BOVINE VIRAL DIARRHOEA
KAI FROLICH
Tierpark Arche Warder e.V. Warder, Germany
Bovine viral diarrhoea (BVD) and mucosal disease (MD) are generalized viral infections caused by pestiviruses affecting a broad range of hosts, mainly artiodactyls.
Wildlife may be infected with more pestivirus species than have been identified at present.AETIOLOGY
Bovine viral diarrhoea virus (BVDV) is a positive-sense, single-stranded RNA virus 12.5 kb in size with a single open reading frame encoding four structural proteins (the capsid protein and the envelop glycoproteins Erns, E1 and E2) as well as six non-structural proteins. The virions are pleomorphic, spherical structures 50—60 nm in diameter, with a bilaminar envelope of cellular origin surrounding a semidense core of 20 to 25 mm diameter. Virions mature within intracytoplasmic membranes, and the virus is liberated by exocytosis of virus-containing membrane vesicles. Infectivity of BVDV is lost at elevated temperatures (e.g. mean half-life at 37°C and pH 7 is 7 hours) and by treatment with detergents and lipid solvents. The viruses are able to withstand a relatively broad pH range (pH 3—11).
Two antigenically distinct genotypes of BVDV exist (BVDV1 and 2) with further subdivisions discernable by genetic analysis. Isolates within these groups exhibit considerable biological and antigenic diversity (Figure 10.4). BVDV1 is clustered into as many as 11 distinct genotypes. Two main genotypes (2a and 2b) have been described for BVDV2(17,18). BVDV2 represents around 50% of the BVDV isolates in North America. In Europe, more than 90% of the BVDV isolates are BVDV1. BVDV1 was isolated in 1954 and BVDV2 was first described in the early 1990s in North America as an emergent highly pathogenic viral genotype. Besides North America, BVDV2 has also been reported in Germany, Belgium, France, the Netherlands, Austria, Slovakia, Italy and the UK but not in Sweden, Norway, Spain, Slovenia or Denmark(19,20).
EPIDEMIOLOGY
GEOGRAPHICAL DISTRIBUTION AND HOSTS
Serologic surveys and virus isolation have provided evidence of infection with BVDV or related pestiviruses in more than 40 species of free-ranging wildlife worldwide. However, the extent of disease caused by BVDV in wild species is more difficult to assess, as many reports lack viral isolation and the clinical signs of various BVD viruses are
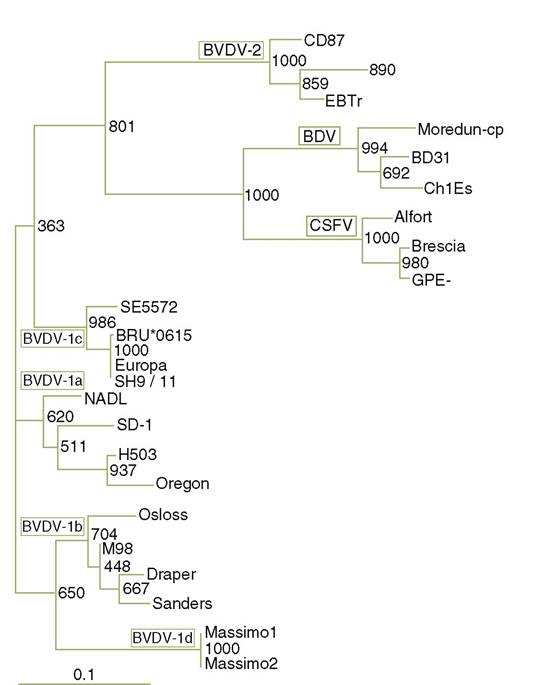
FIGURE 10.4 Phylogenetic tree based on 5' — UTR sequence comparison from different pestivirus (mainly BVDV) strains; modified and used with permission. (See Giangaspero et al., 2001)(16). Giangaspero et al. (2001)(16).
insufficiently specific. The distribution of BVDV infections in wildlife from Europe is shown in Table 10.1.
