The Mycobacterium avium Complex
5.3.1 Mycobacterium intracellulare
Through the 1940s and 1950s, numerous ‘atypical’ or ‘anonymous’ acid-fast pathogens were identified, including several responsible for serious human disease.
In one lethal case of disseminated disease in a young girl, acid-fast bacteria were found in multiple organs, and a mycobacterial infection was suspected (Cuttino and McCabe, 1949). Tuberculosis, leprosy, avian tuberculosis, rat leprosy and, owing to extensive intestinal involvement, paratuberculosis were all considered, but none perfectly matched the pathological features of the case. The causative agent also seemed morphologically distinct from known mycobacteria and was ultimately named Nocardia intracellularis. Another atypical acid-fast pathogen, dubbed the ‘Battey bacillus’ because of its prevalence at the Battey State Hospital (Rome, Georgia, USA), was associated with >300 of cases of chronic pulmonary disease (Corpe, 1964). Features of Battey-type pulmonary illness (e.g. cough, weight loss and lung pathology) closely resembled those of classic tuberculosis. However, the ‘Battey bacillus' was morphologically distinct from M. tuberculosis and harmless to guinea pigs. Epidemiological studies of the ‘Battey bacillus' indicated that it was not transmissible between people. More likely it was acquired from soil (Corpe, 1964). Classification of atypical mycobacteria was a key goal of the Veterans Administration-National Tuberculosis Association Cooperative Study of Mycobacteria (Runyon, 1958). Hundreds of isolates were collected, analysed and eventually divided into four major groups (Runyon, 1958, 1965). This work revealed that the Battey bacillus and N. intracellularis were the same organism. They were renamed M. intracellulare and placed in Group III (non-photochromogens), along with M. avium, M. gastri and M. terrae (Runyon, 1965, 1967; Wayne, 1966).5.2.4 The Mycobacterium avium- intracellulare complex
Traditionally, speciation of non-tuberculous mycobacteria was based on phenotypic characteristics such as pigmentation, growth rate, growth temperature and biochemical activities. Mycobacterium avium and M. intracellulare could be distinguished from other species of the Group III, slowly growing, non-photochromogenic mycobacteria by a positive tellurite test, and negative results for urease and Tween hydrolysis (Kent and Kubica, 1985). However, reliable differentiation of M. avium from M. intracellulare was not possible, even when using laborious procedures, such as cell wall lipid analysis, se- rotyping and classical animal infection experiments. In an attempt to resolve the taxonomy of Group III organisms, the International Working Group on Mycobacterial Taxonomy (IWGMT) employed a panel of 89 isolates, including 47 M. avium and M. intracellulare strains. For each culture, 292 phenotypic ‘characters' were tested. Of these, 118 proved useful and improved classification of several Group III species (Meissner et al., 19 74), but even this numerical taxonomy approach could not reliably resolve M. avium and M. intracellulare strains. Suggestions to reclassify M. intracellulare as an official subspecies of M. avium were never adopted (Wayne, 1966; Meissner et al., 1974; Runyon, 1974), but the concept of a M. avium-intracellulare complex (MAC) did emerge (Meissner et al., 19 74).
5.2.5 MAC in the molecular era
Molecular approaches to mycobacterial diagnostics and taxonomy have changed our view of the MAC. Even early nucleic acid- based typing methods, including DNA-DNA hybridization (Baess, 1983) and commercial DNA probes (Saito et al., 1989, 1990), could readily differentiate M. avium from M. intracellulare. DNA sequencingbased approaches, especially 16S rRNA gene sequencing and 16 S-23S internally transcribed spacer (ITS) sequencing, revealed many distinct sequevars (Frothingham and Wilson, 1993; Mijs et al., 2002) and hinted at the presence of species other than M.
