example) can lead to escape of multiplying bacteria from granulomas and spread to other parts of the body.
CLINICAL SIGNS AND TREATMENT
In general there are few specific signs for avian tuberculosis in the field. Clinical presentation is most typically chronic wasting, with birds becoming emaciated and often exhibiting a prominent keel, or breast bone. Birds are usually weak and lethargic, often with poor or ruffled plumage. In late stages of the disease, abdominal distension as a result of liver enlargement and a build-up of ascitic fluid can give an emaciated bird an unusual ‘bottom-heavy’ appearance. Lameness is relatively common if there is bone
TABLE 20.3 Illustrative levels of Ieucocytosis for various avian species with avian tuberculosis.
| Species | Situation | Normal values | Tuberculous values | Reference | |
| Range ? 109∕l | Range ? 109∕l | Mean ± sd (if available) ? 109∕l | |||
| Quail | Experimentally | 4.75 | 8.6-26.25 | 21.75 | 42 |
| infected | n = 32 (mean range) | ||||
| Domestic fowl | Naturally | 3.7-11.9 | 10.3-230.0 | 52.5 ± 48.6 | 43 |
| infected | n = 30 | n = 26 | |||
| Raptors | Free-living | 11-20 | 5-850 | 170 ± 136 | 45 |
| n = 6 | n = 6 | ||||
| Various zoo birds | Captive zoo | Not given | Not given n = 17 | 62.475 | 42 |
| Cranes | Captive zoo | 2.1-20.1 | 6.2-214.4 | 47.9 ± 45.3 | 43 |
| n = 57 | n = 17 | ||||
| Various species | Captive zoo | 3.5-13.4 | 18.8-61 | 32.7 ± 18.4 | Cromie (unpublised |
| of duck | n = 40 | n = 6 | observations) | ||
involvement. Chronic or intermittent diarrhoea is common.
Ceres and other areas of exposed skin may become progressively paler as the disease progresses. There may be cutaneous or subcutaneous lesions, particularly in raptors)32), and even ocular lesions)40). Respiratory involvement is relatively unusual but this may result in dyspnoea. Involvement of the vertebral or central nervous system may account for neurological signs. Alternatively birds may just be found dead or succumb to another cause of death before these clinical signs are apparent.
Treatment of captive birds with anti-tuberculous drugs is complex and rarely recommended for large-scale use. Immunotherapy has been tried on a small scale but requires further research for treatment of disease in threatened species(41).
DIAGNOSIS
Diagnosis in wild birds, particularly ante mortem, is both complex and far from satisfactory and relies on a combination of tests before a positive diagnosis can be made.
HAEMATOLOGY
Studies with both captive and feral birds have found haematological screening to be relatively useful for ante mortem diagnosis)42-14). Avian tuberculosis often results in marked leucocytosis due to monocytosis, and/or heterophilia. There may also be a lymphocytosis, and mild to moderate anaemia. Table 20.3 provides some values of leucocytosis that have been reported as being indicative of avian tuberculosis in various avian taxa.
Haematological changes may reflect the stage of infection, with clinical cases generally exhibiting the greatest haematological abnormalities. However, during late stages of the disease birds can decline into a state of anergy, which may confound this situation.
Although haematological abnormalities such as leuco- cytosis are not specific for avian tuberculosis, in combination with other data on body condition or other epidemiological factors, they can be useful for suggestive, or preliminary, diagnosis.
Serum Biochemistry
There are no reports of reliable and consistent diagnostic serum biochemical changes during avian tuberculosis. Some authors report elevations in serum aspartate aminotransferase or globulin. Serum biochemistry values are often unremarkable during this disease, at least for pet birds)46).
Haemagglutination
The haemagglutination test is a simple, rapid test that relies on agglutinating antibodies in a drop of serum or whole blood when mixed with a drop of prepared antigen on a white tile. Results are highly variable, with false positives a particular problem, making it of limited practical use.
Complement Fixation Tests
Poor correlation has been found between clinical status and presence of complement-fixing antibodies — hence this is of limited value for diagnosis.
ELISA
Measurement of humoral responses for diagnosis of active avian tuberculosis is fraught with interpretation problems, mainly because of cross-reactivity of antibodies to environmental mycobacteria. However, some ELISA have achieved both high levels of sensitivity and specificity for some avian taxa(44). One of these assays had some capability of predicting tuberculous death within a year of testing1-41).
Ultrasonography
This can be used to identify changes in organ size and possibly extremely large tuberculous masses but is of limited value as a stand-alone diagnostic test.
Laparoscopy
For lesions on serosal surfaces of organs such as the liver, spleen, intestines, lungs or air sacs, laparoscopy offers a means of both visualization and potential biopsy for confirmative diagnosis, but lesions within the parenchyma or less accessible organs or body parts will go undisclosed. Additionally the need for anaesthesia makes this of limited value for working with wild birds.
