TUBERCULOSIS, MYCOBACTERIUM BOVIS AND MYCOBACTERIUM CAPRAE INFECTIONS
In most cases, tuberculosis is a chronic progressive infectious disease with zoonotic potential and a wide host range, including many species of wild animals in Europe.
AETIOLOGY
Tuberculosis in terrestrial wildlife species within Europe is most frequently caused by Mycobacterium bovis.
Tuberculosis caused by M. caprae is emerging in cattle, affects humans and has been reported in wild boar (Sus scrofa) and red deer (Cervus elaphus) in Spain, and in red deer in the Alps(1). Mycobacterium tuberculosis the cause of human tuberculosis; it is adapted primarily to humans and is able to infect wildlife species, but is thought to only occur as a consequence of close contact with infected humans.Strain Diversity and Molecular Epidemiology
Members of the M. tuberculosis complex can be identified at the species level by growth characteristics and deletion or SNP typing. Further characterization of M. bovis (and M. caprae) strains is possible by direct variable repeat spacer oligonucleotide typing (DVR-spoligotyping) and can be refined by variable-number tandem-repeat typing (VNTR). Over 1,400 spoligotyping patterns are recorded in the Mycobacterium bovis Spoligotype Database website(2). Typing allows the identification of differences between host species and coincidences in strain occurrence, spatial and temporal changes in the M. bovis isolate community, and co-infections of a single host by different strains. Hence, molecular typing can be a valuable tool for epidemiological surveys at different spatial and temporal scales(3). Some types are very frequent and widespread, while others are rare. However, biological differences between strains remain largely unexplored. Up to 252 patterns have been recorded in a single country (Spain), and 12 out of the 15 most frequent types were present both in cattle and in wildlife(4).
Combining spoligotyping and VNTR, up to eight different M. bovis strains have been recorded in cattle and wildlife in one single locality (Donana National Park). In general in the UK, the genotypes found in wildlife are similar in both kind and frequency to the genotypes found in cattle in the immediate area.EPIDEMIOLOGY
GEOGRAPHICAL DISTRIBUTION IN EUROPE
Mycobacterium bovis infection of livestock has been eradicated or controlled in large parts of Europe, but remains a serious problem in areas where the presence of wildlife reservoirs of infection hampers eradication. The best- known wildlife reservoirs occur in the Eurasian badger (Meles meles), mainly in the British Isles, and in the Eurasian wild boar mainly in the Iberian Peninsula. Locally in southern England and in Iberia deer may be maintenance hosts in areas of high population density. Cases of tuberculosis in wildlife have also been recorded in other European countries, including areas considered free of bovine tuberculosis. In recent years, tuberculosis has been reported in red deer in Austria, France and Portugal. No cases occur presently in Iceland and Scandinavia, where bovine tuberculosis has been eradicated.
HOST FACTORS
Host Species
Infection has been recorded in many free-ranging mammals in Europe, including not only ungulates and carnivores, but also insectivores, rodents and lagomorphs (Table 20.1).
Most wildlife species exhibit low prevalence of infection and act as dead-end hosts. Some cases of spillover to endangered species, such as the Iberian lynx (Lynx pardi- nus), are of concern for wildlife conservation. In domestic animals, cattle are the main host, but goats, pigs and occasionally sheep and cats are also affected. Mycobacterium bovis infection of domestic cats was common in Europe in the past, before control measures of tuberculosis in cattle and pasteurization of milk were implemented. Tuberculosis constitutes a serious health and economic problem in animals in captivity, in farmed game species — particularly red deer, fallow deer (Dama dama) and wild boar — and in zoos and animal parks, where it affects ruminants and often south American camelids, which are susceptible.
