Cellular organelles
Structures found inside cells are called organelles. Examples include the nucleus, mitochondria, and ribosomes. Most organelles are membrane covered. Other organelles, secretory vesicles and lysosomes, for example, are unique because of their membranebound contents.
Thus, understanding membranes is important to understanding physiology. We begin with lipids and especially phospholipids. Lipids are a very heterogeneous group of molecules, but common attributes include (1) being practically nonsoluble in water but (2) being soluble in nonpolar organic solvents such as ether, ethanol, or chloroform. Lipids include fats, oils, waxes, and related compounds. Figure 2.2 shows the general structure of molecules necessary to produce common fats called triacylglycerols or triglycerides.Neutral fats are esters composed of two building blocks—glycerol and one of any number of different fatty acids. Glycerol is a 3-carbon alcohol that is most
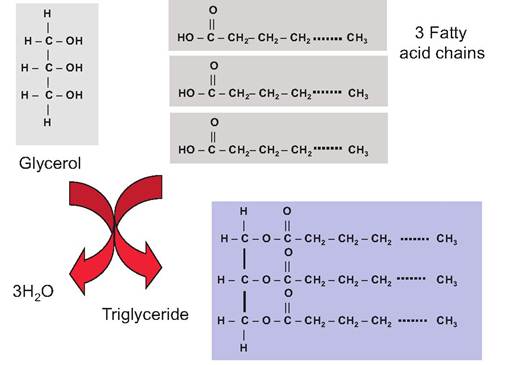
Fig. 2.2. Triglyceride synthesis and structure.
Table 2.1. Common saturated fatty acids.
| Fatty Acid | Formula | Attributes |
| Acetic | CH3COOH | Major encl product of rumen fermentation, energy source for ATP production |
| Propionic | C2H5COOH | End product of rumen fermentation, major precursor for gluconeogenesis |
| Butyric | C3H7COOH | Major end product rumen fermentation |
| Caproic | C5H11COOH | Minor end product rumen fermentation |
| Caprylic | C7H15COOH | Small amounts in many fats |
| Palmitic | C15H31COOH | Common in all animal and plant fats |
| Stearic | C17H35COOH | Common in all animal and plant fats |
often derived from the catabolism of the common hexose sugar glucose.
Fatty acid molecules are linear hydrocarbon chains with a carboxylic acid moiety at one end. This residue or group is the most reactive or functional part of the molecule. Fatty acids vary in length, but the glycerol backbone of the triglyceride is constant. Fatty acids also vary with respect to the number of double bonds between carbon atoms. Those with no double bonds are called saturated fatty acids, those with a single double bond are monosaturated, and those with more than one are polysaturated fatty acids. The degree of saturation and length of the fatty acids affect their properties. For example, the shorter chain members the membrane are positioned so that fewer polar amino acids occur within the central hydrocarbon tailsof the fatty acid chains, with polar amino acids located with the polar heads or aqueous surfaces of the membrane. Examples of complex plasma membrane proteins include receptors for hormones or growth factors (GFs) and those required for transport of metabolites and nutrients.
Cellular membranes are fluid, dynamic, and active structures. Membrane components are also interchangeable between many cellular components. For example, in the mammary gland of a lactating mammal, milk components are packaged into secretory vesicles within the Golgi apparatus. These product-containing vesicles progressively make their way to the apical surface of the cell where their contents are released into the storage spaces of the mammary gland by the process of exocytosis. The membrane surrounding the vesicles becomes part of the plasma membrane. Furthermore, lipid droplets synthesized in the cells progressively enlarge and also migrate to the apical surface of the cells for secretion. However, in this case, the droplets literally begin to protrude from the cells and become surrounded by the plasma membrane. This continues until droplets pinch off with the former plasma membrane now encapsulating the droplet. The membrane is now referred to as the milk fat globule membrane, but its origin was the plasma membrane of the cell.