Tuberculin Test
In poultry this intradermal test involved injection of avian tuberculin into a wattle or comb of a chicken, with resultant thickening at the site of injection within 48—72 hours. It was a powerful tool in managing the disease in these domestic birds for decades.
Areas of accessible suitable skin, let alone a handy dangling wattle, are absent from most wild birds. The necessity to hold a bird for 48—72 hours coupled with the highly variable results from exotic birds make this test of negligible value for the wildlife diagnostician.
Microscopy
The ZN stain has long been used to identify mycobacteria within lesions, faeces or aspirates. Other staining techniques involve use of fluorescent dyes (such as auramine or acridine orange). Microscopy alone cannot diagnose M. avium, merely the presence of acid-fast organisms. Further tests (e.g. molecular techniques or culture) are required for positive identification.
Cytological examination for M. avium-like bacilli is a relatively low technology test, but numbers of bacteria within lesions (from which smears may be made) can sometimes be low and therefore easily missed on examination. Faecal samples can be tested, but negative results may indicate a bird not currently shedding. Alternatively ‘positive’ samples may indicate the presence of other non- pathogenic environmental mycobacteria merely passing through the intestinal tract.
Culture
Culture of the aetiologic agent remains the ‘gold standard’ for diagnosis. However, in general, culture of mycobacteria requires specialist laboratories, as the bacteria have very specific growth requirements e.g. Lowenstein-Jensen or Middlebrook 7H10 culture medium with particular temperature and oxygenation settings. The BACTEC liquid medium system with radio-labelled reagents has reduced a usual 6-week culture time to approximately 2 weeks. Once cultured, identification can be made on colony characteristics and biochemical testing. Alternatively DNA probes can be used or techniques such as high-performance liquid chromatography(47).
Molecular Probes
Molecular techniques for detection of M. avium antigens or gene sequences have greatly improved diagnosis and aided epidemiological investigations. Most involve liberating mycobacterial DNA, DNA amplification by PCR and use of DNA probes to identify sequences. These techniques can be used on tissues (even formalin fixed), biopsies and faeces although the amount of organic material present in the latter can be a problem due to inhibition of PCR.
MANAGEMENT, CONTROL AND REGULATIONS
Avian tuberculosis is difficult to diagnose and treat; it has an insidious nature, with extended shedding periods, a broad host susceptibility and long environmental persistence. Therefore it is important to prevent its establishment in the wild by reducing risk factors, such as contact with infected domestic or captive birds.
The most effective disease control options for domestic and captive animal situations include routine surveillance with resultant culling of infected flocks, cleansing and disinfection (C&D) as happens in poultry farms. Zoo responses often involve euthanasia or quarantine of birds and complete exhibit renovation with C&D. Host immunity can be boosted by reducing stress, better nutrition and even immunoprophylaxis, although the latter is not known to have been tested in wild settings. Risk of infection can be reduced by ensuring good standards of hygiene, aided by use of certain disinfectants[18] [19] [20] [21] [22]-37). Although M. avium is more resistant in the environment than many other bacteria, ultraviolet radiation is a useful sterilizing agent, and exposing substrate (with turning as appropriate) to bright sunlight reduces mycobacterial load (during this time vertebrates should be excluded). Raising pH by the addition of lime is also valuable. Reducing population density (which can also serve to reduce stress) is also recommended. Reedbed treatment systems have been found to remove pathogenic M. avium from watercourses and should be considered where appropriate1-48). In wildlife situations the following management options are available: 6. stresses to wild birds such as anthropogenic disturbances should be minimized, particularly during unavoidable stressful times, e.g. cold weather 7. healthy habitat management should aid reduction of nutritional stresses; paradoxically, provision of supplemental food may reduce nutritional stress and reduce feeding from a potentially contaminated environment yet can increase disease risk through increased population density and subsequent environmental contamination; where supplemental food is provided to raptors, this should be checked to ensure it is free of infection (avoiding avian meat is strongly advised) 8. within wetland systems, constructed treatment wetlands can be created to reduce contamination by M. avium and other pathogens 9. if there is a serious outbreak at a site, extreme measures to decontaminate and exclude wild birds can be taken, e.g. draining, turning soil/vegetation and substrate to expose to ultraviolet radiation while scaring birds away from the area(49) 10. reared game birds must be subjected to rigorous health screening before release 11. for conservation reintroduction programmes, raising birds in biosecure environments from eggs with thorough health monitoring is advised. Although not a notifiable disease, avian tuberculosis in wildlife is classified as an OIE reportable disease. Among other reasons, difficulties in diagnosis prevent rigorous specific regulations with respect to trade or movements of domestic or captive birds; however, health