Badgers
Badgers are mostly affected in regions of Ireland and the UK where bovine tuberculosis is endemic. The prevalence of tuberculosis in badgers in South West England can
TABLE 20.1 Some free-living wild mammals in which M. bovis infection has been confirmed in Europe since 1990.
| Common name | Species |
| European badger | Meles meles |
| Red fox | Vulpes vulpes |
| Feral cat | Felis catus |
| Iberian lynx | Lynx pardinus |
| Stoat | Mustela erminea |
| Western polecat | Mustela putorius |
| Feral ferret | Mustela furo |
| American mink | Mustela vison |
| Otter | Lutra lutra |
| Red deer | Cervus elaphus |
| Roe deer | Capreolus capreolous |
| Fallow deer | Dama dama |
| Reeve’s muntjac | Muntiacus reevesi |
| Sika deer | Cervus nippon |
| European bison | Bison bonasus |
| Eurasian wild boar | Sus scrofa |
| Common shrew | Sorex araneus |
| Wood mouse | Apodemus sylvaticus |
| Yellow-necked mouse | Apodemus flavicollis |
| Field vole | Microtus agrestis |
| Bank vole | Clethrinomys glareolus |
| Grey squirrel | Sciurus carolinensis |
range from 2% to 37%.
Badgers may excrete mycobacteria in sputum, urine and faeces or through draining abscesses and infected bite wounds. Badgers may remain infectious for many years, during which time they may contaminate pastures, or enter farm buildings and contaminate cattle feed. Tuberculosis is sporadically reported in badgers in Spain and France.Wild Boar
Tuberculosis in wild boar is widespread in Europe(5), with a prevalence of 1 to 52%. Prevalence is high in the southern Iberian Peninsula, where artificially high densities of wild boar (up to 90 per square km) and their aggregation at feeders or waterholes create ideal conditions for maintenance of the infection. In these circumstances, the Eurasian wild boar acts as a tuberculosis reservoir. Because of their high susceptibility and the ease with which they can be sampled, wild boar are also good sentinels of infection. Wild boar are able to breach fences and can act as links between farms and wildlife habitats.
Deer
Infection in red deer can last for years, making this host particularly relevant for maintenance and onward transmission of disease to other susceptible species. Local prevalence of up to 30% has been recorded. Tuberculosis in roe deer (Capreolus capreolus) is reported at low prevalence, of approximately 1%, particularly in areas where tuberculosis is endemic in cattle. Wild fallow deer can be important hosts where they are sufficiently abundant. Prevalence varies locally, and in the UK was estimated to be up to 5%, but with local foci of up to 20%(5). From 1997 to 2004 in a reserve in the Bieszczady Mountains in Poland tuberculosis was widespread in a herd of wild European bison (Bison bonasus)6'1, which subsequently had to be destroyed.
Carnivores in general (with the aforementioned exception of badgers) show a low susceptibility to develop severe disease and often develop mild forms of tuberculosis, with minimal or no grossly detectable lesions. Mycobacterium bovis infection in red foxes (Vulpes vulpes) with prevalence of up to 4% is reported in several areas of Europe where there is a high level of infection in wildlife.
Rodents are considered to be relatively resistant to M. bovis infection. Nonetheless, low prevalence of tuberculosis has been detected in some rodent species in endemic areas in the UK(7).Age and Sex
The prevalence of tuberculosis increases with age. Being male is a significant risk factor for infection in red deer. Among badgers, higher prevalence of infection has been recorded in adults than in cubs and in males than females. In addition, infected male badgers exhibit higher rates of mortality than females. Among wild boar, lung lesions are more frequent in juveniles, the dispersing age group. This could contribute to disease spread. There is also evidence that younger badgers may develop more severe forms of disease.
Social Group
Population structure can be an important determinant of disease distribution. In high-density badger populations, where individuals are clustered in territorial social groups, M. bovis infection has been observed to be aggregated in space and time(8). Infection risks in badgers may vary with social group structure and size, with inter-group movements being identified as a risk factor at both individual and group levels1-9). Belonging to an M. bovis-infected social group is a significant risk factor for infection in red deer and wild boar, but not for fallow deer(3).
Host Genetics
Variation in inherent resistance of the host to infection or to progression of tuberculous disease has been observed among individuals of the same species. This is better understood in humans and in mice models but has been little investigated in wild animals. Inbreeding and reduced genetic variability are risk factors for tuberculosis among wild boar and red deer. This may be due to the effect of specific immune-related genes rather than just individual heterozygosity.
Environmental Factors
Host aggregation at feeding or watering sites increases the risk of transmission. In the Iberian Peninsula, the north- to-south gradient in wildlife tuberculosis prevalence probably reflects a climate-mediated gradient of host aggregation, particularly during summer.
