Bacterial Culture and Typing Procedures
Culture is considered to be the diagnostic gold standard for the detection and confirmation of BTB in livestock and wildlife (Liebana et al. 2008; Gavier-Widen et al. 2009). Although it is considered as such, it too is neither 100% Se nor 100% Sp, and this adds to the difficulties of eradicating BTB.
The sensitivity of culture to detect mycobacteria is influenced by a number of factors, including the collection of appropriate specimens, the time taken before the specimens are processed for culturing, the way in which specimens are processed, and the culture media used (Gormley et al. 2014). Using conventional culturing techniques delay making a final diagnosis because of the slow growth of many of the mycobacteria, as it may take up to 15 weeks or more before some of the bacteria become detectable and available for species identification and typing (Corner et al. 2012). As is the case in postmortal examination of animals for the detection of lesions, the inability to culture mycobacteria from submitted specimens does not imply that the specific animal did not contain viable mycobacteria of the MTC, particularly when there were cases with early infection and in the preclinical stage of the disease, or latent carriers (Gormley et al. 2014).That bacterial culture is not 100% sensitive, is due to a number of factors, and they should be kept in mind when collecting and processing specimens for culture (Ramos et al. 2015). Tissue specimens for diagnostic culture should contain visible lesions if there is to be any likelihood of mycobacteria being cultured. If the intention is to detect non-visible lesions, adherence to specific protocols outlining the range of tissues to collect and to maximize the possibility of detecting mycobacteria should be strictly followed and adapted according to the species dealt with. Some of the causes of failure to culture the organisms include:
• The type of culture medium and O2 concentration during culturing: Three types of media are commonly used in Africa for culturing mycobacteria.
These are egg-based media such as Lowenstein-Jensen (LJ) or Stonebrink’s (SB), agarbased media such as Middlebrook (7H10 and 7H11) for primary isolation, and liquid media such as Middlebrook 7H9. All these media contain malachite green, a selective dye required for the growth of mycobacteria in culture (Buxton and Fraser 1977). Details of the techniques are available in guidelines available from the OIE and in various other publications (O’Brien et al. 2008; Mohamed et al. 2011). None of these media has 100% Sp, and hence attention to detail, including specimen collection and decontamination procedures in particular, are important processes that should be addressed, also in the future when developing improved culturing systems (Gormley et al. 2014).• The use of automated liquid culture systems such as the BACTEC 460 and 12B, BACTEC MGIT 960, and VersaTREK systems is an improvement on solid media, as it cuts down the average time for first detecting growth to as short as 15.8 days (Gormley et al. 2014). This technology is probably beyond the reach of most of the African countries because of cost and the lack of infrastructure and adequately trained human resources.
• There are two important factors that determine growth of M. bovis on culture. Glycerol, which is used as the primary carbon source in culture media used for mycobacterial culture, promotes the growth of several species of mycobacteria including M. tuberculosis, but inhibits the growth of M. bovis. It is thus critical to use media with an alternative carbon source such as pyruvic acid, to enhance the growth of M. bovis on culture (Collins and Grange 1983). Furthermore, contrary to most mycobacteria that are aerobic organisms, M. bovis is microaerophilic, and the correct environmental O2 concentration should be maintained when they are cultured (Addo et al. 2007).
• Depending on the culture medium used, and the pre-culture processing, too few organisms may be present in the specimens to allow positive culture (Anon 1994; Collins et al.
1994).• Using an incorrect protocol for the purpose of the investigation when collecting specimens for culture from individuals and groups of animals (Crawshaw et al. 2008).
• Not collecting a sufficient amount of tissue, organs, or exudate for culture. This applies particularly to milk, where about 100 ml from each quarter is recommended to optimize the likelihood of culturing the organism for typing purposes (Kleeberg 1984; Ben Kahla et al. 2011).
• The way in which specimens are collected, stored, and transported and the time involved in getting the specimens to the diagnostic laboratory. Tissues should be collected aseptically and processed immediately for culture. Samples stored at 4-6°C must reach the laboratory as soon as possible and must be processed within 24-48 h after collection; otherwise, they should be frozen for storage and transport. For optimal detection of mycobacteria, the sediment from centrifuged milk must be examined immediately (within 24-48 h of collection) or otherwise frozen.