certification measures should ensure that those birds showing signs of disease are excluded. PUBLIC HEALTH CONCERN The uncommon occurrence of avian tubercle bacilli infection in people even where the disease is common in poultry indicates that, in general, humans are extremely resistant to the pathogen. Given the relatively low prevalence of infection in wild populations of birds and this resistance, the disease in wild birds poses relatively little risk to public health. People most at risk are those with pre-existing underlying chronic pulmonary problems, neoplastic conditions or immunodeficiencies. The advent of HIV/AIDS has increased the incidence of human cases of MAC infections significantly. Although the developing world is burdened with co-infections of M. tuberculosis and HIV, in the developed world MAC is a relatively common co-infection for persons with AIDS. However, serotyping and molecular epidemiological investigations indicate that these MAC infections are generally associated with other environmental MAC strains rather than those of an animal origin, e.g. MAC from domestic supplies of water and other sources within human habitation. It has been stated that ‘for practical purposes all MAI infections in humans are of environmental origin’(19). Evidence of direct transmission from wild birds to humans is lacking, but high-risk persons, or those handling infected carcasses or working in avian tuberculosis control situations, should take appropriate and prudent health and safety precautions. SIGNIFICANCE AND IMPLICATIONS FOR ANIMAL HEALTH Poultry are very susceptible to avian tuberculosis, which can cause both reduced egg production and high mortality with the additional economic impacts of control programmes. Changes in intensive poultry production (such as improved hygiene and, importantly, culling at an early age) have reduced significantly the disease from most European domestic stock. The disease still occurs sporadically in smaller poultry flocks. Free-ranging birds, including farmed ratites, are at greatest risk of infection from wild birds. In general, disseminated MAC infection in domestic mammals is rare and less progressive than the more frequently seen mammalian mycobacterial infections. The infection is more typically diagnosed at slaughter as localized lymph node lesions in e.g. cattle and pigs. Pigs are readily infected with M. avium, with numerous reports of lymphadenitis. It has long been assumed that wild birds have provided the source of infection for pigs; however, like immunocompromised humans, they may just be susceptible to environmental strains(20). Cattle are relatively resistant to MAC and generalized infections are extremely rare. The greatest significance of MAC to cattle is in its non-specific sensitization to bovine mycobacterial antigens, hence reducing the diagnostic value of the intradermal bovine tuberculin test. This antigenic cross-reactivity has resulted in the widespread use of avian and bovine tuberculins being used as comparative tests in many countries. There have been numerous reports of M. avium infection in farmed deer, and these animals are considered reasonably susceptible. Cats and dogs are considered to be highly resistant, and reports of MAC infections in horses, sheep and goats are rare. Cases of avian tuberculosis in wild birds have been recorded throughout the last century. Whether prevalence in wildlife has changed over this time cannot be determined, owing to paucity of standardized long-term surveillance data across broad geographical scales. The disease rarely causes significant outbreaks, and as such is likely to remain, largely, a persistent but relatively unseen cause of morbidity and mortality in wildlife. The reduction of avian tuberculosis in poultry farms is likely to have reduced point sources of infection for wild birds, although the disease within captive collections of birds remains a problem. Rearing, release and intensive management of game birds poses a risk, although close disease surveillance within the rearing flock should help to minimize risks, as does the young age of the birds at time of release. Recovery programmes for threatened bird species frequently use captive environments for the birds, which is associated with risk of infection. Translocations themselves have potential risks for introduction of disease that need to be carefully managed. Threatened and insular species in the wild may be particularly vulnerable to the disease owing to loss of heterozygosity, and additionally the death or morbidity of a few key individuals can have a profound effect on a population. PARATUBERCULOSIS OR JOHNE'S DISEASE AETIOLOGY Mycobacterium avium subsp. paratuberculosis belongs to the Mycobacterium avium complex, (MAC) which includes several species of slow-growing mycobacteria that are isolated from environmental, animal or human samples. The two main species of MAC are M. intracellulare and M. avium. This latter can be subdivided into four subspecies: M. avium subsp. avium, M. avium subsp. silvaticum, M. avium subsp. paratuberculosis and M. avium subsp. homi- nissuis. Map strains are classified into three types (types I, II and III) on the basis of growth characteristics and different molecular typing. These types present host preferences but not host specificity. Type I and III strains (grouped in S ‘sheep’ type) share similar phenotypic traits and have been isolated mainly from sheep and goats and rarely from wildlife. The type II strains (or C ‘cattle’ type) are the most common in Europe. They were primarily isolated from cattle but have a very broad host range and have