PESTIVIRUS OF CHAMOIS AND BORDER DISEASE
IGNASI MARCO
Servei d' Ecopatologia de Fauna Salvatge (SEFaS) — Wildlife Diseases Research Group, Departament de Medicina i Ciru- rgia Animals, Facultat de VeterinHria. Bellaterra, Spain
Border disease virus (BDV) is responsible for border disease (BD), first reported in sheep in the border region between Wales and England in 1959 and now distributed worldwide.
It is a congenital disease characterized by barren ewes, abortions, malformations, stillbirths, birth of small weak lambs and persistent infections of the offspring.Aetiology
BDV is a member of the genus Pestivirus that mainly infects sheep, but also goats and occasionally cattle and pigs. Among pestiviruses, BDV has the greatest genetic diversity and six major genotypes have been reported, although new subgroups are being proposed.
EPIDEMIOLOGY
Evidence for BDV infection in wildlife is scarce. Antibodies against BDV have been demonstrated in roe deer (Capreolus capreolus), mouflon ( Ovis ammon) and Alpine chamois (Rupicapra rupicapra)(2’3). Isolation and characterization of BDV has been described in a reindeer (Rang- ifer tarandus) and a European bison (Bison bonasus) from a German zoo (BDV2 genotype)(4). In free-ranging wildlife, there were no reports of infection with BDV until its detection in an outbreak of a previously undescribed disease in Pyrenean chamois (R. pyrenaica) in 2001(5). Chamois BDV was characterized as BDV4 genotype, which is the same genotype as the BDV circulating in sheep in Spain(6).
The geographical distribution of the disease associated with BDV infection in Pyrenean chamois comprises an extensive area of the Central and Eastern Pyrenees in Spain, Andorra and France. Although the disease has been reported throughout this area, the prevalence varied in the different mountain massifs and valleys, with severe outbreaks reported (Figure 10.2).
The Pyrenean chamois has been the only species affected by the disease, even though it shares the habitat with other wild and domestic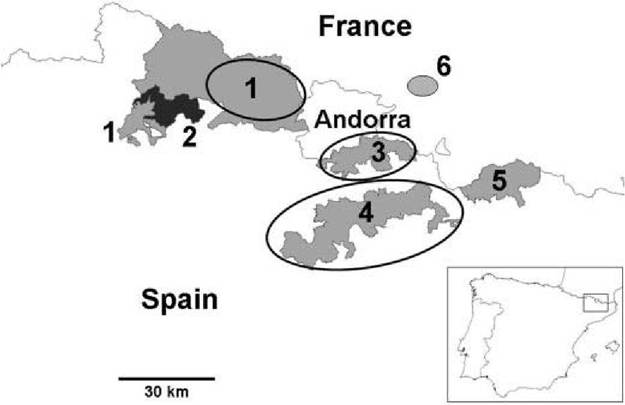
FIGURE 10.2 Map of the Central and Eastern Pyrenees showing the extent of the three main outbreaks of disease (circled areas) and the main protected areas: 1. Alt Pallars-Aran National Hunting Reserve; 2. Aiguestortes i Estany de Sant Maurici National Park (in black); 3. Cerdanya-Alt Urgell National Hunting Reserve; 4. Cadi National Hunting Reserve; 5. Freser-Setcases National Hunting Reserve; 6. Orlu National Game and Wildlife Reserve.
ungulates. No abnormal mortality has been recorded in any other species in areas where chamois were affected and serological and virological investigations indicated that chamois BDV infection among other species was unlikely(7). Nevertheless, BD is widespread in sheep in Spain, although little is known about its prevalence and clinical significance. BDV strains from chamois and sheep may co-circulate in the Pyrenees, and it is possible that they are a source of infection for other wild and domestic species, as ruminant pestiviruses are not strictly host-species-specific.
The main outbreaks of disease in Pyrenean chamois have affected important protected areas: Alt Pallars-Aran, Cerdanya-Alt Urgell and Cadi National Hunting Reserves (NHR). In addition, isolated cases of diseased chamois have been found throughout the area, including Aigu estortes i Estany de Sant Maurici National Park (Figure 10.2). The high, and in some cases, extreme mortality that occurred during these outbreaks, leading to the death of thousands of chamois, has no precedent in pesti- virus infections in wild ruminants. Mortality was difficult to assess accurately because most of the diseased chamois would not have been found due to the large size and remoteness of the affected areas. However, the annual censuses provided an indirect measure of mortality by showing a decrease in the population.