• Decontamination, a process intended to selectively eliminate non-mycobacterial flora and enhance the isolation and detectability of mycobacteria by preventing other bacterial overgrowth, is another factor that influences the isolation of mycobacteria from specimens. The decontamination process may significantly decrease the number of viable mycobacteria, and up to 80-90% may die (Krasnow and Wayne 1969; Anon 1994). The type of decontaminant and the concentration at which it is used, are critical. Decontaminants include detergents such as 0.375-0.75% hexadecylpyridinium chloride (HPC), alkalis (2-4% sodium hydroxide), and acids (5% oxalic acid).
9.8.1 Species Identification and Typing
Culture on its own is a mechanism of detecting M. bovis in specimens collected from animals suspected to be suffering from BTB. Colony morphology does not allow accurate species identification, although the characteristic growth pattern and morphology of the colonies provide sufficient information to make a presumptive diagnosis of M.
bovis.A series of tests based on phenotypic characteristics and biochemical properties was traditionally used during the past decades to identify the various species of the MTC. The process is slow, cumbersome and inaccurate, and results are often ambiguous (Ramos et al. 2014; Ramos et al. 2015). These tests cannot be done in many laboratories, and they have largely been replaced by a number of sophisticated molecular genotyping techniques allowing linage and strain identification (Fig. 9.31). Identifying the individual strains is critical for implementing molecular epidemiological studies to determine the origin and spread of the disease to allow the design of more focused control measures and eradication campaigns (Collins 1999; Courcoul et al. 2014; Drewe and Smith 2014; Gormley et al. 2014).
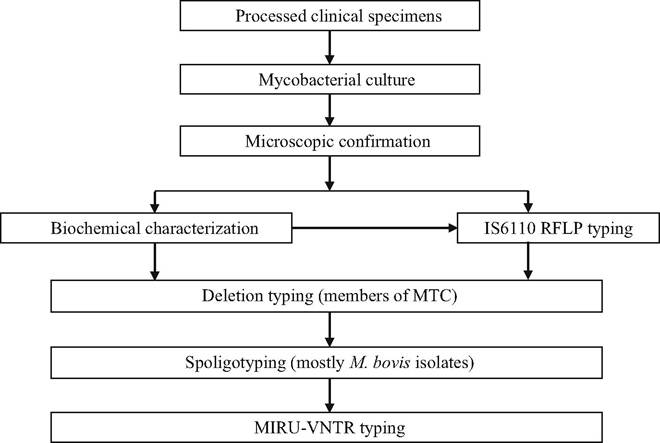
Fig. 9.31 Schematic flowchart showing mycobacterial identification techniques and common molecular diagnosis steps
Recent developments also include the use of different molecular techniques for typing and strain identification of M. bovis to characterize and untangle the molecular epidemiology of the disease to more adequately control its spread and to determine and control the origin of the infection. Few countries in Africa have the financial and human resources to implement the use of these tests. However, because of increasing collaboration with foreign laboratories, some data have become available. This information can be used to determine the source of infections and patterns of spread, enabling more focused approaches to controlling the infection (Cadmus et al. 2011; Ramadan et al. 2012). The details of these matters are addressed in Chap. 8 (molecular epidemiology) and in some of the country reports.
Despite the extensive genetic homology (99.9% genomic sequence homology and identical 16S rRNA sequences) of the members of the MTC, individual species, lineages, and strains can be differentiated by comparing differences in the sequences in the remaining 0.1% of their genomic structure.
These differences are the consequence of variations in single nucleotide polymorphism (SNP) and large sequence polymorphism (LSPs), also known as regions of difference (RDs) (Brosch et al. 2002; Garnier et al. 2003). These characteristics are the basis of the various molecular diagnostic techniques.During the past few decades, PCR-based assays, DNA mapping, and whole genome sequencing have been widely used for species and strain identification of mycobacteria and to elucidate genetic diversity, their evolution, pathogenicity, drug resistance, and the geospatial distribution of M. bovis in parts of Africa.
Much of the current research is focused on developing more sensitive and specific tests that are inexpensive and rapid. They should also be appropriate for use particularly in resource-poor countries, and should be suitable for use in the field in rural areas where there is a lack of infrastructure, electricity, and a cold chain.