Temperature and humidity also affect the survival of mycobacteria in the environment. In high-density badger populations the concentration of badger excretory products at communal latrines may constitute potential sources of environmental contamination. The relatively constant temperature, high humidity and dark conditions in badger burrow systems (setts) may provide an environment in which M. bovis can persist.Transmission
Mycobacterium bovis is efficiently transmitted by aeroge- nous dissemination, so high population density and aggregation of hosts enhances opportunities for transmission. In badgers this is further facilitated by their occupation of setts, where they share confined air spaces. Oral infection from ingestion of contaminated feed or water is frequent under certain conditions, such as during artificial feeding of deer. Predators and scavengers may become infected by eating infected prey or carcasses. Another potential route of transmission may be via bite wounding, and it is suspected that this may lead to rapid haematogenous spread of infection in badgers. Male badgers tend to have a higher incidence of bite wounds than females. Not all infected host individuals have the same significance in transmission. Animals in a more advanced stage of disease progression may excrete greater numbers of mycobacteria and potentially act as ‘super-spreaders’, contributing disproportionately to disease transmission. However, any infected animal has the potential to transmit. The probability of transmission will depend on the number of viable bacilli present, the degree of containment within the lesion, and its anatomical location. The potential for infection of con- specifics or other species is related to host density, distribution and behaviour.
PATHOGENESIS, PATHOLOGY AND IMMUNITY
Infection by the respiratory route is common in most hosts. However, infection by the orophayryngeal route occurs frequently in wild boar and red deer. Inhalation of M. bovis is the most frequent source of infection. Low doses of mycobacteria can establish infection by the aerogenous route. Experimental studies in many species, including cattle, badgers, deer, mice and guinea pigs, show a dose of M. bovis of less than 10 colony-forming units (CFU), which is sufficient to cause infection by the respiratory route. Establishment of infection via the oral route requires a much higher dose of mycobacteria. Infection can also be acquired percutaneously, particularly through biting. Rarely, transplacental infection results from tuberculous endometritis of the dam. The genital route of infection is rare.
The inhaled mycobacteria that escape the mucociliary clearance of the upper respiratory passages gain access to the pulmonary alveoli, where they are phagocytosed by macrophages. These phagocytes attempt to kill the mycobacteria, but the virulent bacilli often persist. The bacilli multiply and destroy phagocytes, and new macrophages accumulate. A cell-mediated immune response is elicited in 10 to 14 days post-infection, which leads to the development of a granuloma. Macrophages carrying M. bovis enter lymphatic vessels and migrate to the lymph nodes, draining the primary lesion. This early focus may evolve into a progressive lesion or may undergo fibrosis and calcification, eliminating or restricting the infection. Foci may maintain a latent infection, which can become reactivated later in life, particularly if the immune defence is weakened. Reactivation causes secondary tuberculosis sometimes several years later or during old age. The lymph nodes draining primary foci usually also develop tuberculous lesions. From these lesions the mycobacteria spread by lympho-haematogenous dissemination, resulting in new foci in other organs. These are most often located in preferential sites, such as lymph nodes and lungs, but may occur in any organ — for example, mammary glands, uterus, liver, kidneys, joints and others. The tonsils are frequent locations of M. bovis infection, but often without gross lesions. Granulomas may erode through mucosae or walls of ducts, resulting in spread and implantation of mycobacteria on the linings, such as the nasobronchial or intestinal mucosae, or of ducts of salivary or mammary glands. Spread along cavities, such as pleural or meningeal may also occur. The miliary form of tuberculosis is characterized clinically by its rapid course and pathologically by numerous solid, white foci, 1 mm to several mm in diameter. It is produced by the haematogenous dissemination of numerous bacilli simultaneously. Depending on the pattern of spread, it can be limited to the lungs or it may affect organs such as the liver, kidneys, spleen and bone marrow.
Infection via the oral route results in uptake of the mycobacteria at the mucosal-associated lymphoid tissues, such as Peyer’s patches and tonsils. This results in development of lesions in the draining lymph nodes.
