Mycobacterium spp. Infection: Tuberculosis, Paratuberculosis
Tuberculosis
Rabbits are quite susceptible to experimental infection with Mycobacterium bovis and M. tuberculosis, but natural infection of domestic rabbits is now rare. Natural cases of tuberculosis in wild rabbits and hares have been reported, but are also rare.
In the early 1900s, infection among domestic rabbits was relatively common due to feeding unpasteurized milk as a supplement to young rabbits. Laboratory rabbits are valued as models of tuberculosis, since they are prone to develop cavitary pulmonary lesions (Fig 6.42), as well as dissemination to other organs. Extrapulmonary dissemination is particularly common when rabbits are infected with M. bovis. Lesions are typical of tuberculosis, consisting of granulomas with giant cell formation and large numbers of acid-fast bacilli. Rabbits with generalized thoracic and abdominal tuberculosis have been reported, which were naturally infected with M. avium. In a single report, a dwarf rabbit was found to have pleural effusion and pale foci in the lungs due to infection with M. genavense. Microscopic findings included severe intra-alveolar infiltration with foamy macrophages and mild to moderate nonsuppurative interstitial pneumonia. Rare acid-fast organisms were observed with the Ziehl-Neelsen stain. Speciation was performed by PCR, since M. genavense is difficult to culture.Paratuberculosis
Wild European rabbits are commonly infected with Mycobacterium avium paratuberculosis and are a significant reservoir for infection of livestock. Mild to severe histiocytic and granulomatous enteritis has been documented in wild rabbits involving primarily the small intestine, GALT, and mesenteric lymph nodes, as well as
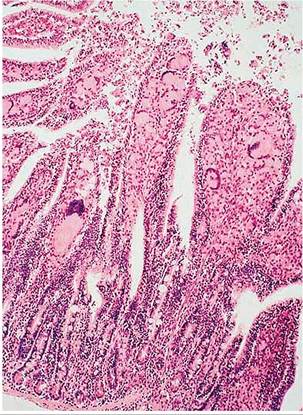
FIG. 6.43. Small intestine of a wild Oryctolagus rabbit naturally infected with Mycobacterium avium spp.
paratuberculosis. The lamina propria is densely infiltrated with histiocytes and multinucleated giant cells. (Source: Beard et al. 2001. Reproduced with permission from Elsevier.).periportal granulomas in the liver. The villus lamina propria and submucosa of affected areas were densely infiltrated with epithelioid macrophages and giant cells were prominent (Fig. 6.43). Foci of epithelioid macrophages were also present in the sacculus rotundus and appendix in the interfollicular zones at the base of lymphoid follicles. Acid-fast organisms were abundant in all lesions when tissues were stained with the Ziehl-Neelsen method (Fig. 6.44). Differential diagnosis includes Law- sonia intracellularis infection and accumulation of histiocytes in GALT lymphoid follicles. Definitive diagnosis can be achieved with acid-fast stains, culture, and PCR. Serology can be used for surveillance.
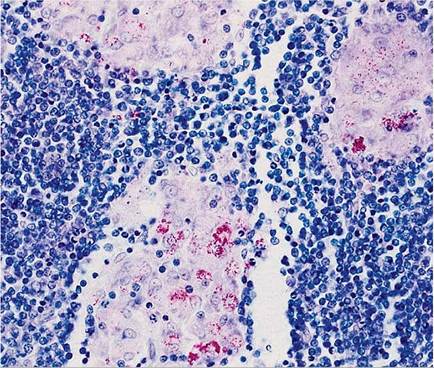
FIG. 6.44. Acid-fast organisms within histiocytes of the GALT of a wild rabbit naturally infected with Mycobacterium avium spp. paratuberculosis. (Source: Beard et al. 2001. Reproduced with permission from Elsevier.)