Figure 2.4 illustrates the organelles and secretion activity of such a mammary epithelial cell. Similar events occur in many other secretory cells, for example, pancreas, liver, salivary gland, and pituitary gland.Microscopy techniques
Beginning with invention of the light microscope in the 1600s and progressive improvements in cell preservation, techniques to embed tissue in materials for sectioning, and staining to identify specific cellular components, much has been learned regarding cell structure and function. However, even simple smears of dislodged isolated cells can be very useful in physiological or clinical situations. The Pap smear is routinely used in women's health to monitor the cells of the cervix. The morphology of the cells is classified to determine if any of the cells appear to have precancer- ous attributes, for example, altered nuclear morphology or staining characteristics. Another example is the blood smear, that is, a small sample of blood is spread and dried on a microscope slide and then stained. Such smears are cover-slipped, and a differential count is performed. In this procedure, the slide is scanned in a standard pattern and the first 100 white blood cells encountered are identified (lymphocyte, neutrophils, etc.) and tabulated. This information is used to produce a distribution profile of the types of leucocytes in the
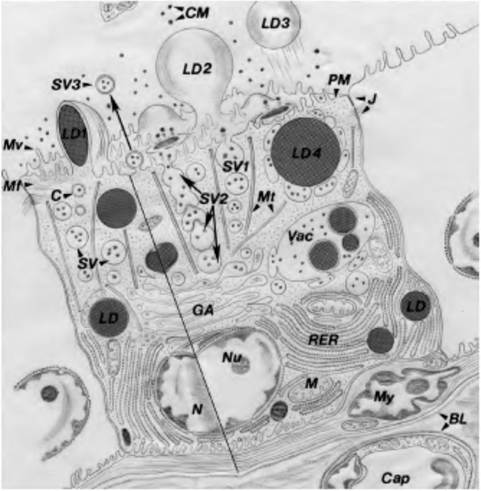
Fig. 2.4. Diagram to illustrate major pathways for cellular synthesis and secretion. Milk precursors in capillaries (Cap) are transported across the endothelial cells and basal lamina (BL) to the interalveolar connective tissue. Nutrients pass across the alveolar BL and/or myoepithelium (My), the basal plasma membrane (PM), and into the cytoplasm. Milk proteins are synthesized in the rough endoplasmic reticulum (RER), enter the RER Iumena, and are transported to the Golgi (GA) for processing and packaging. In typical exocytosis, the secretory vesicles (SV) with casein micelles (CM) and lactose leave the Golgi, translocate to the apical PM, and release contents of the vesicle (SV1).
Alternatively, vesicles can fuse to form chains for secretion (SV2) or fuse with release of double membrane-bound micelles (SV3). Milk lipid is synthesized in the region of the RER and as droplets grow, they also translocate to the apical PM. These droplets are enveloped by PM and protrude from the cell (LD1), and are pinched off from the cell into the lumen (LD2) and into the lumen (LD3). It is also possible that SV can fuse around lipid droplets, with other droplets, and with the apical PM in groups (LD4). Lipid droplets might also be released via coalesced secretion vacuoles (Vac). Other features include mitochondria (M), nucleus (N), nucleolus (Nu), microtubules (Mt), microfilaments (Mf), coated vesicles (C), and Iightjunctions (J). Diagram modified from Nickerson and Akers (1984).sample. For example, the horse averages about 55% neutrophils, 35% lymphocytes, 5% monocytes, 3% eosinophils, and 1% or fewer basophils. Changes in these proportions can reflect various diseases. What would be your prediction about a classmate with mononucleosis or a cat with leukemia?
In dairy animals, mastitis (inflammation of the mammary gland) status is routinely evaluated by the presence and number of leukocytes in the milk. The technology used is based on a well-characterized relationship between cell number and the amount of a specific dye that binds to DNA. As the cell number increases in the milk sample, the amount of dye binding
Table 2.2. Relationship between MSCC, DHI cell counts score, and milk production in dairy cows.
| MSCC | DHI Score | Milk Yield (kg∕d) | Milk Yield 305d (kg) |
| 12,500 | 0 | 29.2 | 8906 |
| 25,000 | 1 | 28.6 | 8723 |
| 50,000 | 2 | 28.0 | 8540 |
| 100,000 | 3 | 27.4 | 8357 |
| 200,000 | 4 | 26.9 | 8205 |
| 400,000 | 5 | 26.2 | 7991 |
| 800,000 | 6 | 25.4 | 7747 |
| 1,600,000 | 7 | 24.6 | 7503 |
| 3,2OO,OOO | 8 | 23.6 | 7198 |
| 6,400,000 | 9 | 22.5 | 6863 |
Adapted from Jones et al.