I n addition to serological evidence of BVDV in wild species in Europe, several characterizations of isolates have been conducted. Isolates from two seronegative free - ranging roe deer (Capreolous capreolus) from Northern Germany were analysed(33,34) and found to belong to the BVDV strain called SH 9/11. If the pathogenesis in deer is similar to that in cattle, the two isolates from roe deer probably were taken from PI individuals or those who had an acute transient form of BVD. The SH 9/11 strain is highly homologous (up to 93% identity) to ‘classical’ BVDV strains. However, SH 9/11 has characteristic variations in its 5 ' UTR distinct from all other BVDV- strains and it is distinct from the other cattle isolates tested. Other isolates from roe deer were found in Hungary(35) and in Southern Germany(36), but phylogenetic analyses were not performed in these cases.
TABLE 10.1 Evidence of BVD infections in wildlife from Europe.
| Species | Country | Antibody (AB), virus isolation (VI), lesions (L) | References |
| Cervidae | |||
| Red deer (Cervus elaphus) | Denmark, Norway, Italy, UK, France, Germany, Austria | AB, VI | 2, 21-32 |
| Roe deer | France, Norway, | AB, VI, L | 2, 23, 25, 26, |
| (Capreolus capreolus) | Germany, Hungary, Sweden, Spain | 28, 30, 33-40 | |
| Fallow deer (Dama dama) | UK, Italy, Germany | AB, VI, L | 2, 22, 27, 41-46 |
| Sika deer (Cervus nippon) | UK | AB | 22 |
| Reindeer (Rangifer tarandus) | Norway | AB, L | 30, 47, 48 |
| Moose (Alces alces) | Sweden, Norway | AB, VI, L | 30, 37, 49 |
| Chinese water deer (Hydropotes inermis) Bovidae | UK | AB | 22 |
| Chamois | France, Italy, | AB, VI | 25, 28, 32, 40, |
| (Rupicapra rupicapra) | Germany | 50 | |
| Ibex (Capra ibex) | France | AB | 25 |
| European bison (Bison bonasus) Suidae | Poland | AB | 51, 52 |
| Wild boar (Sus scrofa) Lepridae | Italy, Germany | AB | 28, 32, 45, 53, 54 |
| Rabbit (Oryctolagus cuniculu() | Germany | AB, VI | 55a, 56a, 57 |
aExperimentally infected individuals
Moreover, BVDV was isolated from captive fallow deer (Dama dama) from Germany and Sweden and from captive and free-living red deer ( Cervus elaphus) from UK and France.
Pestivirus was isolated and confirmed by genetic methods in captive reindeer (Rangifer tarandus) and European bison (Bison bonasus) (Germany)1-58). However, in the light of new results the reindeer isolate was typed as Border disease virus 2(4). BVDV was also isolated from farmed alpacas (Lama pacos) in the UK. Phylogenetic analysis showed that this strain was of subgenotype 1b(59).The principal source of BVDV within a population is PI individuals, which, by virtue of a high titre of virus shed in their secretions and excretions, infect healthy individuals. These PI carriers are especially important as virus excreters in maintaining the infection, because the virus is too unstable to survive long in the environment^0). PI animals typically constitute 1—2% of the population1-61). Transmission of BVDV by acutely infected animals is not as efficient because the virus is only excreted for a relatively short time (2—15 days). The virus may also be present in aborted fetuses, fetal membranes and uterocervical fluids and semen. Transmission by mechanical and insect vectors (Stomoxys calcitrans or Haematopota pluvialis) has been reported(62,63). In livestock in Europe BVDV has been endemic in all countries where no systematic control was applied. Approximately 50% of all cattle herds in Europe have PI animals, and 90% of all cattle become exposed during their lifetime(64).
The role of pestiviruses in wild ruminant populations and the interactions between wild ungulates and domestic livestock are not well understood. Suspected sources of the virus from wild animals include direct contact with infected livestock, shared feed and watering areas, or the presence of pestivirus-infected individuals within wildlife populations. Several investigators have speculated that an independent cycle occurs among wild ruminants in the absence of domestic animals. It has been hypothesized that distinct BVDV strains circulate in the free- ranging roe deer populations1-33’34).
