Microfilaments, microtubules, and intermediate filaments
It was originally assumed that the cytoplasm of the cell was essentially a water-filled space with multiple dissolved substances. However, appropriate fixation and embedment techniques for electron microscopes led to the realization that the cytoplasm contains an elaborate array of structures that make up the cytoskeleton of the cell.
This does not mean that cells are rigid, but microtubules, microfilaments, and intermediate filaments of the cytoskeleton provide an unexpected structure and organization to the cell cytoplasm. Some of these organelles are for communication between the cell surface and interior, for transport of vesicles to be secreted, for cell division, or for cell adhesion.Microtubules are the largest of these organelles and as the name suggests are hollow tubes composed of a and β subunits of the globular protein tubulin. They are slender with an outside diameter of 25nm. When cut in cross section they appear as small circles with 13 subunits of tubulin around the circumference. Its cylindrical structure develops as heterodimers of tubulin pack around a central core, which appears as a space in electron micrographs. At 37oC, purified tubulin polymerizes into microtubules in vitro in the presence of Mg and GTR Several antimitotic cancer drugs (colchicine and it relatives) act by interfering with tubulin polymerization. Another antimitotic drug, taxol, stabilizes microtubules and arrests cells in mitosis. These effects demonstrate the critical role of microtubules in cell division. Polymerization of tubulin to form microtubules occurs initially in a region near the nucleus called the centrosome. This has been demonstrated most clearly in cultured cells first treated with colchicine to disrupt the microtubules. After various periods of time, groups of cells were fixed, and the microtubules were stained by using fluorescent-tagged antibodies against tubulin.
When the drug is removed, new microtubules can be seen growing out from the centrosome to create a star- like structure called an aster. The microtubules thenelongate toward the outer regions of the cell to reestablish the microtubule network.
It is also known that disruption of microtubules dramatically impairs cellular secretion. For example, intramammary infusion of colchicine into the lactating mammary gland virtually stops milk secretion, but once the treatment is ended, milk secretion rapidly returns to normal. This demonstrates the requirement for microtubules for trafficking and exocytosis of secretory vesicles. An increase in the relative abundance of microtubules in mammary epithelial cells corresponds with increased milk secretion following parturition or increased secretory activity in other epithelial cells. Two families of microtubule-dependent motor proteins, kinesins and dyneins, are involved in organelle transport in the cytoplasm, in mitosis, and in movement of vesicles of neurotransmitter from sites of synthesis in the cell body to sites of release at the ends of axon terminals. Table 2.3 provides an example of changes in microtubule number and orientation related to cell function, and Figure 2.11 shows the appearance of microtubules in the mammary cells of a lactating cow.
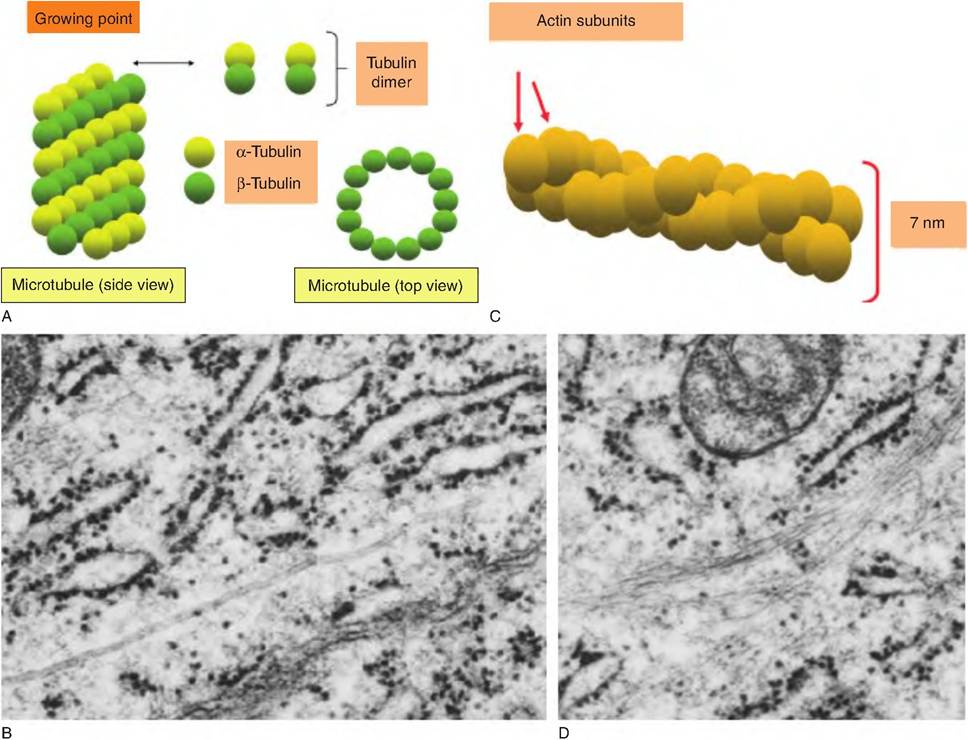