been isolated from small domestic ruminants, different wildlife species and humans. Type II is predominant in wild cervids. A ‘bison’ type has been described, but this type is currently included in type II. The complete genome sequence of Map K10 strain is published (GenBank accession number: AE016958). Map is an acid-fast, aerobic, non-spore-forming, non- motile bacillus (1 to 2 μm by 0.5 μm). In ZN-stained smears, the bacilli appear as short, acid-fast rods in clumps, indicative of intracellular growth. Like other mycobacteria, Map possesses a lipid-rich cell wall, thus resulting in acid fastness, hydrophobicity and increased resistance to chemicals such as chlorine. On the commonly used media, colonies are initially round, smooth and white. Map is a very slow-growing, fastidious organism, and many strains require the presence of mycobactin. Map is the slowest growing of the cultivable mycobacteria, with a generation time of over 20 hours, and visible colony formation on solid media may require up to 6 months. EPIDEMIOLOGY GEOGRAPHICAL DISTRIBUTION AND HOSTS Presence of Map in wildlife is documented in ruminant and non-ruminant species in several European countries. In Spain, a recent serologic survey described a widespread distribution of exposure to Map in red deer (Cervus elaphus)(50). A 2-year study (2002—2004) has shown increased cases of paratuberculosis in wild cervids in Austria(51). Map (or specific antibodies) has also been recorded in free-ranging populations in the Czech Republic, Slovakia, Italy, Greece, France, Belgium, Netherlands, Germany, Norway, the UK and Poland. Reports of occurrence in European ruminants show different results, depending on the country and the wild species studied. Comparison between countries is difficult, because some studies are large-scale serologic surveys and others are based on isolation and molecular characterization of Map from a few targeted wild animals. One study performed in free-ranging red deer in Spain reported a seroprevalence of 30%(50). Presence of Map or antibodies was also recorded in Italy and Spain in free-ranging red deer(52), fallow deer (Dama dama)(53), Cantabrian chamois (Rupicapra pyrenaica parva)(54), mouflon (Ovis aries musιmonf5' and ibex (Capra ibex)(56). In Norway, antibodies were detected in red deer (3.8%), roe deer (Capreolus capreolus) (12.2%), moose (Alces alces) (1.9%) and semidomesticated reindeer (Rangifer tarandus tarandus) (3.4%), whereas wild reindeer were seronegative(57). Red deer with antibodies to Map (2%) were also detected in the Czech Republic(58). In roe deer, a prevalence (by culture and PCR) of 22% was reported in Italy(52). In Europe, reports related to the presence of Map in roe deer are less frequent than in red deer. In North America, many studies demonstrate the presence of Map in a large range of free-ranging ruminants, including Rocky Mountain bighorn sheep (Ovis canadensis), Rocky Mountain goats (Oreamnos americanus), whitetailed deer (Odocoileus virginianus), Key deer (Odocoileus virginianus clavium), tule elk (Cervus elaphus nannodes) and bison (Bison bison)59- Paratuberculosis is primarily a disease of ruminants, but the presence of Map in several non-ruminant wildlife species (Table 20.4) underlines its multi-host infectivity and suggests that the epidemiology of the disease is complex. In lagomorphs, the presence of Map was suspected for the first time in a hare (Lepus europaeus) in the UK. Thereafter, several studies reported the presence of mycobacteria in wild rabbits (Oryctolagus cuniculus), first in Scotland, and then in other countries. The most relevant studies were undertaken in Scotland (in areas where paratuberculosis was endemic in domestic ruminants), showing a prevalence of 20 to 60% in subclinically infected rabbits(60). The most probable way of inter-species transmission is through ingestion of grass with contaminated TABLE 20.4 Wildlife host range in which Map has been detected in Europe. and rodents could cause Map contamination of food stored for cattle. A characteristic of paratuberculosis is the higher susceptibility of young ruminants. This is well known in cattle and farmed deer. Oral experimental infection with a high dose of Map of weaned calves, yearlings and adult red deer showed a strong age-related resistance of adults, which became infected but did not develop clinical or subclinical disease1-62). Genetic susceptibility to Map infection and/or disease is demonstrated in some breeds of cattle and sheep. Similar genetic studies have not been undertaken in wildlife. ENVIRONMENTAL FACTORS Map is an obligate intracellular pathogen of animals, but the organism can survive for long periods outside the host (in soil and water). Survival for more than 1 year was observed in a dry, fully shaded environment, and the survival decreased in locations exposed to direct sunlight. In favourable environments (high humidity, low pH and shaded areas), Map may survive for several months in pasture contaminated by shedding animals. Results of these environmental studies suggest that the major threat to Map survival is large temperature variations. The presence of dormancy-related genes have been demonstrated in the Map genome, and other potential survival mechanisms of the bacteria include biofilm formation and aero- solization. Moreover, some studies suggest that nematodes, insects or protozoa could play a role as vehicle for this mycobacterium(63). faeces from shedding cattle. Subsequently, several methods of transmission have been demonstrated between lago- morphs. The horizontal route (faecal-oral route) is the most frequent; however, isolation