Two categories of tests utilizing genetic markers exist: those that use whole genome techniques and those that are based on partial genome techniques. The partial genome techniques include restriction fragment length polymorphism (RFLP) analysis, spoligotyping, variable number of tandem repeat typing (VNTR), IS6110-ampliprinting (MPTR), random amplified polymorphic deoxyribonucleic acid analysis (RADP), RD typing (RD), and single nucleotide polymorphism (SNP) typing. Whole genome typing allows the use of the microarray technique for comparing particular strains to sequenced reference strains (Gormley et al. 2014).
The first step following culture is to distinguish the MTC mycobacteria from non-tuberculous mycobacteria, and this can only be done by the use of nucleic acid amplification methods such as PCR and spoligotyping. Both these tests are technically challenging, time-consuming, and expensive and, although highly discriminatory, are prone to homoplasy or convergent evolution, where the same genetic profile could be obtained in distinct MTC strains that are phylogenetically unrelated, thus confounding strain classification and phylogenetic inference (Hauer et al.
2016). Given the limitations of many of the current techniques, and the cost and technical ability required to use them, only some have been used in Africa, and it appears that there are specific problems in various countries limiting their use. The following typing tests are more often used in Africa:IS6110 Restriction Fragment Length Polymorphism (RFLP) This typing test has for long been used as the gold standard for differentiating M. tuberculosis strains (van Embden et al. 1993), but it has a limited value for typing M. bovis in which certain strains, such as those found in Europe, have one or only a few copies of IS6110 (Smith 2012). In Africa too about 90% of isolates in Madagascar (Razanamparany et al. 2006) and 83% in Cameroon (Njanpop-Lafourcade et al. 2001), possess only a single copy of IS6110, while 44% of isolates in South Africa contain only two copies (Michel 2008). The resolution, because of the low number of IS6110 copies in M. bovis strains, is too low for it to be of value. In addition, the test is time-consuming, labor-intensive, and technically demanding and requires large amounts of high-quality DNA (Deriemer and Daley 2004). Despite these limitations, the use of IS6110 RFLP typing remains essential in Africa as it is used to screen AFB-positive bacteria from cultures to differentiate between members of the MTC and NTMs or other related mycobacterial genera (Rigouts et al. 1996; Michel et al. 2008; Malama et al. 2014).
Spoligotyping (Spacer Oligonucleotide Typing) Spoligotyping involves PCR amplification of highly polymorphic repetitive units in a particular genomic region, known as the direct repeat (DR) locus, which are only present in members of the MTC. The DR locus belongs to the clustered, regularly interspaced, short palindromic repeat (CRISPR) family of repetitive DNA (Skuce and Neill 2001). The DR region in MTC could be associated with selective advantages and may be of specific biological importance to the pathogen (van Embden et al. 1993).
The DR locus includes a series of well-conserved, virtually identical, 36 bp DRs separated by unique, non-repetitive DNA spacer sequences of 34-41 bp length (the direct variant repeat (DVR) units). The spacers usually occur only once (occasionally twice) and are separated by one or several DRs and other spacers (Boer et al. 2004). The DR region may contain over 60 DVR units; however, only 43 of the spacer units (37 spacer sequences of M. tuberculosis H37Rv and 6 of M. bovis BCG P3) are used in standard spoligotyping of the MTC. The DR region is polymorphic because of the loss (deletion) of a single or multiple spacers, a condition that cannot be repaired by recombination, and, hence, is lost from a lineage forever (Smith et al. 2006) and thus remains a stable marker that does not change over time.
Spoligotyping detects the presence or absence of these 43 spacers of known sequence in the DR region by hybridization of PCR-amplified spacer DNA to a set of immobilized oligonucleotides, representing each of the unique spacer DNA sequences (Boer et al. 2004). The spacers are arranged on the blot in the order in which they appear in the H37Rv DR sequence (spacers 1-19, 22-32, and 37-43) and the sequence of M. bovis BCG P3 (spacers 20, 21, and 33-36). Polymorphism among spoligotypes of MTC is based on the presence or absence (loss of hybridization signal) of one or more spacers, compared to the complete 43-spacer set of M. tuberculosis H37Rv (Kamerbeek et al. 1997). The simplicity and robustness of this procedure and the ease and accuracy of presenting results in highly diverse portable numerical values make spoligotyping a widely used tool in clinical laboratories and for application in molecular epidemiology, evolutionary, and population genetic studies (Zumarraga et al. 1999).