In Alt Pallars-Aran NHR, the estimated population decrease was 31% for the whole reserve and 42% in the main affected area. In Cerdanya- Alt Urgell and Cadl NHR, the estimated decrease was 86% and 63%, respectively1-8’9). It has been estimated that for the whole Central and Eastern Pyrenees from Northeastern Spain, chamois population decreased from approximately 13,000 animals in 2000 to about 9,000 in 2006, a 30% reduction.The epidemiology of the disease has followed different patterns in each of the different outbreaks. Research is being conducted to determine associated factors, such as pathogen, host and host population characteristics and ecological factors. Lack of immunity at the population level and the presence of secondary infections may have been important in determining the outcome of BD. Secondary infections have supposedly been magnified by the immunosuppressive effects of BDV infection and are most likely to be responsible for the mortality. The first outbreak of disease at Alt Pallars-Aran NHR showed a similar seasonal pattern for the 2 years it lasted (2001 and 2002): most of the affected chamois were found between late winter (February) and spring (June). Piroplasmosis was the most common secondary infection and severe lesions consistent with haemolytic anaemia were observed in spleen and kidney(8). The second outbreak at Cerdanya-Alt Urgell NHR in 2005 produced the highest mortality. It lasted almost 2 months, and bacterial pneumonia was the major contributing factor to the deaths. The third outbreak at Cadi NHR lasted for about 31 months and spread progressively to cover an extensive area of chamois habitat. Bacterial pneumonia was also the most common pathological finding in this outbreak1-9). Pasteurella spp. and Mannheimia haemolytica were frequently isolated from the lung lesions. The role of other concurrent infections that can influence the development of the disease has only partially been studied.
The absence of significant mortality in some parts of the Central and Eastern Pyrenees remains poorly understood. The chamois population is relatively continuous and abundant over the whole mountain chain. However, some areas with high density of chamois, such as Aiguestortes i Estany de Sant Maurici National Park, which is only 7 km away from a severely affected area, have not been affected. It was not until March 2009 that the virus was detected and isolated from the first reported clinical case. Factors such as natural movement of chamois, climatology and possibly other unknown factors, may have influenced this geographical distribution. Freser- Setcases NHR, in the Eastern Pyrenees, has probably the highest density of Pyrenean chamois from all the study areas (Figure 10.2). To date, no abnormal or mass mortality has been detected, despite this population being connected to the population of Cadi NHR. However, BDV similar to the previous chamois isolates were identified and isolated in 2006 from a healthy chamois and in 2007 from an isolated case of disease1-9). Therefore, the virus has reached or at least is present in this reserve, but to date it has not caused significant mortality. Prevalence of antibodies against pestivirus in this area was high (71% in 2003), thus, the immunological status at the population level may have been an important factor for the absence of the disease in this area. Nevertheless, the seroprevalence has been decreasing gradually since then, and was 38.5% in 2008. If this trend continues in the next few years, it could lead to an epidemic, if the virus is still circulating and if the population is losing antibody protection.
I n a retrospective study performed to detect BDV in archived frozen spleen from chamois from the Central and Eastern Pyrenees between 1990 and 2000, two pestiviruses were identified by reverse transcription polymerase chain reaction (RT-PCR) and isolated from one healthy chamois and one diseased chamois with pneumonia, both sampled in 1996 in Freser-Setcases NHR.
Sequence analysis revealed that the two viruses were grouped into the BDV4 genotype, closely related to the other BDV chamois isolates, indicating that BDV strains from chamois origin may have been present in the chamois population several years before the first cases of the disease in 2001(10). Therefore, among host and ecological factors, virus mutation and transformation into a pathogenic strain should be strongly considered.After the first severe die- off in 2001 and 2002 in Alt Pallars-Aran NHR, a high seroprevalence (71.9%) was observed in the chamois population. However, it decreased significantly after 2004, and since then frequent cases of BDV infection have been detected, suggesting that the infection became endemic, but could still be responsible for the continued slight decline in the chamois population over recent years(6). In the French Pyrenees, at the Orlu National Game and Wildlife Reserve, periodic oscillations of the chamois population have not been explained, because no diseased chamois have been observed and mass mortality has never been reported. However, in a retrospective epidemiological study, the high prevalence of viraemic animals suggested that BDV infection in chamois may have had a clinical effect on reproduction and on juvenile mortality, which may explain these variations in the population size(11).
