Bacterial Enteric Infections Brachyspira spp. Infection
Wild rodents, including house mice, have been shown to carry a wide variety of Brachyspira spp., including pathogenic porcine and avian Brachyspira spp., and mice are experimentally susceptible to infection.
In addition to known Brachyspira spp., wild mice have also been found to carry slow-growing and weakly hemolytic spirochetes that represent a novel species within the genus Brachy- spira. Infection among laboratory mice has not been reported, but the authors of this book have observed natural infection in NSG mice (Fig. 1.48). In addition to culture, Brachyspira organisms can be detected and spe- ciated by PCR.Citrobacter rodentium Infection: Transmissible Murine Colonic Hyperplasia
Citrobacter rodentium causes a syndrome in mice called transmissible murine colonic hyperplasia (TMCH). It has also been termed “hyperplastic colitis," “catarrhal enterocolitis,” and “colitis cystica.” Unlike most Citrobacter spp., pathogenic mouse isolates are nonflagellated and nonmotile. Isolates from different outbreaks possess similar sugar fermentation and other biochemical profiles, but minor differences exist. The causative agent was formerly classified as Citrobacter freundii but has been reclassified as C. rodentium. TMCH promotes the
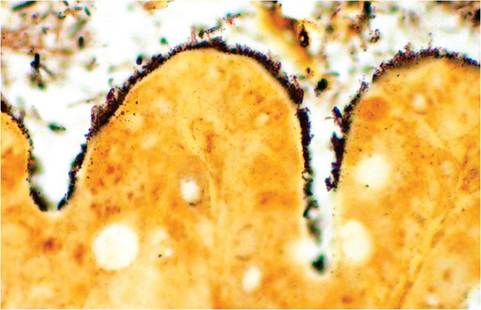
FIG. 1.48. Brachyspira spp. infection of the intestine in a NSG mouse. Note the numerous bacteria attached to the brush border. (Warthin-Starry stain).
evolution of preneoplastic and neoplastic change during chemical carcinogenesis and in Apc+/Mm (Min) mice. Citrobacter rodentium has gained popularity as a model for the pathogenesis of attaching and effacing E. coli, as it possesses an analogous genetic pathogenicity island. Thus, there is increased potential for iatrogenic introduction of this pathogen to laboratory mouse populations.
Epizootiology and Pathogenesis
TMCH has been reported in laboratory mice in the United States, Europe, and Japan, but its prevalence in wild mice is unknown. Citrobacter rodentium appears to be highly species-specific in its host range. When present in a mouse population, it is associated with disease. The bacterium does not establish itself as permanent microflora and is not subclinically carried by mice. Its source of introduction into mouse colonies is presumably through contaminated food or bedding. The organism spreads slowly among mice, requiring direct contact or fecal contamination. Following oral inoculation, C. rodentium transiently colonizes the small intestine, and then selectively colonizes the cecum and colon within 4 days. The bacteria intimately attach in large numbers to surface mucosa of the descending colon, displacing other aerobic bacteria. Bacterial attachment is facilitated by a genetic pathogenicity island encoding bacterial intimin and type III secreted bacterial proteins, including the translocated intimin receptor (Tir). These proteins induce dissolution of the brush border, actin filament rearrangement, and pedestal formation by the plasma membrane (Fig. 1.49), similar to attaching and effacing enteropathogenic E. coli (EPEC) and enterohemorrhagic E. coli (EHEC). Through undefined mechanisms, bacterial colonization elicits an intense mucosal epithelial hyperplasia. The acquired immune response is important in clearance of infection but also contributes to
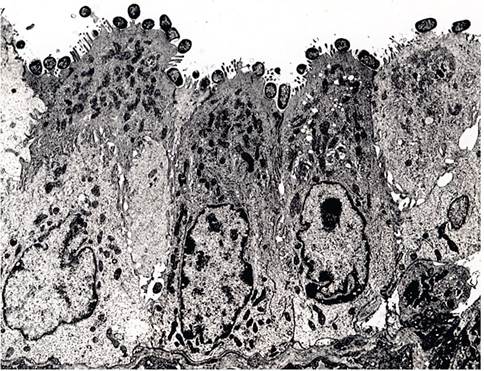
FIG. 1.49. Electron micrograph of Citrobacter rodentium attached to colonic enterocytes. Note the intimate attachment of bacteria with disruption of the brush border. (Source: Johnson et al. 1979. Reproduced with permission from Elsevier.)
inflammation and morbidity. As hyperplastic cells migrate to the surface, they displace infected cells, which are exfoliated from the surface.