of Map in the placenta, fetus and milk suggest that vertical and pseudovertical (through suckling) transmission routes are possible in rabbits (61). These results demonstrate that Map may persist within populations of wild rabbits even in the absence of infected domestic ruminants. Among other wild species, Map has been detected in predators and scavengers. In the UK, a high prevalence (80%) has been detected in foxes (Vulpes vulpesfa't. The bacteria has also been isolated from the lymph nodes of stoats (Mustela erminea) and weasels (Mustela nivalis), from faeces of other mammals including rodents and wild boar (Sus scrofa), and from birds. Birds EPIDEMIOLOGICAL ROLE OF THE AFFECTED SPECIES Although Map has been isolated from many wild species, these may be classified into two categories according to their epidemiological significance for livestock. The first group includes wild ruminants and lagomorphs (especially rabbits),which may be considered as maintenance reservoirs. These wild species can maintain infection independently of the presence of infected domestic ruminants, and ecologically they share the same pastures. The second group are dead-end hosts; these are carnivores that may contract the infection but do not maintain it if the primary reservoir is absent. This group also includes some rodents and birds, which may disseminate Map passively in the environment. In this context, the transmission of Map from wildlife to livestock is assumed to be by faecal contamination of feed and forage, on pastures or in the barns. The interspecies transmission has been demonstrated experimentally; cattle may be infected with a rabbit strain of Map, and conversely. TRANSMISSION Map is predominantly transmitted through the faeco-oral route, via mycobacteria-faecal-contaminated colostrum, milk, water or feed. Moreover, contaminated colostrum and milk produced by the infected dam is the major route of transmission for neonates. The bacterium has also been isolated from semen from domestic cattle. Fetal infection is also possible in utero, and the rate of intrauterine transmission has been demonstrated to be higher in farmed red deer than in cattle or sheep. Carnivores may also be infected by eating an infected prey species. PATHOGENESIS, PATHOLOGY AND IMMUNITY After ingestion, Map enters the intestinal tissue through the M cells in the Peyer’s patches of the small intestine. Regional enterocytes may also be involved in the uptake of the bacterium. The bacterial cells are then phagocytosed by resident subepithelial macrophages or dendritic cells, in which they multiply1-64). Map is able to survive within the macrophages by inhibiting phagosome acidification and phagolysosomal fusion. This is a crucial process in establishment and evolution of the disease. With disease progression, infected macrophages migrate into local lymphatics, resulting in bacterial spread to mesenteric lymph nodes. The infection may be disseminated, with mycobacteria detectable in extra-intestinal lymph nodes (mammary, pulmonary, hepatic and head) and associated organs. Subsequent to infection, Map is initially controlled by a T helper (Thl) response with IFN-γ production. The progression of the disease from a subclinical to a clinical state is associated with a switch from a Thl to a Th2 immune response, leading to antibody production that is less effective in controlling infection. The loss of control in some hosts may be due to genetic factors or may be induced by external factors, including parturition, malnutrition or secondary viral or bacterial infections. The interplay between Thl and Th2 cytokines has been reviewed1-64). The host response to Map infection results in granulomatous lesions in the small intestine in which thickened mucosa and associated lymphadenopathy are characteristic. The proliferation of reactive lymphocytes, epithelioid macrophages and giant cells results in infused, blunted villi with decreased absorptive capacity. Malabsorption in the ileum, exudation from the inflamed intestinal mucosa, and lymphatic stasis all act to overload the absorptive capacity of the bowel, resulting in weight loss associated with a protein-losing enteropathy. Pathology in Wild Ruminants Cervids that die with terminal disease present with severe emaciation and faecal staining around the perineum and hind legs (Figure 20.5). Gross lesions and histopathology may be suggestive but may not distinguish between lesions induced by Map, M. bovis and M. avium avium. The gross lesions are not always present and early cases may be missed at post mortem examination. The lesions are mainly observed in the lower part of the jejunum, ileum, ileo- caecal junction and associated mesenteric lymph nodes (Figure 20.6). The classical picture is a diffuse thickening of the intestinal mucosa, where transverse folds may be observed. Frequently in wild cervids, the only gross lesion FIGURE 20.5 Adult female red deer with paratuberculosis, killed because of cachaexia and signs of diarrhoea. FIGURE 20.6 Cross-section of mesenteric lymph node from an adult male red deer found dead (severe emaciation and signs of diarrhoea), revealing extensive fibrosis and multifocal necrosis. Both mesenteric and mediastinal lymph nodes were enlarged and ZN- positive, PCR IS 900-positive and culture-positive for M. paratuberculosis. present is enlargement of mesenteric lymph nodes, with none or only mild intestinal-wall thickening. Lymph nodes are pale and oedematous with indistinct cortico- medullary junctions. Occasionally, they contain creamcoloured caseous exudate. Prominent lymphatic vessels may be observed as