Centrioles
These structures are composed of a short cylindrical arrangement of microtubules. They occur as a pair near the center of the centrosome. The centrosome acts as an organizing center for building microtubules and serves as the spindle pole during mitosis.
The two centrioles are oriented at right angles to each other in an L-shaped pattern. The centrosome duplicates and divides into two equal parts during the interphase period of cell division so that each half contains a duplicated centriole pair. The daughter Centrosomes migrate to either side of the nucleus at the start of mitosis to form the two poles of the mitotic spindle. The granular appearance of the cytoplasm in the region surrounding the centrioles is believed to result from a complex of proteins and fibers involved in the movement and duplication of the centrioles. Each centriole resembles a pinwheel made of each of nine triplets of microtubules arranged to form a hollow tube. They also form the basal structures of cilia and flagella and are usually called basal bodies based on the original interpretation that they were different from centrioles. It is now known that the structures are related. However, whereas the centriole has the pattern of nine microtubule triplets, the basal bodies have an arrangement of nine pairs of microtubules oriented around a single pair of microtubules in the center. Although each of the central microtubules is complete, the outer doublets are fused so that the pair shares a common layer. This 9 + 2 organization is characteristic of most if not all types of cilia and flagella. The bending of the central core of the structure, which is called the axoneme, produces the movement of the cilia or flagella.Figure 2.12 illustrates a cross section through the tail of a bovine sperm cell. The arrangement of doublets of microtubules around a central pair of microtubules is apparent.
The diagram shows that the microtubules are linked with molecules of the protein nexin to form the circular array. The doublets are decorated with inner and outer arms composed of the protein dynein and anchorage proteins that position the outer doublets around the central core. In sperm cells, the asymmetric arrangement of filaments around the outside of the axoneme allows the movement of the tail to follow a figure-eight pattern of motion characteristic of bovine sperm cell motility.Nuclear structure
With the exception of mitochondrial DNA, the majority of the DNA in eukaryotic cells is confined to the nucleus. Comparable to the double membrane of the mitochondria, the nucleus is delimited by a double membrane called the nuclear envelope. Unlike mitochondria, these membranes are interspersed with nuclear pores. A complex of proteins populates these areas and acts to control passage of molecules into and
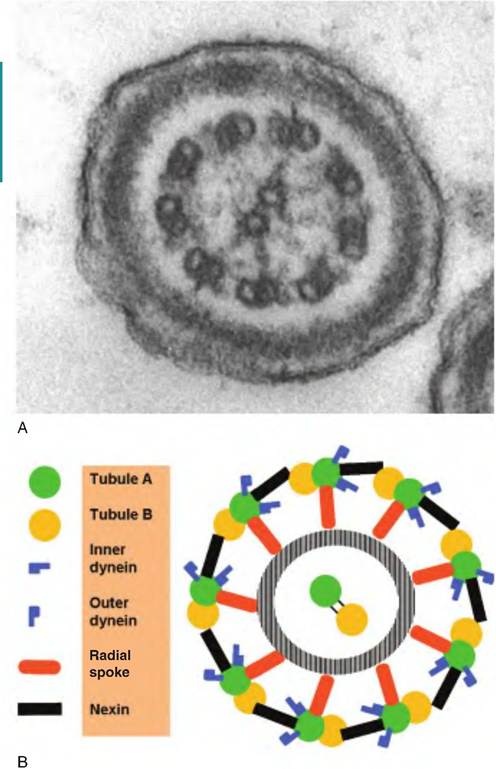
Fig. 2.12. Centrioles and cellular movement. Panel A shows a cross section through the tail of a bovine sperm cell. Panel B gives a diagrammatic representation of the molecular and associated proteins.
out of the nucleus. This is important since the DNA or genes must be transcribed to generate molecules of messenger RNA. Newly synthesized mRNA molecules are processed and transferred to the cytoplasm for translation by the ribosomes to create the myriad of proteins needed by the cell. The presence of the nucleus in eukaryotic cells allows processing related to DNA synthesis and gene activation to be localized away from other activities in the cytoplasm. This likely serves to minimize possible disruption of these critical gene-related activities. In short, the eukaryotic cells have evolved with the creation of numerous membrane-bound organelles that allow for segregation of many specific, divergent, biochemical reactions. This is believed to increase metabolic and biochemical efficiency of eukaryotic compared with prokaryotic cells.