Fig. 2.11. Microtubules and microfilaments. Panel A illustrates the organization of a microtubule and its development from dimers of α- and β-tubulin. Panel B shows a transmission electron microscopic view of the apical region of an epithelial cell from the mammary gland of a lactating cow. The arrows indicate microtubules that have been cut longitudinally. Panel C shows the helical organization of monomers of actin arranged to create microfilaments. Panel D shows bundles (arrows) of microfilaments in secretory epithelial cells.
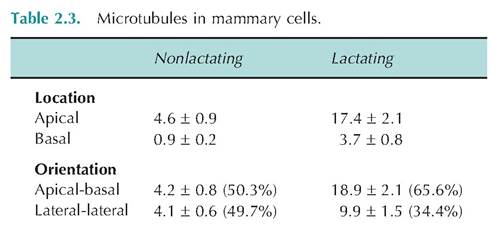
Average number of microtubules in the apical or basal cytoplasm observed in an apical to basal or lateral to lateral orientation with respect to the plasma membrane in mammary epithelial cells of nonlactating and lactating cows.
Adapted from Nickerson et al. (1982).Microfilaments are the smaller, thin components of the cytoskeleton composed of the protein actin. In many cells, actin accounts for 5% or more of the total protein in the cell. Actin can exist as a monomer or, similar to the tubulin of microtubules, can polymerize to form thin thread-like structures (~7 nm in thickness) called filamentous actin. In most cells, about half of the actin is present in the monomeric conformation because it is bound to the regulatory protein thymosin. Rapid changes in rates of polymerizationdepolymerization produce changes in the cell surface that produce Iamellipodia (essentially cell projections) and ultimately cell movement and migration. Specific arrangements of microfilaments within the cytoplasm can be induced by the activation of cell surface receptors. This is important for the action of highly mobile phagocytic cells of the immune system. Bundles of microfilaments are also found near cell surfaces and are highly ordered within the microvilli of absorptive epithelial cell layers. In this way bundles of actin filaments provide structural integrity for the microvilli. This is functionally significant because the adaptation of having the microvilli on the surface of an intestinal epithelial cell, for example, markedly increases the surface area available for absorption. A single intestinal enterocytes has several thousand microvilli. Actin filaments do not act independently, rather a variety of actin-binding proteins control rates of filament formation and creation of the specific types of filament groupings. For example, cross-linked microfilaments can form loose gels or rigid bundles to anchor plasma membranes.
Intermediate filaments are less labile than microfilaments or microtubules and are more elemental members of the cytoskeleton. The protein structure of these filaments can vary between cell types, but the proteins are bound together something like a braided, woven rope. These filaments are called neurofilaments in nerve cells and keratin filaments in many epithelial cells. Regardless, they provide additional support for the cell. They are especially important in the creation of desmosomes. Desmosomes are a type of anchoring junction that serves to hold adjacent epithelial cells together. In these regions, the plasma membranes of the neighboring cells do not touch, but linker proteins (cadherins) extend outward from the desmosomal plaque of each cell. On the cytoplasmic side of the plaque in each cell, intermediate fibers extend into the cytoplasm of the cell to provide interactions with other Cytoskeletal elements and additional support. Types of cell junctions will be considered in other chapters.