thickened cords in the mesentery leading to the mesenteric lymph nodes. Disseminated infection with lesions in the lung and liver has been reported in severe cases in farmed and free-ranging cervids. Microscopic changes are characterized by granulomatous inflammation in the mucosa of the small intestine and associated lymphatics and lymph nodes. Occasionally, focal granulomas may be observed in extra-intestinal tissues, especially liver, lung and associated lymph nodes. In affected tissues, focal or multifocal granulomas consist of macrophages, variable numbers of Langhans giant cells and mild infiltrations of lymphocytes. In severe cases, epithelioid macrophages may accumulate in lymph nodes and foci of necrosis may occur, but mineralization is rarely observed in wild cervids. As in domestic ruminants, acidfast organisms may be observed in variable numbers (pau- cibacillary and multibacillary forms) in macrophages and Langhans giant cells on ZN-stained sections. These two distinct forms are associated with two different immune responses. On the one hand, the multibacillary form (also named lepromatous form) is characterized by numerous intracellular bacteria and epithelioid macrophages. In this form, animals mount a weak cell-mediated immune response, but they develop serum antibodies. On the other hand, the paucibacillary form is associated with increased lymphocytes, a strong cell-mediated immune response and a poor antibody response. In animals with paucibacillary lesions, intracellular bacteria are present in lymph nodes or Peyer’s patches but not always in the intestinal mucosa. A histopathological grading system to differentiate these two forms of paratuberculosis in red deer has been developed1-65). Pathology in Non-Ruminant Wild Animals Typical lesions have only been observed in wild rabbits and in some carnivores. In rabbits, some studies reported thickening of the intestinal wall and enlarged lymph nodes(66) whereas others did not report these gross lesions(60). In severely affected wild rabbits, gross lesions may be observed in intestines, gut-associated lymphoid tissue, mesenteric lymph nodes and liver. They consist of chronic granulomatous inflammation with numerous macrophages and giant cells and many intracellular acidfast organisms(60). In fox, stoat and weasel, positive lymph node cultures associated with mild histopathological lesions suggest that these carnivores may be infected by Map. In comparison with ruminants and rabbits, the lesions in these carnivores are subtle, suggesting that these animals may be ‘dead-end’ hosts for Map. Similar mild histopathological lesions have also been observed in a crow (Corvus corone) and a wood mouse (Apodemus sylvati- cus))66°. In other species (Table 20.1), the isolation of mycobacteria was not associated with lesions, suggesting a simple passage of ingested Map through the intestinal tract(60). CLINICAL SIGNS AND TREATMENT Clinical paratuberculosis is infrequently diagnosed in free- ranging cervids, but it is more frequently observed in farmed red deer. In advanced clinical disease, animals are weak, emaciated and usually have chronic diarrhoea. This clinical sign is frequent in cattle but not always present in wild ruminants. Most studies report subclinical cases in which Map is isolated from apparently healthy cervids, or results of serological studies for which there is no information about the health status of the sampled wild animals. In Europe, some clinical cases have been observed in fallow deer in Spain)53), in red deer in Austria, Italy and Belgium1-24’67’68), in roe deer in the Czech Republic)69) and in Alpine ibex in Italy)56). In the majority of cases, these animals were found dead, emaciated and with rough coat hair. Emaciation is progressive and fatal, but the appetite is conserved, and the wild cervids remain alert until the terminal stages. It appears that the progression of paratuberculosis is much more rapid in wild cervids than in cattle and it is not uncommon to isolate Map from young red deer under 2 years old and even in fawns, which may be found dead and emaciated. In farmed deer, two clinical syndromes, both fatal, are described: sporadic cases in adults with progressive weight loss over a few months, and acute cases in young deer (8—15 months old) with profuse diarrhoea and rapid weight loss within a few weeks)70). Growth of antlers may be adversely affected. In nonruminant wildlife species, no clinical cases have been associated with Map isolation. Long-term treatment of paratuberculosis is not considered in individual animals, because antibiotic therapy is cost-prohibitive, and efficacious drugs are prohibited in food-producing animals. In vitro, Map strains are susceptible to drugs such as clarithromycin, azithromycin, amikacin and rifampicin. Diagnosis Diagnosis of paratuberculosis, especially subclinical cases, continues to be a challenge in domestic ruminants. Poor performances of current diagnostic tools are a major problem for the control of paratuberculosis in livestock. In wildlife, the diagnosis is also difficult because routine tests, especially serologic tests, used in domestic livestock are generally not validated for wildlife. Thickened mucosa of the distal small intestine and enlarged associated mesenteric lymph nodes are the most frequent lesions observed in infected wild cervids. However, gross and microscopic lesions may be suggestive of paratuberculosis but are not diagnostic. The confirmation is based on culture and/or PCR. For histopathology and isolation of Map, targeted samples are the lower part of the jejunum, ileum, ileo-caecal junction, and