Spoligotyping also has serious limitations. Its main disadvantage is that all genetic polymorphisms are restricted to a single genomic region, the DR locus, which limits its resolution. Furthermore, spoligotype patterns can be homoplasic, i.e., identical spoligotype patterns can be found in epidemiologically unrelated strains (Milian-Suazo et al. 2002), nullifying the assumption that strains with the same spoligotype patterns are identical by descent. However, homoplasic spoligotype patterns can be rare (Smith et al. 2006), and could be differentiated, when present, by variations in their VNTR patterns and other molecular markers. Therefore, from a practical point of view, spoligotype patterns are useful in the diagnosis of TB under African condition, since all MTC strains from humans and animals can be differentiated (Munyeme et al. 2009). This is important because it is becoming clear that animals that appear to be affected by M. bovis may actually be infected by M. tuberculosis or M. africanum (Sanou et al. 2014). The extent to which M. tuberculosis can occur in lymph nodes of cattle with BTB-like lesions is surprising; as in a study in the Eastern Cape, South Africa, 96.9% of isolates from 182 MTC-positive lymph nodes were strains of M. tuberculosis (Bhembe et al.
2017). This has implications for disease management (as MTC stains differ in their drug resistance profiles) and for establishing the source of infection. The advantage of combining spoligotyping with MIRU-VNTR (Weniger et al. 2010) and Hain®Genotype MTBC was demonstrated in Cameroon where it was determined that M. bovis strain Afrl was the dominant clonal complex with 97 unique genotypes. It also determined that a third of infected animals with multiple lesions had multi-strain infections. The data generated by the combination of these techniques allow characterizing stable and unstable endemic infections in different parts of the country, thus enabling the implementation of suitable control measures for the specific situation (Egbe et al. 2017).
Spoligotyping has several attributes that make it particularly suitable for resourcepoor African countries, and it is the most popular molecular technique in Africa for the following reasons:
• It can detect and type MTC directly in a range of clinical samples (Deriemer and Daley 2004; Kamerbeek et al. 1997),
• It is fast and relatively easy to perform and requires only a small amount of DNA (Deriemer and Daley 2004; Zumarraga et al. 1999).
• Spoligotyping patterns can easily be compared with results from other countries or regions by using the freely accessible international database (http://www. Mbovis.org) (Smith and Upton 2012). This database holds information and authoritative names for spoligotype patterns of all animal-adapted MTC strains (and M. africanum), in which RD9 is deleted.
Region-of-Difference (RD) Deletion Analysis RD variability is attributed to deletion or truncation of specific genomic elements due to replication slippage of DNA polymerase (Brosch et al. 2002), and has a nonrandom distribution, with a tendency to aggregate in a specific region (Rao et al. 2005). These may be missing genes that encode virulence factors such as the organism’s development of resistance to drugs and adaptation to environment and host, the presence or absence of which could give clues to clonal lineage of various mycobacterial isolates in specific geographic regions (Rao et al. 2005). Mycobacterium tuberculosis lacks 16 RDs, labeled as RD1 to RD16 that occur in other members of the MTC. Some are deleted from specific members of the MTC, and their absence allows distinguishing between the various species of the MTC (Parsons et al. 2002). Such comparative analysis of deleted distinct regions from the MTC genomic structure, provides a basis for differentiation and understanding the evolutionary lineage/relationship of members of the group. For example, Parsons et al. (2002) were able to accurately differentiate 88 well-characterized isolates of M. tuberculosis, M. africanum, M. microti, M. bovis, and M. bovis-BCG using PCR-based RD analysis of only six regions (RD1, RD3, RD5, RD9, RD10, and RD11).