The propensity to develop severe disease varies among species. This is illustrated by the high frequency of severe tuberculous lesions observed in deer, whereas at the other end of the spectrum mild forms of tuberculosis, often with no gross lesions, occur in red foxes. The progress of the infection and the development of lesions are highly dependent on the host mounting a successful immunological response, which controls the multiplication rate of mycobacteria. The outcome of the tuberculous infection depends on a combination of host factors (i.e. the state of acquired immunity, immune competence and natural resistance, age, species, general condition, stress and concomitant infections) and mycobacterial factors (i.e. infection dose, route and virulence). In this way, individuals of any species, including those with lower propensity to develop severe disease, have the capacity to develop generalized or advanced forms of tuberculosis. In most cases the course of infection is dynamic and may result in intermittent shedding of mycobacteria. The tuberculous lesions may grow through the walls of ducts and cavities, eroding mucosae and structures and opening into the lumen of respiratory, digestive or urinary tracts, often excreting mycobacteria. These lesions may later become healed and circumscribed by a fibrotic capsule, thereby interrupting the shedding of mycobacteria to the environment. This may be followed by new cycles of growth and erosion, alternating with healing and containment, resulting in intermittent excretion of M. bovis, in a process which may last for years.
In most cases, tuberculosis progresses slowly. Over the course of months or years, the granulomas develop, grow in size and coalesce to form larger lesions. Simultaneously, there may be reseeding of mycobacteria, causing new lesions. Large areas of tuberculous bronchopneumonic consolidation may replace increasingly larger parts of the pulmonary parenchyma. Similarly, the tuberculosis lesions in other organs may also expand, leaving little healthy tissue and ultimately causing organ failure and death.
Tuberculosis has a remarkable spectrum of lesion sizes, from a barely detectable white focus (1 mm) to very large tubercles or completely consolidated lungs. Often, no gross lesions are observed in infected animals, but microscopic lesions may be found. This is termed ‘no visible lesion tuberculosis’ (NVL). A single gross lesion in a lymph node, particularly of the head or respiratory system, is often the only pathological finding in infected animals (reviewed in(10)). The lesions may be solid (with no necrosis) or may have a centre of coagulative necrosis or caseation, with or without mineralization, or liquefaction. The gross appearance of tuberculosis lesions varies among taxonomical groups (Table 20.2). The classical tuberculous granuloma is described in cattle as formed by macrophages (or epithelioid cells, which are elongated macrophages) surrounded by a zone of lymphocytes and Langhans giant cells, with a fibrotic capsule, central necrosis and mineralization. However, the morphology of granulomas shows considerable variation among species, with some exhibiting very different forms to cattle (Table 20.2). For example, in badgers the granulomas are formed essentially by whorls of epithelioid cells, with lymphocytes and no giant cells (Figure 20.1). Acid-fast bacilli (AFB) in ZN-stained histological sections are easily demonstrated when they are abundant, during active phases of the infection. Granulomas with few AFB still indicate potential infectivity, as a single AFB in a 1 cm2 area of granuloma indicates that a minimum of 2,000 AFB may be present in that particular granuloma. In many cases of confirmed M. bovis infection, AFB are difficult to find or not observed at all in the granulomas.
The immune response to infection is characterized by activation of T cells that produce a range of
TABLE 20.2 Summary of gross and microscopic tuberculous lesions in selected wild mammals.