associated draining lymph nodes, including other intestinal segments, is advised. Direct microscopy of ZN-stained smears (faeces, intestinal mucosa or lymph nodes) is a rapid method but is not specific and has low sensitivity. Short, red-staining rods, characteristically in clumps, indicating intracellular growth, is characteristic of Map. The detection of single acid-fast rod bacilli is inconclusive, because they may be environmental mycobacteria. The isolation of Map by culture (from faeces or gut tissue) is considered as the post mortem diagnostic ‘gold standard’, and this technique is essential if further molecular typing is required. Cultures are time-consuming; however, although slow- and fast-growing Map strains are described and decontamination protocols may lead to false-negative results, the specificity is 100%. The recommended procedures for decontamination and culture are reviewed)71). Several solid media of choice may be used, all containing the iron-chelating growth factor mycobactin J: i) egg-based media such as Herrold’s egg-yolk medium and Lowenstein-Jensen medium; ii) serum-based medium such as Dubos medium; and iii) synthetic medium such as Middlebrook medium. On solid media, primary colonies of Map may appear from 5 weeks to 6 months after inoculation. The environmental mycobacteria appear more rapidly in the same media. For identification, a small inoculum of suspect colonies is subcultured on the same medium with and without mycobactin (dependence test). This criterion must be used with caution because the mycobactin requirement is not confined to Map. Compared with the culture on solid media, the liquid-culturebased BACTEC system reduces the time and is considered more sensitive)71). Molecular tools give higher sensitivity and faster results than culture. PCR tests may be used to identify specific mycobacteria in faeces or tissues and also to confirm identification of colonies. Several Map-specific genomic sequences such as insertion sequence IS900, f57, ISMav2, ISMap02, ISMap04 have been identified. Of these, IS900 is by far the most widely used target for PCR detection of Map. However, the presence of IS900-like sequences in non-Map mycobacteria has lead to caution in the use of IS900 and indicates the need to verify the result with DNA sequencing. In order to improve the specificity and to make differentiation between single and mixed mycobacterial infections, multiplex PCR have been developed with success)72’73). Among serological tests, ELISA are the most widely used in wildlife. Several ELISA kits are available and, for the majority, sera are pre-absorbed with M. phlei to increase specificity. However, these ELISA are not validated for wild species. For serology of clinically affected farmed deer, an IgG1 antibody ELISA using two antigens (Paralisa®, with sensitivity >80% and specificity >98%) has been developed in Australia1-74). Sensitivity of serological tests is lower during the subclinical phase. For free-ranging cervids, an ELISA has been developed in Spain(50). Tests of cell-mediated immunity such as the intradermal skin test and the lymphocyte transformation test provide poor specificity because of the widespread exposure of deer to environmental mycobacteria. The IFN-γ test is successfully used as an indirect diagnostic tool in the early stage of the disease in cattle. No commercial IFN-γ test for use in deer or other wildlife is currently available. MANAGEMENT, CONTROL AND REGULATIONS Control of paratuberculosis is difficult in wildlife. Several management measures related to the wild host and their habitats may be taken in combination to reduce the prevalence of the infection. However, if Map is established in a wild cervid or rabbit population, eradication of the disease is unrealistic. Management Practices Related To Host and Habitat If a point source for a paratuberculosis outbreak is detected in wild cervids, selective culling of clinically diseased animals (severe emaciation with/without diarrhoea) may be carried out to eliminate potential excretors and in this way prevent Map from spreading. In some countries, this procedure is legal and is carried out by hunters and forest rangers throughout the year, particularly in ‘hot spot areas’. Necropsy of the animals culled is mandatory to confirm paratuberculosis. Selective culling should be limited to focal outbreaks and caution must be taken to avoid the disruption of the social structure of the group, for example by causing dispersal of infected animals. Measures relating to control of host species density are also important. When wild cervids are artificially gathered under higher than natural densities, the risk of transmission of many pathogens, including mycobacteria, is increased. Campaigns may be carried out to inform stakeholders about the infectious risks inherently associated with disease and the overabundance of cervids(75), including the recommendation to reduce artificial feeding in the habitat. During hunting activities in hot spot areas, hunting pressure may be increased and hunters should be encouraged to arrange laboratory analysis of cervids that are found dead. Moreover, in all hunting areas, correct disposal of viscera must be organized to limit the sylvatic cycling of mycobacterial infections. The success of such measures is largely dependent on the participation of field sports authorities, who have an important role in the control of stock densities and also in the early detection of disease cases. Despite these measures, a proportion of undetected shedders will remain as a continuing source of infection. Paratuberculosis has persisted