There are few data on host and environmental factors associated with BDV infection in Pyrenean chamois. No sex or age predisposition was observed in two of the main outbreaks reported. However, during the first outbreak, which had lower mortality, adult males were predominantly affected(8). In the French Pyrenees, at the Orlu National Game and Wildlife Reserve (Figure 10.2), the same BDV genotype as in Spain was detected in chamois; however, no mass mortality or clinical signs have been observed. In this area, higher seroprevalence and low prevalence of infection have been reported in adult chamois than in individuals aged 2 years or less.
This concurs with the dynamic of the infection in domestic ruminants, in which young animals are most susceptible to acute infections(11).In sheep, the key for the transmission of BD is persistently infected (PI) animals that are infected during gestation and then become immunotolerant after birth, shedding high titres of virus throughout their lives. Thus, BDV transmission occurs mainly by direct contact by the oronasal route between PI animals and susceptible animals. Infection in non- pregnant animals is usually subclinical and leads to acutely infected animals, which are considered to be inefficient virus transmitters. However, by contrast, in naturally and experimentally infected adult Pyrenean chamois, detection by RT-PCR and virus isolation of BDV from oral, nasal and rectal swabs, urine samples and sera indicated a high level of virus excretion and viraemia(12) (Cabezon, unpublished data). Thus, acute infection in chamois leads to a high viraemia and efficient virus elimination that would explain the high horizontal transmission rate of the virus in the severe epidemics reported.
The existence of PI animals in Pyrenean chamois has not been confirmed. In domestic ruminants, confirmation of PI status requires identification of virus in two separate samples with a minimum of 21 days. More than 100 free- ranging diseased chamois have been studied since 2001, and almost all were viraemic and antibody-negative, using an enzyme-linked immunosorbent assay (ELISA) test. However, they could not be re- tested to check their PI status because most of them died shortly after capture. The epidemiological pattern of the three mentioned outbreaks, with rapid development and very high mortality, indicate that it is unlikely that the diseased chamois were PI. The diseased chamois included adult and old animals, which is uncommon in BD of sheep, because PI animals have poor performance and survival. Another aspect is that the presence of PI animals in domestic ruminants is associated with moderate to high flock seroprevalence. Previous data from chamois populations before the outbreaks is only available from the Cadi NHR, where in 2005 a low antibody prevalence (5%) was reported, suggesting that it was unlikely that there were PI animals in the population1-9). A diseased BD Pyrenean chamois survived in captivity for more than 1 month, and blood samples were taken five times during this period. Serology using a blocking ELISA test and virological studies by RT-PCR to detect pestiviral RNA in this animal showed no antibodies against pestivi- rus but the presence of BDV at all five time points. Accordingly, this chamois can be considered a PI animal(5). However, in an experimental infection of adult chamois, virus shedding was confirmed for more than 1 month until the animals were euthanized and antibodies were not detected using the same ELISA test, but high titres were detected with a virus neutralization test (VNT), using the homologous strain (Cabezon, unpublished data). Thus, BDV infection in non-pregnant Pyrenean chamois induces a long-lasting infection and an antibody response. In naturally infected chamois, the lack of antibody response should be further confirmed with a VNT using homologous strains.
I n healthy seropositive Pyrenean chamois, high cross- r eactivity was observed among different reference ovine BDV and Bovine viral diarrhea virus (BVDV) strains, which is frequent in pestivirus infection in domestic rumi- nants(6), and even the different BDV chamois isolates show different cross-reactivity. Therefore, cross-neutralization studies should be performed because antigenic differences may exist, not only between sheep and chamois BDV strains, but also between chamois isolates from the different locations of affected areas in the Pyrenees.
Vertical transmission was demonstrated in the fetus of a naturally infected female chamois that died before par- turition(12). In another study, three pregnant female Pyrenean chamois were experimentally infected with a BDV chamois strain. All animals became viraemic, one female aborted and all three females died before the end of gestation (Martin, unpublished data). Therefore, the existence of PI in chamois may be possible, but owing to the severity of the disease in this species, females may abort or die before giving birth.