(1984).
increases proportionally. Although these assays are now automated, a milk smear is used to calibrate these machines. Table 2.2 illustrates the relationship between milk somatic cell count and milk production.
In some experimental situations, it is useful to know the stage of the estrus cycle. For many laboratory animals, it is problematic to collect a sufficient number of repetitive blood samples to measure reproductive hormones (estrogen, progesterone, follicle-stimulating hormone, luteinizing hormone) for this purpose. However, daily vaginal smears can be used to evaluate the stage of the estrus cycle. The number of cornified epithelial cells and leukocytes varies according to the stage of the estrus cycle so that changes in these cellular profiles can be used to determine the stage of the estrus cycle.
Although information obtained from smears of various cells is useful, the technique is limited since most cells are part of tissues. More importantly, the organization and differentiation of the various cell types and their products is fundamental to understanding the physiology of a tissue or organ. For this evaluation, it is necessary to infiltrate the tissue and cells with a medium that is sufficiently solid to allow sections thin enough for light to penetrate to be prepared. The most common embedding medium is paraffin wax. Since tissues and cells are largely water based, fresh tissues are first preserved or fixed in an aqueous solution containing chemicals that cross-link major cell structures and macromolecules. These include formalin, formaldehyde, glutaraldehyde, and others. After a period of fixation, the tissues are dehydrated by transferring the tissue through a series of increasing concentration of ethanol, then into xylene, a mixture of xylene and paraffin, and finally, pure paraffin. This gradual process allows the water to first be replaced by ethanol, then the ethanol by xylene, and the xylene by paraffin. The tissue blocks are submerged in additional paraffin in a mold and allowed to harden.
This can be imagined by thinking of the string in the center of a candle as being the processed tissue. If the candle is carefully sliced in cross section, a piece of string would be in the center of each slice, representing the fixed embedded tissue. The slicing of the tissue blocks needs to be relatively precise and uniform. Embedded blocks of tissue are sliced in a machine called a microtome. The machine uses a thin steel blade, much like a razor blade, and the sections, which are cut one at a time, usually come off in a ribbon. The sections are floated on a water bath and then transferred to a microscope slide. Once the sections are dried, the slides are generally dipped in xylene and processed back to aqueous environment to allow the sections to be stained. The staining allows structures to be seen in a standard bright field microscope. H&E, or hematoxylin and eosin, are very common stains. This technique makes the cell nuclei dark blue, the cytoplasm various shades of blue to pink, and extracellular components pink to red. Most histological slides used in physiological classes are H&E stained. Many other stains have been developed for specific uses, and examples are given in various sections of the text. One especially exciting recent innovation in tissue staining is the use of specific antibodies to localize proteins within particular cells or even within particular cellular organelles. Figure 2.5 shows tissue blocks, molds, and processed sections for tissue embedded in paraffin.Despite the widespread use of paraffin-embedded tissue sections and the rich experimental and pathological history of this technique, there are serious limitations. One of these is that tissue sections thinner than about 5 μ cannot be easily prepared. A reasonable approximation of an epithelial cell is about IOxlOx 10 μm. This means that for the study of intracellular organelles, the sections are thick so that it becomes difficult to distinguish these structures. These limitations lead to the development of plastic resins that could be used for embedding cells and tissues. With subsequent development of specialized microtones designed to use pieces of fractured glass or even diamond knives, it became possible to section fixed tissues embedded in plastic very thin indeed. In fact, for light microscopic study, sections of 0.5-1 μ in thickness can be easily prepared. To distinguish sections from the paraffin blocks from those in plastic, they are often called semithin sections.