for over 20 years in bighorn sheep in Colorado, despite selective culling and increased hunting pressure in the infected area. As eradication is not possible, minimizing contact between wild and domestic herds, by managing separate areas of grazing, may be considered in some regions. Besides wild ruminants, Map is also extremely persistent in rabbit populations. In the UK, a simulation model of Map dynamics in rabbit populations demonstrated that repeated annual rabbit culls (>40%) over many years are more effective at reducing infection than single (not repeated) and massive (>95%) population reduction1-76). There are several live attenuated and killed Map vaccines for use in farmed deer. Some provide significant protection against clinical disease and reduce faecal shedding, but they do not prevent infection and they also interfere with several bovine tuberculosis diagnostic tests. Vaccination studies are not reported in wildlife. Paratuberculosis is included in the OIE listed diseases affecting wild animals. Annual notification is mandatory. PUBLIC HEALTH CONCERN Although not currently classed as zoonotic agent, Map is suspected to be involved in the pathogenesis of Crohn’s disease (CD) in humans. It is recognized that CD involves the interaction of at least three components: a genetic predisposition, an environmental trigger and an unregulated immune response. The involvement of Map in CD is supported by several factors, including: i) CD and paratuberculosis share clinical and histopathological similarities; ii) viable Map has been cultured from intestinal tissues, blood and milk of patients with CD; and iii) antibodies to Map antigen have been identified in the blood of Crohn’s patients. Moreover, recent meta-analysis studies indicate that there is an association between the presence of Map and CD, but whether this association is causal or coincidental is not known. Arguments against a role of Map in CD are that TNF-γ inhibitors, which would not be effective if Map were the cause of CD, are effective in treatment of CD, whereas anti-mycobacterial treatment has limited efficacy(77). In conclusion, the infection of humans with Map and possible association with CD remains a controversial issue. In some countries, precautionary control measures are taken in food sectors, as there is a probability that dairy and meat products are contaminated by this mycobacteria. Of major concern are the studies demonstrating that Map has been cultured from pasteurized retail milk and that Map may be present in drinking water(78). For wildlife concern, the decision to class Map as zoonotic agent or not will have an impact on the level of control in wildlife. SIGNIFICANCE AND IMPLICATION FOR ANIMAL HEALTH In the past 10 years, the prevalence of paratuberculosis in livestock appears to be gradually increasing worldwide, despite efforts in expensive control programmes in farm animals. In this context, the existence of wildlife reservoirs with the potential to transmit the infection to livestock may affect the success of these programmes. Several molecular epidemiological studies have demonstrated sharing of Map strains between wild and domestic animal species(79). However, the risk of transmission from wildlife to livestock has to be put into perspective. In an infected herd, the major problem is young calves infected by their mothers or congeners. Wildlife reservoirs may have epidemiological importance on farms that have eliminated all infected livestock from the premises or on farms free of paratuberculosis but located in the same geographic areas as infected farms. In Europe, some studies suggest that the presence of paratuberculosis in wild rabbits and cervids may represent a risk for livestock. However, dead-end hosts, such as predators and scavengers, are probably not high-risk factors for interspecies transmission. The implication of paratuberculosis for wild animal health is mainly for captive and endangered wildlife species. Since 1991, paratuberculosis has been responsible for deaths of numerous hoofed mammals housed at the Zoological Society of San Diego(80). For free-ranging ruminants, it is difficult to determine the effect of paratuberculosis on wild populations. In some populations, there is a high rate of subclinical infection and the effect of chronic infection is difficult to quantify. However, it is unlikely that wild populations are severely affected, because the mortality is generally low. The risks from paratuberculosis should be considered in reintroduction or translocation programmes. ACKNOWLEDGEMENTS The author acknowledges Dr Fabien Gregoire for taking the photographs and Julien Paternostre for the laboratory technical help.
Mammals Cervidae Red deer Cervus elaphus Roe deer Capreolus capreolus Fallow deer Dama dama Reindeer Rangifer tarandus Moose Alces alces Bovidae Mouflon Ovis aries musimon Ibex Capra ibex Chamois bgcolor=white>Rupicapra rupicapra Suidae Wild boar Sus scrofa Canidae Red fox Vulpes vulpes Mustelidae Badger Meles meles Stoat Mustela erminea Weasel Mustela nivalis Ursidae Brown bear Ursus arctos Muridae Brown rat Rattus norvegicus Black rat Rattus rattus Wood mouse Apodemus sylvaticus Common vole Microtus arvalis House mouse Mus musculus Leporidae European rabbit Oryctolagus cuniculus Brown hare Lepus europaeus Soricidae Lesser white-toothed shrew Crocidura suaveolens Birds Carrion crow Corvus corone Rook Corvus frugilegus Jackdaw Corvus monedula Black-headed gull Larus ridibundus European curlew Numenius arquata Ruff Philomachus pugnax Common cuckoo Cuculus canorus Savi’s warbler Locustella luscinioides Starling Sturnus vulgaris