The nucleus houses all of the chromosomes and therefore the genes.
Fortunately for most cells, only a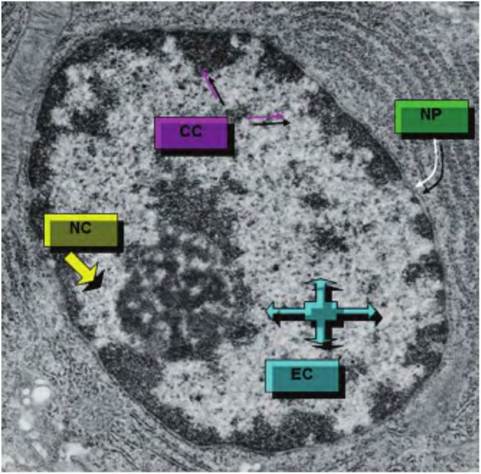
Fig. 2.13. The nucleus of epithelial cell is shown. Regions of condensed chromatin (CC) and extended chromatin (EC) are indicated along with nuclear pores (NP) and the nucleolus (NC).
fraction of the total DNA is being actively utilized at any given moment. For example, although all cells would contain the gene's copies for making the milk proteins or for synthesizing lactose, these genes would only be activated in the epithelial cells of the lactating mammary gland. Many other genes are activated only at a particular developmental period or in response to very specific stimuli. Consequently, much of the DNA is tightly compacted in the nucleus.
Most cells have a single nucleus, but there are exceptions. Skeletal muscle cells, bone osteoclasts, cardiac cells, and some liver cells are multinucleated. This is usually associated with cells that have a larger-than- normal cytoplasmic volume. With the exception of mature red blood cells of mammals, all cells are nucleated, and even these cells have the nucleus until late in their developmental sequence. Of course, without the nucleus and the genes necessary for protein synthesis, these cells cannot replace proteins that are progressively degraded by normal functioning. Although it is expected that the appearance of the nucleus would change dramatically during cell division, even in nondividing cells (so-called interphase or G0 phase of the cell cycle), there are distinct differences between cell types. These differences can be useful to identify some cell types. For example, plasma cells have a distinct pattern of condensed chromatin around the periphery of the nucleus resembling a clock face. Neutrophils have elongated, Iobed nuclei that make the appearance of these cells unique.
Figure 2.13 shows an electron microscopy section through the nucleus of an epithelial cell.
During most of the life of the cells, the DNA is in a complex with strongly basic proteins called histones, some nonhistone proteins, and a small amount of RNA. This combination of proteins and DNA is called chromatin. While the double helix structure of DNA is widely familiar, the degree of order or compaction of the DNAwithin the chromatin matrix varies. Compaction is extreme in cells that are preparing for the final stages of mitosis as the chromatin appears as distinct pairs of chromosomes. However, even in cells in G0, degrees of chromatic condensation can be seen. Darkstaining areas indicate regions containing condensed, presumably inactive, chromatin. Lighter areas contain more active, extended chromatin. Clumps of condensed chromatin often appear around the periphery of the nucleus (peripheral chromatin) along with scattered islands of condensed chromatin throughout the nuclear space. The interphase nucleus also has a protein network called the nuclear matrix. Much of this material appears as a thin, interwoven layer (the nuclear lamina) that adheres to the internal surface of the nuclear envelope. This provides support and anchorage for the nuclear pores. An extension of the nuclear lamina radiates into the interior of the nucleus. This layer also interacts with a similar lamina that surrounds the nucleolus. Together they regulate nuclear shape, reinforce the inner membrane of the nuclear envelope, secure the location of nuclear pores, and anchor condensed chromatin to the nuclear envelope. This organization is maintained except during mitosis. It is accepted that maintenance of the nuclear matrix is essential for routine gene transcription.