Pasteurella multocida Infection: Pasteurellosis; “Snuffles”
Pasteurella multocida is the most common bacterial pathogen of laboratory rabbits, and is a major cause of disease and mortality among pet rabbits and in many commercial rabbitries. Patterns of disease include purulent rhinitis, atrophic rhinitis, otitis media/interna, conjunctivitis, bronchopneumonia, abscessation, genital tract infections, abortions, neonatal mortality, and septicemia. For many years, P. multocida has been classified into a number of serotypes, based upon 5 capsular antigens (A, B, D, E, and F) and 16 serovars (formerly serotypes) based upon lipopolysaccharide (LPS) antigens. Historically, serovars 12:A, 3:A, and occasionally 3:D were the usual types found in rabbits with pasteurellosis, but serotype F originating from fowl cholera was recently discovered in rabbits in the Czech Republic.
A variety of molecular genetic approaches can now be used to detect and classify P. multocida.Epizootiology and Pathogenesis
The upper respiratory tract is the primary nidus of infection in affected rabbits. The organism can be spread by various routes from this site: to the lower respiratory tract by the aerogenous route; to the middle ear by the eustachian tube, hematogenously, or by local extension; to the external genital tract by venereal spread or nasal inoculation; and to other areas of the body by the hematogenous route or by local spread. In addition, infections of the upper and lower respiratory tract have been produced experimentally by subcutaneous or intravenous inoculations. Inapparent carriers of P. multocida are frequent, and even deep nasal swabs may fail to detect all animals carrying the organism in the nasal passages and nasopharynx on culture. Rabbits negative on nasal culture may be harboring P. multocida in the tympanic bullae, and the organism may be as readily recoverable from the middle ear as from the nasopharynx. Subclinical infections involving the tympanic bullae are relatively common. The organism may be acquired through various sources, including direct nasal contact with a shedder animal and contact with infected vaginal secretions. The vagina may serve as an important means of venereal spread to breeding males. Similarly, bucks may harbor P. multocida in their genital tract, and the organism may be transmitted to the does at mating. In young rabbits born to infected does, colonization with P. multocida may occur as early as 3 weeks of age. We have seen acute bronchopneumonia in kits as early as 3-4 weeks of age.
Aerosols do not appear to play a major role in the spread of infection in facilities with adequate air exchange. Housing and husbandry practices undoubtedly have a significant influence on the incidence of pasteurellosis in commercial and laboratory facilities. The reduction of air changes during colder months, poor sanitation, and overcrowding are all conditions that promote the elevation of the ammonia levels above a critical level of 25 ppm, increasing the likelihood of respiratory disease.
The role of fomites in transmission under field conditions is not known. Pasteurella is often recoverable from the watering nipples used by rabbits with snuffles. In view of the demonstrated adherence of type A P. multocida to rabbit pharyngeal cells, this may be one means of spread. The possibility of interspecies transmission is an important consideration. Strains of P. multocida from other species (bovine and avian isolates) have been passed in mice and rabbits. Following conjunctival inoculation of New Zealand White rabbits with either of the strains, acute pasteurellosis occurred. Experimental inoculation of serotype F (fowl cholera) resulted in fibrinopurulent or hemorrhagic pneumonia by intranasal route in 3-6 days and septicemia by subcutaneous route in 2-3 days. Peroral challenge showed no clinical signs of disease or macroscopic lesions. In the laboratory setting, experimental manipulations may enhance susceptibility to the disease, particularly procedures that may have an impact on the immune system. Conversely, it is often difficult to consistently produce significant lesions in unmanipulated “healthy” rabbits inoculated intranasally with virulent strains of P. multocida.Pathology
The most common clinical manifestations of pasteurellosis are rhinitis and vestibular syndrome. Chronic rhinitis, with catarrhal to mucopurulent exudate, is associated with the typical upper respiratory tract form of the disease. When performing a necropsy, it is essential that the nasal bones overlying the turbinates be removed to provide adequate exposure for a thorough examination of this area. Turbinate atrophy has been observed in rabbits naturally or experimentally infected with a 12:A serotype of P. multocida (Fig. 6.45). The prevalence of this manifestation has not been determined. Upper respiratory infections are often accompanied by conjunctivitis and otitis media, involving one or both tympanic bullae. In many cases, there may be no clinical evidence of middle ear infection; thus, it is essential that the tympanic bullae be opened and examined at necropsy.