avium and M. intracellu- lare. MAC now refers to a group of mycobacteria that exhibit overlapping phenotypic features and cause similar disease syndromes. Currently, MAC includes 12 validly published species: M. avium, M. intracellulare, M. chimaera, M. colom- biense, M. arosiense, M. vulneris, M. marseillense, M. timonense, M. bouchedurhonense, M. yongon- ense, M. paraintracellulare and M. lepraemurium (Fig. 5.2.). Insights from WGS data are redefining the taxonomy of the MAC: using sequencingbased criteria, MAC organisms share an average nucleotide identity (ANI) >85% and exhibit high sequence identity in targets commonly used for multilocus sequence analysis, including the 16S rRNA (>99.5%), hsp65 (>97.0%) and rpoB (>94%) genes (van Ingen et al., 2018). Because of their extensive genomic similarity, it has been proposed that M. yongonense and M. chimaera be reduced to the rank of subspecies, i.e. M. intracellulare subsp. yongonense (Castejon et al., 2018) and M. intracellularesubsp. chimaera (Nouioui et al., 2018), and that M. paraintracel- lulare represents a later heterotypic synonym of M. intracellulare (Nouioui et al., 2018). Genomic characterization of M. bouchedurhonense and M. timonense has been compromised by the absence of reference material. The strains deposited in culture collections do not match the description of the type strains of these species (Tortoli et al., 2017; van Ingen et al., 2018).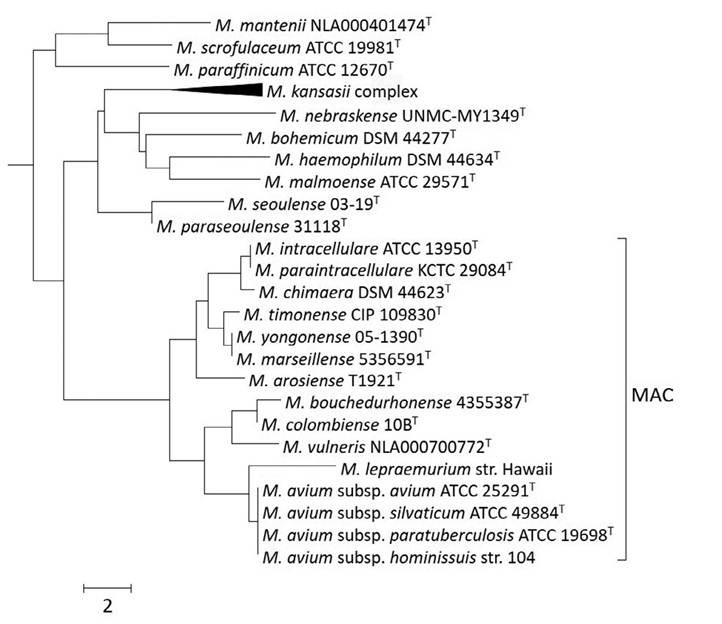
Fig. 5.2. Phylogeny reconstruction of ~1400 base pairs of the 16S rRNA gene using the neighbourjoining method and performed using the MEGA v7 software. The species shown represent the subcluster that included Mycobacterium avium complex (MAC) species in the context of a comprehensive 16S rRNA gene sequence alignment of all mycobacterial species. Scale represents the number of base pairs difference.
5.4
More on the topic The Mycobacterium avium Complex:
- Mycobacterium avium Complex
- Environmental Reservoir of the Mycobacterium avium Complex
- Mycobacterium Avium Complex Infection
- Mycobacterium avium
- Cultivation of Mycobacterium avium subsp. paratuberculosis
- Proteins and Antigens of Mycobacterium avium subsp. paratuberculosis
- Molecular Genetics of Mycobacterium avium subsp. paratuberculosis
- Infection of Non-Ruminant Wildlife by Mycobacterium avium subsp. paratuberculosis
- Mycobacterium avium subsp. paratuberculosis in Animal-Derived Foods and the Environment
- Drug Susceptibility Testing and Antimicrobial Resistance in Mycobacterium avium subsp. paratuberculosis
- Host-Pathogen Interactions and Intracellular Survival of Mycobacterium avium subsp. paratuberculosis
- Comparative Genomics and Genomic Epidemiology of Mycobacterium avium subsp. paratuberculosis Strains
- Chapter 6 The Mycobacterium tuberculosis Complex in Africa
- TUBERCULOSIS, MYCOBACTERIUM BOVIS AND MYCOBACTERIUM CAPRAE INFECTIONS
- Mycobacterium spp. Infections
- INFECTIONS BY OTHER MYCOBACTERIA OF THE M. TUBERCULOSIS COMPLEX
- Mycobacterium Tuberculosis
- VITAMIN B COMPLEX DISORDERS