This hypothesis is supported by the results of serological surveys; on the one hand, no significant differences in antibody prevalence among deer in habitats with high, intermediate and very low density of cattle were found, suggesting that free- ranging deer can become infected with BVDV without having contact with cattle(2). On the other hand, a causal relationship between the spread of BVDV in cattle and its occurrence in deer has been suggested, and it has been speculated that wild ruminants may serve as a reservoir of BVDV for cattle(62). At present there is no evidence that BVDV spreads from free-living animals to domestic ruminants. However, theoretically it cannot be excluded that transmission of a highly virulent pestivirus strain from domestic to wild animals or vice versa could occur and have a profound effect on both eradication programmes and wildlife populations1-1).PATHOGENESIS, PATHOLOGY AND IMMUNITY
Bovine fetuses infected with ncp BVDV during the first trimester of gestation may become PI with the virus. These fetuses are born persistently infected with the BVD virus and remain in this state for the rest of their lives until death; death is usually caused by the MD manifestation of BVDV infection. The PI fetuses usually develop as unthrifty calves that may be otherwise normal. PI bovines infected with ncp BVDV may succumb to fatal MD when superinfected with a cp strain. MD usually occurs in calves and young cattle between 6 and 24 months of age but also occasionally in older animals.
Two patterns of disease development can be observed. In one, cattle become moribund within 2—3 weeks after superinfection, developing an early-onset of MD. In the second pattern, animals recover after a transient phase of mild disease, but they may develop a late-onset MD after a time period varying from 4 weeks to several years. Both patterns of MD terminate in severe diarrhoea, erosions of the mucocutaneous membranes, anorexia, depression and ocular and nasal discharge, followed by generalized deterioration of the affected animal.
Erosive and ulcerative lesions in the alimentary tract and depletion of lymphoid tissues are regarded as being characteristic for MD. Animals eventually show skin lesions and lameness due to interdigital ulceration and inflammation of the coronary bands.Erosive to ulcerative lesions of oral mucosa and other mucocutaneous membranes of the digestive tract and catarrhal to fibrinous-diphteroid enteritis accompanied by general physical impairment are typically found in the MD form of BVDV infection in cattle.
Comparison of the course of BVDV infection in livestock to that in wild ruminant species is difficult, and at present there is insufficient data to make comparisons between the complex pathogenesis of BVDV in cattle and pathogenesis of BVDV infection in any wild species. Transplacental infections have not been reported in wild animals; however, mummified fetuses and stillborn and normal healthy fawns were observed in white-tailed deer (Odocoileus virginianus) experimentally inoculated with a BVDV originally isolated from a white-tailed deer in North America1-65). Inoculation of red deer, white- tailed deer and mule deer ( Odocoileus hemionus) with ncp BVDV resulted in subclinical infections1-21’66). In reindeer, infection with cp BVDV resulted in transient mild diarrhoea, coronitis and laminitis(67). The transmission of BVDVlf from a PI lesser Malayan mousedeer (Tragulus jaυanicus) to two bovine calves has been demonstrated1-68). In another study a lesser Malayan mousedeer, persistently infected with ncp BVDV type 1f, was experimentally superinfected with a cp BVDV type 1c, which antigenically partially matched the endogenous strain. Following superinfection, the lesser Malayan mousedeer did not develop clinical signs or lesions consistent with MD(69). This may indicate a different course of BVDV infection in the lesser Malayan mousedeer than in domestic ruminant species.
Development of antibodies to BVDV has been reported in reindeer(67), and in mule deer and white-tailed deer(66). In naive animals, antibodies are detected 8—15 days after infection; however, no systematic research has been performed regarding titre development in wild species after experimental infection. Therefore, it can be only assumed that, like in cattle(70), titres persist for life in wild ungulates.
CLINICAL SIGNS
The reason for the worldwide distribution of BVDV lies in its ability to cause two types of infection: i) transient (or ‘acute’ ) infections in the majority of cases; and ii) persistent infection.
The usual form of a BVDV infection in livestock (and probably in wildlife) is subclinical, but acute disease of high morbidity and low mortality or chronic disease of low morbidity and high mortality may occur. Initial clinical signs include pyrexia, anorexia, nasal discharge and lacrimation, leucopenia or general physical impairment followed by diarrhoea of varying intensity and duration.
The primary visible clinical signs in wild ungulates are haemorrhagic mucosal inflammation and general physical impairment (e.g. weight loss, apathy, immunosuppression). In several cases animals have erosive, ulcerative and nerotizing lesions of the digestive mucosa and catarrhal to haemorrhagic enteritis.
Some BVDV isolates in livestock may cause the so- called haemorrhagic syndrome, characterized by petechial haemorrhages in skin and mucous membranes, due to severe thrombocytopenia.
