Cell cycle
The cell cycle is a coordinated sequence of molecular events that regulate normal cell division and is comprised of four discrete phases termed M phase, S phase, G1 phase, and G2 phase.
Inclusive of these four active phases of cell division, there exists an additional quiescent phase known as G0 (Figure 3.1) (Golias et al., 2004; Schafer, 1998). In tissues that undergo rapid cell division, referred to as labile tissues, including the bone marrow and intestinal epithelium, a substantive fraction of resident cells is actively recruited into the cell cycle. In contrast, cells derived from organs that rest in a homeostatic state, referred to as stable tissues, remain in the non-proliferative G0 phase, unless cell replication is stimulated through mitogenic signals that recruit cells into the G1 phase. 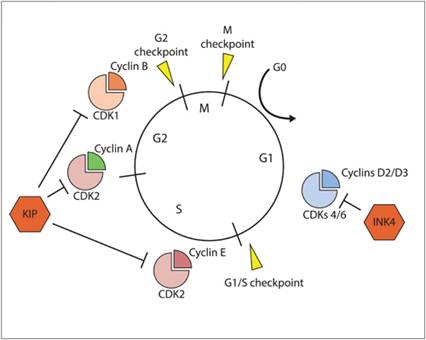
Figure 3.1 The cell cycle and regulatory control points. The cell cycle is divided into four phases (G1, S, G2, and M) and the inclusion of an additional quiescent phase (G0). Three cellular checkpoints (G1/S, G2, and M) ensure that DNA is replicated and segregated to daughter cells with high fidelity. Sequential and orderly procession through the cell cycle is regulated by activities mediated by cyclin-CDKs.
Coordinated progression through the cell cycle is mediated by molecular checkpoints under the control of enzymes called cyclin-dependent kinases (CDKs). The activities of specific CDKs are responsible for regulating the passage of cells through discrete phases of the cell cycle (Figure 3.1) (Grana & Reddy, 1995; Satyanarayana & Kaldis, 2009). Stringent control of the cell cycle is necessary to minimize the generation of heritable errors in DNA replication and the avoidance of genomic instability, and the activities of CDKs are tightly regulated through the following redundant molecular mechanisms:
· Full enzymatic activities of CDKs require the coupling/pairing of preferential cyclin proteins with specific CDK subunits.
Although the transcription of CDK subunits is relatively constant throughout the cell cycle, the translational stability of cyclin proteins is highly variable (except for Cyclin D), and mediated by proteasome degradation pathways, and selective cyclin protein expressions are specific to different phases of the cell cycle. Given the cyclic nature of cyclin protein stability, the activities of CDKs can be tuned and regulated by cyclin expression. Specific couplings of cyclins and CDKs are listed below and their cell cycle regulating activities are summarized in Table 3.1.Table 3.1 Cell cycle and associated regulatory kinases
| Cell cycle phase | Cyclin | Cyclin-dependent kinase |
| G0 | C | CDK3 |
| G1 | D and E | CDK2, CDK4, and CDK6 |
| S | A and E | CDK2 |
| G2 | A | CDK2 and CDK1 |
| M | B | CDK1 |
o D-type cyclins (D1, D2, and D3) partner with CDK4 or CDK6.
o E-type cyclins (E1 and E2) partner with CDK2.
· A-type cyclins partner with CDK2 or CDK1.
o B-type cyclins partner with CDK1.
· Phosphorylation positively and negatively regulates the activities of cyclin/CDK complexes. Full CDK kinase activity requires an activating phosphorylation and is performed by CDK-activating kinases. Dephosphorylation of inhibitory sites through the activity of phosphatases is necessary to fully activate cyclin/CDK complexes too. As such, phosphorylation and dephosphorylation serve as molecular switches to control cyclin/CDK activities.
· Activity of cyclin/CDK complexes is directly regulated by two discrete classes of inhibitory proteins known as CDK inhibitors (Harper, 1997), which include the following:
o INK4 proteins. This family of proteins specifically binds to CDK4 monomers and distorts the cyclin-binding domain, reducing the affinity for CDK4 to bind with D-type cyclins (Carnero & Hannon, 1998).
Specific proteins belonging to the INK4 family include p16INK4A, p15INK4B, p18INK4C, and p19INK4D. Given their binding to CDK4 monomers, INK4 proteins regulate cell cycle progression restricted to the G1 phase and principally regulated genomic integrity prior to the S phase of the cell cycle.· CIP/KIP proteins. This family of proteins bind to cyclin/CDK heterodimers and obstruct the ATP-binding site in the catalytic cleft of CDKs (Hengst & Reed, 1998; Nakayama & Nakayama, 1998). Specific proteins belonging to the CIP/KIP family include p21Cip1, p27Kip1, and p57Kip2. The CIP/KIP proteins are promiscuous with their binding to cyclin/CDK heterodimers, and therefore can regulate the cell cycle at all checkpoints, and are not restricted to being operative only in the G1 phase of the cell cycle like the INK4 family of proteins.
In normal cells, the p53 protein is principally responsible for controlling cell cycle progression. Following DNA damage, p53 behaves as a transcription factor for scores of genes and promotes the gene expression of the CIP/KIP family of CDK inhibitors (Boulaire et al., 2000; Gartel et al., 1996). The multi-layered regulation of the cell cycle is critical for ensuring that cells harboring DNA damage will be arrested prior to DNA synthesis and permits the correction of aberrant genomic information through the activities of DNA repair systems. The regulatory checkpoints within the cell cycle minimize the chances for perpetuating heritable genome mutations (initiation phase of the malignant transformation cycle) that could be passed onto daughter cells and thus serve as a principal safeguard against the initiation and continued development of cancer.