| Species | Predominant location of tuberculous lesions | Gross appearance of tuberculous lesions | Histological features of tuberculous lesions | Comments |
| Badger (Meles | Lungs, tracheobronchial | White nodules, often small | Rounded granulomas formed | Pulmonary tuberculosis is |
| meles) | and mediastinal LN. | (1 mm to a few cm), with | by clusters of epithelioid | most frequent (60% of |
| Skin, (tuberculous bite | necrosis and calcification | macrophages and | cases). Often develop | |
| wounds) and regional | in large tubercles. | peripheral lymphocytes. | slowly progressive | |
| LN. Retropharyngeal, | Enlarged LN with | Discrete circumscribing | tuberculosis, resulting in | |
| parotid, mandibular, | necrotic or solid lesions. | fibrosis. Central necrosis | mild forms with small | |
| hepatic, renal and | Pulmonary consolidation. | and sometimes | lesions. The NVL | |
| mesenteric LN. Generalized disease in up to 5% of the cases. Kidneys. Liver. | Miliary tuberculosis. | mineralization in larger granulomas. Often difficult to find AFB. No giant cells. | presentation is frequent. Severe generalized disease is more frequent in young badgers. | |
| Eurasian wild | Characteristically in | Tubercles with thick fibrotic | Epithelioid cells granulomas. | Juvenile wild boar (between |
| boar (Sus | mandibular LN | capsule and necro- | Necrocalcific centre, | 12 and 24 months) |
| scrofa) | (92-100% of cases). | calcified center (Figure | surrounded by cellular | usually develop the most |
| Medial retropharyngeal LN, parotid and tonsils. Lung and thoracic LN (19-83%). Mesenteric LN. Spleen, liver and joints. Generalized lesions. | 20.2). Variable size, from 1 mm to 15 cm. Small tubercles (to develop | |||
| (Cervus | LN. Lungs, | foci, round tubercles | with central necrosis | severe lesions. Intestinal |
| elaphus) and | tracheobronchial and | 1-10 cm diameter with | sometimes with | tuberculosis is frequent in |
| fallow deer | mediastinal LN. | caseous necrosis and | calcification, peripheral | wild red deer. Thoracic |
| (Dama dama) | Mesenteric LN and | sometimes calcification. | zone of epithelioid | lesions are more frequent |
| ileo-caecal valve. Tonsils. Generalized lesions in head, abdomen and thorax. | Large abscesses with liquefied necrosis (Figure 20.3). Pulmonary tuberculous consolidation. | macrophages, Langhans or irregularly shaped giant cells, lymphocytes, plasma cells and neutrophils. Variable numbers of AFB, not always observed. | in fallow deer. | |
| Red fox (Vulpes vulpes) | Mesenteric LN. Lungs. | No gross lesions in most cases. A few foxes show gross lesions, small tubercles, often calcified. | NVL tuberculosis in the majority of cases. A few foxes develop severe disease. |
AFB - acid-fast bacilli; LN - lymph node(s); NVL - no visible lesion
pro-inflammatory cytokines and other mediators, typified by interferon gamma (IFN-γ) in particular. Antibodies are produced during infection with M. bovis but are generally easier to detect as the disease becomes more advanced.
CLINICAL SIGNS
Most infected animals do not exhibit clinical disease. Respiratory signs of disease such as coughing or nasal exudates may be observed in severe pulmonary tuberculosis. Tuberculous lesions in any organ or tissue can cause signs of organ dysfunction. Enlargement of superficial lymph nodes containing lesions may be evident, and in some cases form fistulas opening through the skin to drain purulent debris. This is frequently observed in red deer and wild boar. Tuberculous arthritis is observed mostly in wild boar. Slowly progressive tuberculosis in the terminal stages is manifested by progressive emaciation and lethargy. Miliary tuberculosis has a rapid and most often lethal clinical course.
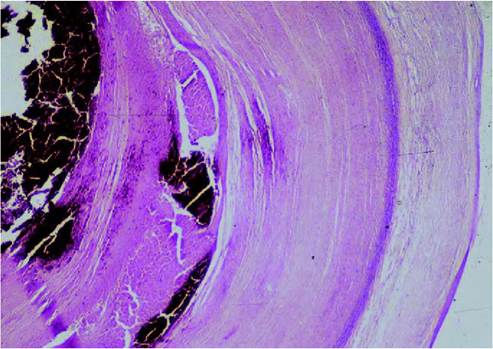
FIGURE 20.2 Histological section of an old tuberculous granuloma of a European wild boar, showing a thick fibrotic capsule and a necro- calcified centre. Photo: D. Gavier-Widen, SVA.
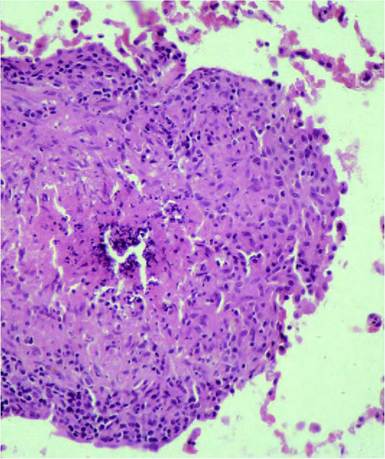
FIGURE 20.1 Badger lung with a small granuloma caused by Mycobacterium bovis infection. The granuloma is formed by macrophages/ epithelioid cells, with smaller numbers of lymphocytes in the periphery. There is necrosis and mineralization in the centre of the lesion. No giant cells are observed in tuberculous lesions in badgers. Haematoxylin and eosin. Enlargement 400?. Photo: D. Gavier-Widen, SVA.