Transplacental infection with BVDV may, depending on the time ofgestation, result in the development ofa PI fetus, abortion, stillbirth, fetal malformation (cerebellar hypoplasia, microphthalmia, mandibular brachygnathism) or weak neonates. Early fetal losses are manifested as infertility and prolonged intervals between successive calvings as a result of these early embryonic deaths. Abortions can occur at any stage of gestation, depending on when the dam is infected. PI animals may appear either completely healthy or show abnormal signs such as growth retardation.
DIAGNOSIS
Various methods are available for diagnosing acute or persistent BVDV infections: immunohistochemistry (IHC), antigen and antibody ELISA, RT- PCR assays, and virus isolation. For detection of seropositive reactors by ELISA in wildlife, competitive ELISA is recommended, because specific anti-species antibodies are usually lacking. However, VNT is the ‘gold standard’ for the detection of specific antibodies in wild animals.
Acute infection with BVDV may be demonstrated by showing seroconversion using sequential paired serum samples (at an interval of at least 2 weeks) from several animals in the group.
Buffy coat cells, whole blood, washed leucocytes or serum are suitable for isolation of the virus from acutely infected or suspected PI live animals. At necropsy, samples of thymus, spleen, lung, liver, mesenteric lymph nodes, tonsils, intestine and kidneys should be collected for virus culture or RT-PCR. Viral antigen can be detected in acetone-fixed frozen tissue sections with immunofluores- cent antibody techniques or in formalin-fixed paraffin- embedded tissues by IHC analysis. Confirmation of MD is possible by histopathology to demonstrate typical lesions coupled with IHC to show association of viral antigen with lesions, and if necessary isolation of the cp biotype as well as the ncp biotype.
PUBLIC HEALTH CONCERNS
Specific anti-BVDV antibodies have been found in up to 87% of animal handlers and veterinarians1-71). Among children under 2 years, pestivirus antigens were present in 24% of specimens from diarrhoea episodes that could not be explained by more common enteric pathogens1-72). Currently, genotypes of pestivirus RNA were detected in live virus vaccines for human use (e.g. mumps, measles)1-16).
MANAGEMENT, CONTROL AND REGULATIONS
Control of BVDV infection of cattle is generally achieved by vaccination with attenuated live, modified live or killed BVDV vaccines, and by the identification and removal of persistently infected carriers of the virus.
Live vaccines contain cp BVDV. This vaccine is unable to produce PI animals. However, fetal infections after vaccination have been observed. Most probably they were attributable to non-cp contaminants of the vaccine. Furthermore, if PI animals are vaccinated with cp BVDV strains, MD may be triggered due to genetic recombination between the vaccine and the persisting virus. Killed vaccines are safer to use than live vaccines and are mostly used in wild animal species. However, they tend to be substantially less effective than live vaccines and require frequent boosters (twice a year). Various inactivated vaccines contain both cp and ncp strains of the virus.
In livestock, large-scale control efforts have been implemented in France, the Netherlands, Germany and Italy. Time-limited, project-type control efforts have also been implemented in Greece and Spain. In the UK, Ireland, the Netherlands and Slovenia, only killed BVDV vaccines are licensed. The Scandinavian countries and Austria do not permit the use of BVDV vaccines; instead, large-scale eradication schemes are in place. To build upon the growing interest in BVDV control a Thematic Network on BVDV Control, funded by the European Union Commission’s 5th Framework Programme was formed in 2002 (www.bvdv-control.org)(73).
Wild ruminants share range (pasture), feed and water sources with cattle; therefore development of adequate control programmes is dependent on knowing whether there are reservoirs of BVDV on the ranges other than domestic cattle(74). It is assumed that wildlife reservoirs can seriously undermine the effectiveness of any domestic BVDV control strategy by acting as potential sources of virus that can infect cattle, especially at the wildlife— livestock interface(75). Also, it has been suggested that the reintroduction of BVDV to negative cattle herds in the UK can be due to contact with a wildlife reservoir1-76). However, the natural mode of transmission of BVDV to wild ungulates, and the question of whether wildlife could serve as a reservoir, is not yet clear.
SIGNIFICANCE AND IMPLICATIONS FOR ANIMAL HEALTH
BVDV is a major pathogen of cattle worldwide, with significant economic impact on livestock production. The impact of BVDV-associated diseases on the health of free- ranging wild ruminant populations is currently unknown; there are few confirmed cases of pestivirus-caused disease in these species but no evidence that these viruses have significant population impacts(65).