In Africa, RD analysis is frequently used to differentiate M. bovis (which lacks RD4) from other members of the MTC and to differentiate M. tuberculosis from animal-derived strains of the MTC, including M. africanum. Additionally, RD analysis provides an understanding of the lineages of the current population of mycobacterial pathogens, and in addition to its diagnostic use, RD analysis could be a useful tool in epidemiological and evolutionary studies of mycobacterial species (Gordon et al. 1999; Brosch et al. 2002). Currently RD is considered to be the gold standard for genotyping mycobacteria in the MTC (Faksri et al. 2016).
Multilocus Variable-Number Tandem Repeat Analysis (MLVA) Typing MLVA is a PCR-based molecular typing technique that exploits natural variations in tandem repeat (TR) DNA sequences, also known as variable number tandem repeat (VNTR) loci, that are found in multiple loci in the genome of a variety of organisms (Ablordey et al. 2005). In MLVA, specific primers target the flanking sequences of these VNTR regions (Diguimbaye-Djaibe et al. 2006), the variations of which generate PCR amplicons of different fragment length; such length polymorphism forms the basis for the generation of diverse alleles. VNTRs used in mycobacterial studies are known as mycobacterial interspersed repetitive units (MIRUs), Queen’s University Belfast (QUB), exact tandem repeats (ETRs), or Mtub (M. tuberculosis) (Diguimbaye-Djaibe et al. 2006). Although there are subtle differences between the loci identified by these approaches, there is an overlap, and some of the chromosomal regions are known as ETR, MIRU, and/or VNTR loci (Smith et al. 2006). More than 40 of these loci have been investigated so far. Nearly half of them produced no discrimination, and the remaining loci gave varying degrees of discrimination among M. bovis isolates depending on the panel of VNTRs used (Collins 2011). When small numbers of VNTR loci are used, the level of discrimination achieved is not good. Larger panels and other sets of polymorphic repeats (MIRU-VNTR) involving 12 loci and 15 or 24 loci subsets, are frequently used, but this causes confusion, as the results of the different combinations cannot be compared. Of 29 loci combinations (QUB, ETR, MIRU, and Mtub) in South Africa, only 13 loci were stable and polymorphic, 14 were monomorphic, and 2 could not be amplified (Hlokwe et al. 2013), and of 26 MIRU-VNTR loci from 32 M. bovis strains in Burkina Faso, 9 loci were monomorphic (Sanou et al. 2014). In Chad, five loci were highly discriminative, two were moderately so, three were poor, and six loci showed no polymorphism at all (Hilty et al. 2005) (Table 9.3). The technique has also been used to type and determine the origin of M. bovis strains in Ethiopia (Biffa et al. 2014) and the molecular epidemiology of the disease in Nigeria (Jenkins et al. 2011). These reports emphasize the importance of evaluating the discriminatory power of different loci for each country or geographical region. It is thus important that the loci employed in individual countries should be those that will provide the biggest likelihood to discriminate the characteristic clonal complexes of its M. bovis population. The strain genetic variability in a country depends on the dominant existing clonal complexes, and the best results will be rendered by a combination of typing methods (Hauer et al. 2016).
Using the web-based server (http://www.miru-vntrplus.org/) makes it easier to determine the lineages of MTC strains. Furthermore, comparison between strains can also be made by comparing information maintained on the server including copy numbers of 24 MIRU loci and other molecular features. Additionally, the web-based server allows standardized nomenclature of different MIRU genotypes for ease of scientific communication (Hilty et al. 2005; Allix-Beguec et al. 2008). However, in order to adequately exploit what the server has to offer, African diagnosticians and researchers will have to align their selection of loci combinations for diagnostic purposes, as the combinations used in different countries in Africa are significantly different from the 24 loci set contained in the server.