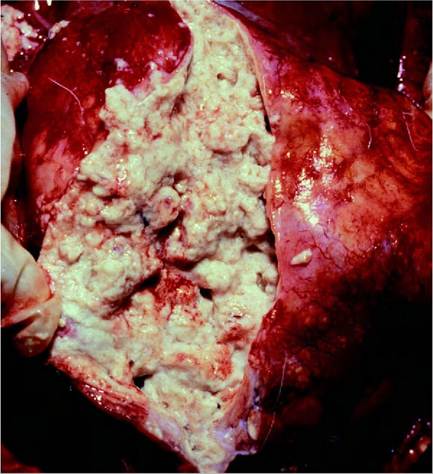
FIGURE 20.3 Bovine tuberculosis in a fallow deer. A large tuberculous abscess with caseo-purulent necrosis replaces lung structure. Photo: Bengt Ekberg, SVA.
DIAGNOSIS
Post mortem examination: a presumptive diagnosis may be based on the presence of gross lesions consistent with tuberculosis, but the definitive diagnosis requires the demonstration of M. bovis. Many cases present with only single or few lesions, which may be discrete. If these are not detected at necropsy and sampled for the demonstration of M. bovis, the diagnosis of tuberculosis most likely will remain unconfirmed. For this reason, to increase the chances of detecting lesions, a thin slicing of target lymph nodes and of lungs at necropsy is recommended.
Histopathology can improve detection of small tuberculous lesions and can contribute to the differential diagnosis. Identifying histological lesions typical of tuberculosis for the species and with the presence of AFB in ZN- stained sections support a presumptive diagnosis of tuberculosis but cannot differentiate between infections caused by M. bovis and those caused by other mycobacteria — for example, M. avium. The main differential diagnosis, grossly and histologically, is the parasitic granuloma, frequently found in mesenteric and respiratory lymph nodes and lungs of cervids and other wild hosts.
The isolation and culture of M. bovis from tissues obtained post mortem remains the ‘gold standard’ for the confirmation of tuberculosis. Detection rates are highest where macroscopic lesions are present, but often M. bovis may be isolated from tissues with NVL(10). For greatest sensitivity, samples for culture from NVL animals should include the most frequently affected lymph nodes in the species. In badgers these are the lymph nodes of the head, respiratory tract and digestive tract; in wild boar, they are the tonsils and mandibular lymph nodes; and in deer, they are the tonsils and medial retropharyngeal lymph nodes, ideally supplemented with the left bronchial and mediastinal lymph nodes and the mesenteric and ileo-caecal lymph nodes. Isolation of mycobacteria from samples taken from live animals, such as urine, faeces, exudates or lavage samples of upper respiratory tract frequently has low sensitivity as a diagnostic procedure, largely due to the intermittent nature of mycobacterial excretion1-11).
As well as its low sensitivity, mycobacterial culture is an expensive and labour-intensive process, and it can take up to 12 weeks to confirm a positive result. Bacterial culture has the advantage that M. bovis isolates are available for molecular typing using methods such as spoligotyping or mycobacterial interspersed repetitive unit-VNTR (MIRU- VNTR) typing, which may allow greater understanding of the epidemiology of the infection. For many wildlife species there are no proven diagnostic alternatives to bacterial culture.
The confirmation rate of tuberculosis in badgers was increased when more tissues were examined grossly and histologically, when a greater quantity and variety of tissues were cultured and when the culture period was extended1-12). This is likely to be true for other species.
Genetic probes can be used to reduce the time for culture confirmation, but most are only MTBC-group- specific. Polymerase chain reaction (PCR) offers the promise of faster and more specific detection of M. bovis from tissues, clinical and environmental samples. However, despite widespread use, a standardized, validated procedure for PCR detection of M. bovis does not yet exist, and culture has proven to be more sensitive than PCR for the detection of M. bovis from post mortem samples.