Grossly, the affected middle ears contain a white or dull yellow to gray, thick, viscid exudate (Fig. 6.46). The lining of the tympanic bullae is light tan and opaque. On microscopic examination, there is often squamous metaplasia of the epithelium lining the tympanic bulla, with leukocytic infiltration into the submucosa and tympanic cavity. Rupture of the tympanic membrane may also occur, with extension into the inner ear and occasionally the brain.Pulmonary lesions arise from extension of upper respiratory infections or by the hematogenous route. Bronchopneumonia may vary from localized cranioven- tral involvement to acute necrotizing fibrinopurulent or fibrinohemorrhagic bronchopneumonia (Fig. 6.47).
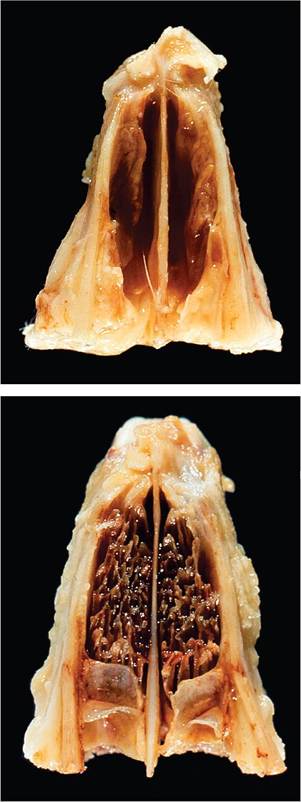
FIG. 6.45. Turbinate atrophy associated with chronic rhinitis due to Pasteurella multocida infection. Note the loss of turbinates in the affected (top) compared to the normal control rabbit (bottom). (Source: DiGiacomo et al., 1989, reprinted with permission from American Veterinary Medical Association.)
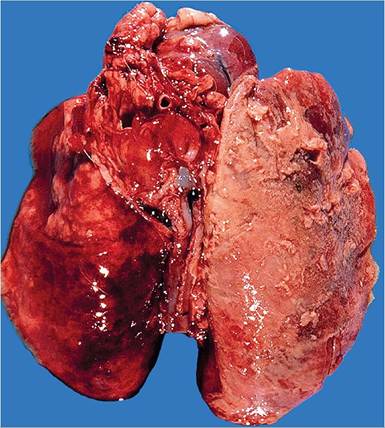
FIG. 6.47. Fibrinohemorrhagic bronchopneumonia and pleuritis in a rabbit with peracute pulmonary pasteurellosis. Fibrinous exudate is present on the pleural surface.
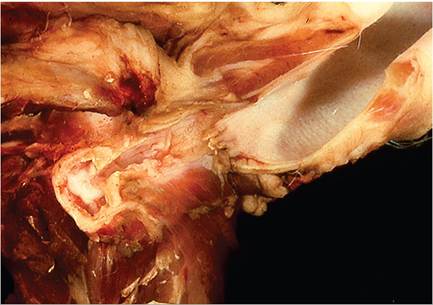
FIG. 6.46. Suppurative otitis media in a rabbit with chronic Pasteurella multocida infection. Purulent material is present within the opened tympanic bulla and the wall of the bulla is thickened.
Affected lung tissue in the acute necrotizing form is swollen and moderately firm, frequently with concurrent fibrinous pleuritis and pericarditis. In some cases, pulmonary lesions may be confined to one lung lobe. Chronic disease may encompass an entire lung lobe, with fibrinopurulent pleuritis, pericarditis, and empyema.