PATHOGENESIS, PATHOLOGY
AND IMMUNITY
The extreme severity and mortality of the disease associated with BDV in Pyrenean chamois is unprecedented in pestivirus infections in wild ruminants. The pathogenesis of the disease in chamois is different from BD in sheep. While healthy young and adult sheep exposed to BDV experience only mild or inapparent disease, chamois infected with BDV develop severe clinical signs and lesions. Embryonic and fetal death, abortion and the birth of dead or weak animals with poor growth, which are characteristic of BD in sheep, are very difficult to study in free-ranging animals, like chamois, because aborted fetuses, birth products and weak neonates and calves are quickly removed by predators and scavengers in the wild and are not readily found by disease investigators.
Experimental infection studies using a chamois BDV isolate have been performed in pig, sheep and Pyrenean chamois. Infected pigs and sheep developed a short viraemia (almost undetected in sheep) and an important antibody response and virus clearance, without presenting clinical signs, macroscopic or histological lesions. However, five Pyrenean chamois presented a persistent viraemia from day 2 post- infection through all the experimental period (34 days), with seroconversion starting at day 11 post-infection (Cabezon, unpublished data). In all experimentally infected chamois, the homologous BDV4 was isolated from sera, main organs and nasal, oral, rectal and urine swabs. Although no evident clinical signs were detected, significant pathological lesions were observed in the brain, similar to those observed from field cases of infected chamois. Therefore, these results suggest that this BDV is the primary aetiological agent of the disease.
At necropsy, the most consistent finding in naturally infected chamois is emaciation with serous atrophy of fat. Enlargement of lymph nodes, especially the superficial cervical ones, is frequently observed. Bacterial bronchopneumonia, ranging from small pneumonic foci to large areas of lung consolidation, is the most common internal lesion. Other lesions are associated with concurrent infections and include abscesses, verminous pneumonia and orchitis. The most severe microscopic lesions are found in the brain. They consist of oedema, diffuse moderate spongiosis and moderate to evident diffuse gliosis with astrocyte hypertrophy. Occasionally, glial nodules are detected. Neuronal degeneration and necrosis occur throughout the brain, with the hippocampus and the Purkinje cell layer in the cerebellar cortex most severely affected(8). Occasionally discreet inflammatory infiltrates of mononuclear cells surrounding blood vessels have been observed. In lambs with BD, lesions in the central nervous system are different, as they are infected during gestation. Mainly, they consist of a deficiency of myelin and an increased density of interfascicular glia, which can persist for years.
In the skin, the alopecic areas show follicular atrophy and telogenization of the remaining hair follicles. In the epidermis, hyperplasia and melanosis with evident orthok- eratotic hyperkeratosis is observed, and in the dermis, mild mononuclear interstitial inflammatory infiltrates can be seen occasionally. These lesions are different from those described for BD in lambs, which consist of abnormal and straight birthcoat in fine or medium fleeced breeds, with an increased size of primary wool follicles and decreased number of secondary follicles.
The immune response to infection with BDV in Pyrenean chamois is not well understood. The epidemiological data indicate that populations with moderate to high seroprevalence of antibodies may be protected against disease, but affected populations and diseased animals do not have such antibody protection. However, in experimentally infected animals, despite the production of neutralizing antibodies, virus clearance was not attained.
CLINICAL SIGNS
Clinical signs most commonly observed in naturally infected Pyrenean chamois are non- specific, and include emaciation, depression, weakness and difficulties in locomotion. Skin lesions are frequent and consist of different degrees of alopecia and skin hyperpigmentation. The alopecia ranges from small asymmetric areas on the head (mainly at the base of ears and periorbital regions), neck and trunk, to extensive areas, with hair persistence only on ears, distal parts of the limbs and tail (Figures 10.3). Affected animals frequently exhibit abnormal behaviour, with absence of flight reaction and tameness, allowing their easy capture. These alterations may be related to the microscopic brain lesions observed. Dyspnoea is a frequent clinical sign, always associated with secondary bacterial pneumonia.
Pale mucous membranes are associated with anaemia caused by cachexia and inflammation, but also by intense tick infestation and presence of piroplasms (Theileria sp. OT3) in some cases(13’14). Other clinical signs occasionally observed in chamois with BD are diarrhoea and keratitis, both probably associated with secondary infections.