Diagnosis of tuberculosis is increasingly based on the detection of an immune response to M. bovis infection, either in association with, or as a replacement to, culture. The principal immunological response of the host to infection with M. bovis is the acquired cellular immune response, exemplified by the proliferation of lymphocytes and the production of cytokines such as IFN-γ. The mainstay of diagnosis of tuberculosis in cattle, the tuberculin intradermal skin test, is a method of detecting the cellular response in M. bovis-infected animals, but is impractical for free- ranging wildlife because of the need to examine animals for any cutaneous reaction 24—72 hours after the injection of tuberculin.
An increasing number of alternative ex vivo immunological tests are becoming available for the diagnosis of tuberculosis in wildlife. The optimum sensitivity of detection is generally seen through the measurement of IFN-γ by enzyme immunoassay following stimulation of whole blood or isolated peripheral blood mononuclear cells with tuberculin or specific antigens. Such a test has been reported for badgers and red deer(13), but in most cases the reagents involved are species-specific and would need to be generated for each species. Furthermore, for greatest success the test requires blood samples to be processed within 24 hours or less by a specialist laboratory, so in some settings the logistics or costs involved may make it unfeasible to operate the test.
Where low-cost, simple and rapid testing is required, immunological tests based on an antibody (serological) response may be more suitable. A wide variety of such tests have been published in the research literature for their application in numerous wildlife species (reviewed in (14)). Some enzyme-linked immunosorbent assay (ELISA) tests are available commercially for badgers, deer and wild boar. The principal limitation associated with serological tests in general is their relatively low sensitivity. There are exceptions to this, such as those developed for wild boar. Serological tests appear particularly suitable for detecting animals with advanced disease. For the monitoring of tuberculosis prevalence in wildlife populations, a combination of diagnostic tests may prove the most sensitive and cost-effective approach. For example, serology or lesion recording post mortem, combined with targeted application of more expensive but sensitive tools such as culture, PCR confirmation, or an IFN-γ test.
Although the presence of gross lesions consistent with tuberculosis is not an ideal tool for estimating prevalence of disease, such information is still valuable for exploring the magnitude and general distribution of infection in wildlife, provided a sufficient sample size is obtained from a large enough area.
MANAGEMENT, CONTROL AND REGULATIONS
Bovine tuberculosis is a reportable disease for the World Organisation for Animal Health (OIE); it is notifiable in wildlife species in some European countries and is included in the list of diseases in the eradication and monitoring programme of the EU.
However, tuberculosis in free-living wildlife has proven very difficult to control, and is probably impossible to eradicate. There are three main reasons to control tuberculosis infection in wild animals in Europe. Firstly, to prevent wildlife populations acting as a reservoir of infection for domestic animals; secondly, to reduce disease in wildlife hosts (particularly in protected and endangered species); and thirdly, to reduce the potential risks to public health. The principal approaches are reducing host abundance, which is usually achieved by culling, vaccinating host populations, or some form of management intervention to reduce opportunities for transmission either within wildlife populations or onwards to domestic species.
The approaches described here should not be considered in isolation, as the greatest positive impact may result from their use in combination. For example, in some species the effectiveness of a vaccination programme could potentially be increased by the addition of effective fertility control to curtail the recruitment of susceptible young animals. Alternatively, targeted culling may have a role in certain circumstances if employed with other measures such as vaccination and improved biosecurity of domestic animals.
CULLING
A large-scale UK field experiment to test the efficacy of badger culling in reducing transmission risks to cattle identified both negative and positive consequences(15). The outcome of culling operations may vary geographically, as a similar field trial in the Republic of Ireland identified only a net positive effect of badger culling on cattle herd breakdown rates(16). Such variation may potentially arise as a result of differences in culling techniques and strategies, and in the density and social behaviour of host populations. Ongoing developments in methods to diagnose tuberculosis and an improved understanding of tuberculosis dynamics in wildlife may increase the potential for targeted culling to be used. A potential alternative to culling wildlife populations is fertility control by immunocontraceptive vaccination, although this approach requires more research on immunocontraceptive agents, the demographic consequences of fertility control, and methods of delivery before its potential can be realized.