Peak hyperplastic response occurs within 2-3 weeks, at which point the causative agent may no longer be isolated from the colon. Clinical signs are most prominent at this interval. Young mice and certain mouse genotypes (C3H substrains, AKR, and FVB) tend to develop more severe disease with varying mortality due to secondary inflammatory and ulcerative lesions in the hyperplastic mucosa. DBA/2, BALB/c, B6, and NIH Swiss mice are susceptible to hyperplasia, but tend to develop less inflammation. In the ensuing weeks, lesions regress with a period of excessive goblet cell differentiation and development of cryptal cysts that can fill with mucin. By 2 months, the mucosa returns to normal. There is no known carrier state, and recovered mice are refractory to reinfection.Studies have dissected the role of host immune response in controlling infection and in modulating the severity of disease, using a variety of GEMs with specific immune defects. Innate defenses, including beta defensins, IL-12, and IFN-gamma, affect early colonization and growth of the bacteria. Infection stimulates an acquired immune response, with recruitment of CD4 T cells into the mucosa, and the evolution of a Th1- polarized T-cell-dependent systemic antibody response. T-cell-dependent serum antibodies (IgM, IgG2c, or IgG2a), but not secretory IgA or IgM, are involved in clearance of infection and recovery, and this response does not require T- or B-cell responses in gut-associated lymphoid tissue. Effective immunity requires CD4 T cells and B cells but not CD8 T cells. The acquired immune response is necessary for clearance, but it is also a major factor in disease severity, since hyperplasia and inflammation are more severe in immunocompetent mice compared to Rag-1 null mice. Mice devoid of B and T cells (Rag- 1 null), CD4 T cells, or B cells (μMT null) fail to clear infection and succumb from sepsis, with bacteremia arising from both C. rodentium and other gut bacteria.
Pathology
Affected mice may be runted and have sticky, unformed feces that smear the cage walls.
Rectal prolapse often occurs (Fig. 1.50). Careful examination of the bowel will reveal a contracted, thickened, opaque, descending colon devoid of feces (Fig. 1.51). Lesions can extend into the transverse colon. The cecum is also frequently but variably involved. During the early stages of infection, the brush border of surface mucosa of affected bowel is heavily colonized by a carpet of intimately attached cocco-bacillary bacteria. As the lesion progresses, these infected cells are pushed aside by uninfected, hyperplastic epithelium, with retention of cells at the extrusion zones (Fig. 1.52). Inflammation and erosion can also occur, especially in infant mice or mice of certain genotypes or immune status, as discussed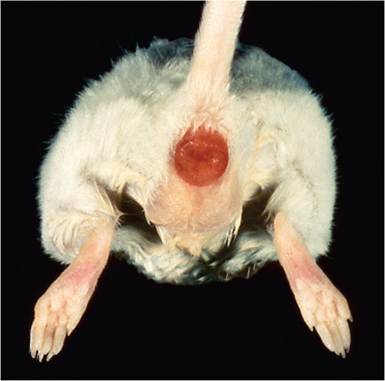
FIG. 1.50. Rectal prolapse in a mouse with transmissible murine colonic hyperplasia due to Citrobacter rodentium.
above. In mice that are incapable of mounting effective immunity, mortality occurs from bacteremia, with multifocal hepatitis and splenitis. As the hyperplasia regresses, cells can undergo differentiation into excessive numbers of goblet cells, and crypts can become distended with mucin and cellular debris (colitis cystica). Once regression is complete, the mucosa returns to normal. Secondary focal nonspecific ischemic necrosis and inflammation may arise in the liver during early infection.
Diagnosis
Infection is transient with no carrier state in immunocompetent mice. The causative agent is often absent when clinical signs are most apparent. Infection can be localized to the descending colon, and only a small percentage of mice in the population may be infected because of its low contagiousness. Isolation is enhanced
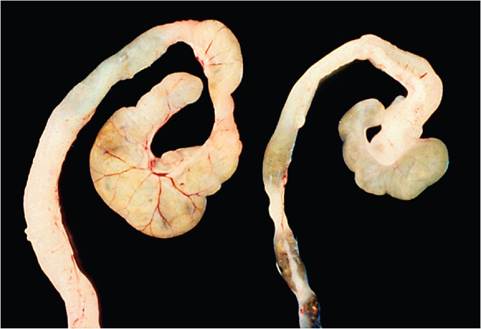
FIG. 1.51. Cecum and colon of a mouse infected with Citrobacter rodentium (left) compared to a normal mouse (right). The infected bowel is contracted, opaque, and devoid of feces. (Source: Barthold et al. 1978. Reproduced with permission from SAGE Publications.)
52" class="lazyload" data-src="/files/uch_group75/uch_pgroup311/uch_uch7290/image/image052.jpg">
FIG. 1.52. Descending colon from a mouse infected with Citrobacter rodentium. There is marked hyperplasia of the crypt epithelium with retention of cells on the surface and leukocytic infiltration in the lamina propria.
by culturing feces or descending colon from multiple mice in the early stages of infection before clinical signs are evident. Citrobacter can be readily isolated on Mac- Conkey agar. Differential diagnoses include other agents that cause hyperplastic colitis, including E. coli and Helicobacter spp. Hyperplastic colitis has also been observed in nude mice chronically infected with enterotropic MHV. Rectal prolapse is frequently associated with TMCH, but can also occur spontaneously or in association with colitis of other causes.
More on the topic Bacterial Enteric Infections Brachyspira spp. Infection:
- Actinomyces spp. Infection
- BIBLIOGRAPHY FOR BACTERIAL INFECTIONS
- Aspergillus spp. Infection: Aspergillosis
- Enterococcus spp. Infection: Enterococcal Enteropathy
- BACTERIAL INFECTIONS
- BIBLIOGRAPHY FOR BACTERIAL INFECTIONS
- Malassezia spp. Infection: Malasseziasis
- Brachyspira infections in birds
- COVER IMAGE ACKNOWLEDGEMENTS
- suttonella ornithicola sp. NOV. INFECTIONS OF SPECIES OF TIT AND LONG-TAILED TITS