Haematological and serum biochemical studies in naturally infected chamois showed reduced red blood cell count, haemoglobin concentration, packed cell volume, mean corpuscular volume, lymphocyte count, glucose,
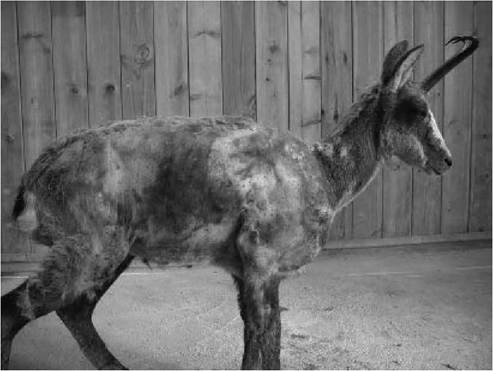
FIGURE 10.3 Extensive alopecia, emaciation and skin hyperpigmentation of a Pyrenean chamois (Rupicapra pyrenaica) with BDV infection.
lactate, triglycerides, creatinine, total protein concentrations and alkaline phosphatise. They also demonstrated higher neutrophil and platelet counts, total bilirubin, urea and aspartate aminotransferase. Most of these changes are non-specific and may be associated with the cachexia and inflammation in the diseased chamois, whereas the lymphopenia may be directly related to the BDV infection(14).
DIAGNOSIS
The diagnosis of BDV infection in Pyrenean chamois presents little difficulty in the field if animals are observed with emaciation, alopecia, depression, dyspnoea and abnormal behaviour. Nevertheless, laboratory confirmation is necessary, as other diseases described in chamois, such as sarcoptic mange, dermatophytosis, pneumonia, toxoplasmosis and cerebral coenurosis, should be considered in the differential diagnosis. RT-PCR and virus isolation have been used to detect pestivirus in chamois, as described for domestic ruminants(15). Antigenic characterization of isolates of BDV has been performed by the immunoperoxidase monolayer assay (IPMA) test using monoclonal antibodies C-16 (pestivirus-specific), CA-3 and CT-6 (BVDV1-specific), WV433 (BVDV2 specific) and WS363 (BDV-specific)(8). Viral antigen can be also demonstrated in most of the tissues of PI domestic animals by immunohistochemistry. Preliminary studies in chamois have also been performed on formalin-fixed, paraffin- embedded spleen and kidney samples, using the monoclonal anti-pestivirus antibody 15C5 directed against the 48 kDa glycoprotein of BVDV with positive results(5).
For population screening, there are several commercial ELISA antibody and antigen diagnostic tests for BDV, although they are not validated for wild ruminants. These have been used in several investigations in Pyrenean and Alpine chamois, and in some cases the results were confirmed by VNT. Six pestivirus strains have been used for these studies: namely BVDV1 strain NADL, BVDV2 strain atypical, BDV strain Spain 97, BDV strain Moredun, BDV strain 137/4 and BDV strain chamois, isolated from a diseased chamois in 2002. In healthy seropositive chamois, higher titres for sheep BDV strains than for the chamois strain have frequently been found. This could indicate an additional exposure in Pyrenean chamois to another BDV strain, probably of ovine origin, apparently without pathological consequences(6).
MANAGEMENT, CONTROL AND REGULATIONS
Control and eradication measures for pestivirus infection in domestic ruminants are mainly based on detection and removal of PI animals and vaccination. As it is not really known if PI animals are important for the spread and maintenance of the disease in Pyrenean chamois, and as these practices are not feasible in free-ranging populations of wild ruminants in remote mountain areas, it is unlikely that the infection can be controlled in the near future.
Control programmes for BD in sheep are not currently in place in the EU. BD is not included in the World Organisation for Animal Health (OIE) list and there is no designated reference laboratory.
PUBLIC HEALTH CONCERNS
There are no public health concerns, as humans are not susceptible to BDV infection.
SIGNIFICANCE AND IMPLICATIONS IN ANIMAL HEALTH
BDV infection in sheep has a worldwide distribution and causes economic losses through the impact on reproduction and health. In wildlife, no other information on mortality in species other than Pyrenean chamois has been reported.
ACKNOWLEDGEMENT
Research was supported by Direccio General del Medi Natural del Department de Medi Ambient de la Generali- tat de Catalunya, Instituto Nacional de Investigacion y Tecnologia Agraria y Alimentaria and Ministerio de Ciencia e Innovacion of Spain.