Table 9.3 Molecular techniques to characterize M. bovis and other members of the MTC in Africa
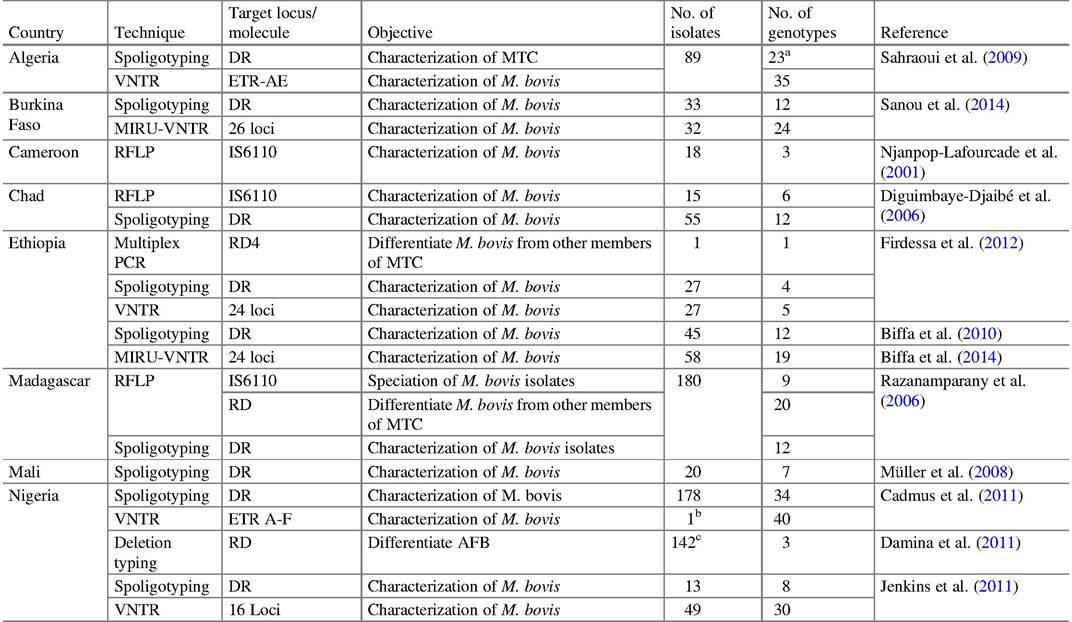

"M. caprae (SB 1451)
bSB0944
cM. bovis (107), M. tuberculosis (6), M. africanum (2), others (27) not typed
Lateral Flow Immunochromatographic Device The recently developed rapid lateral-flow immunochromatographic device (LFD) to confirm the identity of M. bovis is less sensitive than the qPCR methods. Its development is an attempt to create a rapid, inexpensive test. Currently the LFD results vary from inconsistent, as the test does not detect all spoligotypes, particularly because of M. caprae that also causes bovine tuberculosis and contain spoligotypes not detected by LFD, to almost 100% concurrence with other tests. The intention for the test is not to replace spoligotyping but to aid in the detection of, M. bovis-positive cultures that are not detected by Ziehl-Neelsen staining (Stewart et al. 2017).
9.8.2 Diagnosis of BTB in Wildlife
The presence of BTB in wildlife in the same ecosystem as that of cattle has become an international complication when attempting to eradicate BTB from cattle. There is increasing recognition of the role that different species of wildlife play in the epidemiology of the disease and in sustaining the infection by becoming maintenance hosts. Africa is richly endowed with a wide variety of wildlife species, most of which are probably susceptible to infection with members of the MTC. Some reference has been made to the diagnosis of the infection in wildlife in other sections of this chapter. Little attention, however, has until recently been given to the diagnostics of BTB in African wildlife, and authorities dealing with BTB face an enormous challenge to cope with the issues given the number of species and distribution of them throughout Africa. Against the background of the history of the role of wildlife in developed countries and elsewhere, dealing with wildlife, in the process of eradicating BTB will in all likelihood be a mammoth task.
Maas et al. (2013) recently comprehensively reviewed the problems associated primarily with the ante-mortal diagnosis of tuberculosis in wildlife. One of the main impediments of making a correct diagnosis in these species, is the lack of specific and sensitive diagnostic tests that have been validated and are specific and sensitive enough to satisfy the demands of the various reasons for testing them for the presence of the disease. These include surveillance, monitoring, diagnosis in individual animals, and certification of the BTB status for various purposes including those of import and export.
It has become a habit, and a bad one at that, to use the current ante- and postmortal tests, without validation, in a variety of wildlife species, and to rely on the results based on the extrapolation of cut-off points and responses in various other species. This practice cannot be condoned, although certain importing and exporting countries seem to accept these test results. Allowing this type of testing and accepting the results are high-risk practices, and it emphasizes the need to validate the tests, at least for those wildlife species that are traded in, and for vulnerable species facing extinction.
9.9