VACCINATION
Of the vaccines available, Mycobacterium bovis strain bacille Calmette-Guerin (BCG) is the tuberculosis vaccine of choice for use in wildlife in the near future and is currently being developed for use in a variety of domestic and wild animal species. BCG vaccination via subcutaneous, intramuscular and mucosal routes has been shown to confer significant protection to a number of wildlife species, including badgers, farmed red deer and wild boar. An obstacle to effective BCG vaccination of some wildlife populations is a practical means of delivering a stable vaccine preparation to target species in the field. A lipid matrix has been developed in New Zealand, which allows BCG bacilli to be maintained in a viable state suitable for oral delivery. Recent studies in badgers, brushtail possums and whitetailed deer (Odocoileus virginianus) have shown that oral vaccination with lipid-formulated BCG can induce protection against M. bovis that is comparable to that induced by injected vaccine. Specific baits have also been developed recently for the selective vaccination of wild boar piglets. To license BCG for use in wildlife it is necessary, among other things, to demonstrate efficacy against M. bovis, and the safety of a vaccine overdose to wild animals of the target species. These studies have been completed in the UK for BCG in badgers. In March 2010, a licence was granted for the use of BCG in badgers in the UK as an injectable (intramuscular) vaccine — ‘BadgerBCG’. This is the first licensed vaccine against tuberculosis in animals to be granted. It will take several years to generate data required to license oral BCG for wildlife in Europe, although work is in progress.
BIOSECURITY - REDUCING CONTACT BETWEEN LIVESTOCK AND WILDLIFE
Tuberculosis transmission between wildlife and domestic animals could be reduced if the two populations could be physically separated. Fences can be used successfully to control movements of larger mammals such as deer. Where transmission risks are concentrated in a particular location (e.g. badgers entering farm buildings), exclusion measures may be targeted accordingly. Exclusion of small mammals or those that burrow is more difficult. In game species, such as deer and wild boar, the management of spatial aggregations at supplementary feeding sites or waterholes, and the safe disposal of viscera by hunters could contribute to reducing tuberculosis transmission risks.
PUBLIC HEALTH CONCERN
The manipulation of live or dead wild animals may present a risk of infection for humans. In Spain, molecular typing of M. bovis showed that 6 out of 11 spoligotypes identified in wildlife were similar to those described in human cases. This suggests that hunters, wildlife managers or gamemeat consumers may be at risk of infection. Post mortem inspection of deer and condemnation of affected organs or whole carcasses markedly reduces the danger of infection by consumption of venison. Infection of semidomesticated cats and domesticated cats and dogs may represent direct zoonotic potential(5).The demonstration of human-to-human transmission of M. bovis indicates the potential public health risks of humans acquiring M. bovis infection*17).
SIGNIFICANCE AND IMPLICATIONS FOR ANIMAL HEALTH
The consequences of tuberculosis in wild animals include adverse effects on disease eradication in livestock, and adverse effects on wildlife management and conservation, as the disease can affect the productivity and density of wildlife populations with an economic or recreational value. The potential role of wild animals in the maintenance and spread of M. bovis infection in livestock represents the greatest economic impact of the disease in certain European regions. The disease is of particular importance in countries where eradication programmes have substantially reduced the incidence of tuberculosis in cattle but where disease persists and new outbreaks occur. Tuberculosis has an impact on conservation of endangered species. The critically endangered Iberian lynx, which may become infected through consumption of tuberculous carcasses, may be at particular risk.
ACKNOWLEDGEMENT
The authors acknowledge Noel Smith for the review on strain diversity and molecular epidemiology.
More on the topic TUBERCULOSIS, MYCOBACTERIUM BOVIS AND MYCOBACTERIUM CAPRAE INFECTIONS:
- TUBERCULOSIS, MYCOBACTERIUM BOVIS AND MYCOBACTERIUM CAPRAE INFECTIONS
- Mycobacterium bovis Infection in Humans
- Mycobacterium bovis Infection in Humans in Egypt
- Molecular Typing for Epidemiologic Studies of Mycobacterium bovis
- Genetic Diversity of Mycobacterium bovis Strains in Tanzania
- Tuberculosis Caused by M. bovis in Humans in Uganda
- 5.3 Maintenance Hosts of Mycobacterium bovis
- Mycobacterium spp. Infection: Tuberculosis, Paratuberculosis
- INFECTIONS BY OTHER MYCOBACTERIA OF THE M. TUBERCULOSIS COMPLEX
- The Mycobacterium avium Complex