Microscopically, in animals with localized lower respiratory tract disease, the lesions may vary from chronic bronchitis with peribronchial lymphocytic infiltration to alveolitis with infiltrating leukocytes, heterophils predominating. In the acute, necrotizing form of the disease, destruction of alveoli and small airways, alveolar flooding with fibrinous exudate and erythrocytes, and infiltration with heterophils are typical findings. Multinucleated giant cells may be present in affected alveoli. Hematogenous infection of the lung may also occur, with formation of single or multiple abscesses.Macroscopic findings in the female reproductive tract may include pyometra, with chronic suppurative salpingitis and perioophoritis or localized suppurative lesions. Acute, transmural, necrotizing metritis has been associated with peracute pasteurellosis. This form occurs during the perinatal period, presumably due to invasion via the dilated cervices. Abortions and stillbirths may precede the death of the doe. Affected does usually die within a few hours after showing signs of disease. At necropsy, fibrinous exudate may be adherent to the uterine serosa, with ecchymoses. The uterine wall is thickened and contains necrotic material. Microscopically, there is acute, necrotizing transmural metritis and serositis, with changes in other organs consistent with a bacterial septicemia. In addition, chronic suppurative metritis and pyometra may also occur (Fig. 6.48). In
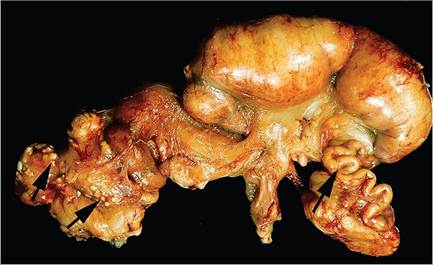
FIG. 6.48. Pyometra in an adult doe associated with chronic Pasteurella multocida infection. There is asymmetric distention of the uterine horns, and the fallopian tubes are distended with purulent exudate (right arrow). Small abscesses are scattered on the serosa (left arrows).
bucks with genital tract lesions, there may be suppurative orchitis with abscessation.
Circumscribed abscesses containing thick, yellowgray exudate may involve various sites, including subcutaneous tissue, mammary glands, brain, lung, visceral organs, and bone. Abscesses involving the jaw are particularly difficult to treat, since they tend to excavate bone and are difficult to drain because of the viscid nature of pus in this species. Localized infections may spread to other sites, resulting in manifestations such as vegetative endocarditis.
Acute septicemia is another possible manifestation of the disease. Affected animals may be found dead, with no previous evidence of disease. There may be a concurrent rhinitis and/or otitis media evident at necropsy, but frequently there is no gross evidence of lesions at necropsy. Microscopically, there should be changes consistent with an acute bacterial septicemia, including hemorrhage and variable thromboses of small vessels. Acute suppurative meningoencephalomyelitis may also occur, occasionally with concomitant optic neuritis and iritis.
Diagnosis
Definitive diagnosis is best achieved with bacterial culture. In cases of suspected septicemic pasteurellosis, the organism should be recoverable from a variety of parenchymatous organs and heart blood. Nasal cultures, although useful in identifying carriers/shedders of the organism, will not necessarily detect all infected animals. ELISA has been used to serologically identify animals that were consistently negative for Pasteurella on deep nasal culture. PCR can be used for detection, but molecular methods are more useful for characterizing isolates. Differential diagnoses of suppurative lesions or respiratory infections include other pyogenic infections, such as Staphylococcus, Bordetella, and rarely Klebsiella.
More on the topic Mycobacterium spp. Infection: Tuberculosis, Paratuberculosis:
- Infection of Non-Ruminant Wildlife by Mycobacterium avium subsp. paratuberculosis
- TUBERCULOSIS, MYCOBACTERIUM BOVIS AND MYCOBACTERIUM CAPRAE INFECTIONS
- Mycobacterium spp. Infections
- Mycobacterium Tuberculosis
- Cultivation of Mycobacterium avium subsp. paratuberculosis
- Proteins and Antigens of Mycobacterium avium subsp. paratuberculosis
- Molecular Genetics of Mycobacterium avium subsp. paratuberculosis
- Mycobacterium avium subsp. paratuberculosis in Animal-Derived Foods and the Environment
- Chapter 6 The Mycobacterium tuberculosis Complex in Africa
- Drug Susceptibility Testing and Antimicrobial Resistance in Mycobacterium avium subsp. paratuberculosis
- Host-Pathogen Interactions and Intracellular Survival of Mycobacterium avium subsp. paratuberculosis
- Comparative Genomics and Genomic Epidemiology of Mycobacterium avium subsp. paratuberculosis Strains
- Chapter 22 Holes and Patches: An Account of Tuberculosis Caused by Mycobacterium bovis in Uganda
- Malassezia spp. Infection: Malasseziasis
- Bacterial Enteric Infections Brachyspira spp. Infection
- Aspergillus spp. Infection: Aspergillosis
- Mycobacterium Avium Complex Infection
- Actinobacillus spp. Infection
- Actinomyces spp. Infection