Perhaps, more importantly, these breakthroughs allowed even thinner sections to be examined using the electron microscope. While a detailed consideration of the electron microscope is beyond the scope of our text, some analogy with the readily understood light microscope is useful. The standard compound microscope is essentially a two-part magnifying system

C D
Fig. 2.5. Preparation of tissues for microscopic examination. Panel A shows tissues that have been embedded in paraffin (upper) or in plastic (lower) for preparation of sections for light microscopy. The tissue in the paraffin block is visible as a faint yellow mass (arrow) in the center. The width of the block is about the size of a U.S. quarter. The tissue in the lower plastic block is not visible but occupies a position near the center of the block. Panel B shows a view of a typical microtome for preparation of paraffin-embedded tissues. Panel C shows a paraffin- embedded tissue block mounted in the microtome. With each up and down motion of the microtome, the block of tissue moves past a stationary knife so that a ribbon of serial sections (typically ~5μm thick) is cut. The ribbon of sections is transferred to a water bath (panel D) and ultimately floated onto a microscope slide. The sections are allowed to dry, then the paraffin is removed, the sections hydrated, stained, and subsequently cover-slipped for examination. Processing of tissue embedded in plastic resins follows a similar procedure, but thinner sections (-0.5μm) can be prepared for light microscopy, and with the proper microtome, sections thin enough for electron microscopy can be prepared.
in which the specimen is first magnified by the lens in the objective barrel and secondly by the lens of the eyepiece or ocular. The total magnification is the product of the magnification of the objective lens used and that of the eyepiece. For example, using a 20? objective lens with a typical IOx eyepiece produces an image that is 200-fold greater than the original. The specimen is placed on a stage below the objective lens. Light is then directed from a light source, through an aperture, then a substage condenser, and through the specimen. Light rays from the specimen pass through the objective lens and are focused for view through the eyepieces. This is accomplished mechanically by raising or lowering the position of the objective lens relative to the specimen. Resolution is the degree of separation that can be seen between adjacent points in a specimen—in other words, the degree of detail. The smaller the distance that can be distinguished between two points, the greater the detail in the image. With the unaided eye, points appear as independent structures only if a distance of 0.2 mm or 200 μm separates them, but with a good microscope, points as close as 0.25 μm can be distinguished. Ultimately, resolution of the bright field microscope is limited by the wavelength of light and sample preparation. The maximum useful magnification of the light microscope is about 1,400-fold. Images can be reproduced to larger sizes in the printing process, but this does not increase true resolution.
In the electron microscope, a beam of electrons replaces the beam of light. The sample (now only about 900 nm in thickness) is positioned on a copper grid, and the grid is inserted into a sealed chamber and placed under vacuum. A beam of electrons passes through the sample. To increase the electron density of the sample, the tissue is treated with heavy metals (usually lead citrate or uranyl acetate) that bind with macromolecules in the sample and improve sample resolution. The electron beam penetrates the tissue located in the open spaces of the grid, and the image produced is brought into focus by altering the voltage applied to a series of electromagnets located on either side of the column which houses the electron beam. The image is viewed first on a phosphorescent screen, and the image is saved by positioning and exposing film. The film is developed, and images of the specimen are prepared by making photographic prints from the film. The process as described is called transmission electron microscopy. A similar process called scanning electron microscopy relies on images produced by coating surfaces often with a thin layer of gold. Detailed images of intracellular structure became possible only with electron microscopy; several examples are included along with descriptions of cell organelles. Figure 2.6 shows a block of tissue prepared for study in an electron microscope, one of the small copper grids, and an example of an exposed, developed photographic plate.
Organelles of the cytoplasm
While many complete texts are devoted to aspects of cell and molecular biology, a basic appreciation of cell structure is important in understanding cellular physiology and ultimately tissue, organ, and systems-level physiology. It is imperative to appreciate that essentially all organelles are found in all cells. However, the total number and arrangement of these organelles vary markedly from cell type to cell type. Numbers can also change dramatically within a given cell type depending on the activity of the cell. This is often described as the degree of differentiation of the cell. For example, mammary alveolar epithelial cells taken from a nonlactating pregnant animal have a very different complement of cellular organelles than cells collected during lactation. This is a reflection of differences in activity between these stages of development.

Chapter 2
Fig. 2.6. Comparisons between tissue preparation for light microscopy and transmission electron microscopy. The upper row shows samples prepared for paraffin, plastic, and electron microscopy. The holder in the center of the upper row is designed to hold a series of small copper grids (just below the coin to the right) that have very small ribbons of plastic embedded tissue. The tissue fragment is first embedded in a bullet-shaped mold that is filled with plastic resin. Once the plastic is polymerized, the mold is removed, and the hardened plastic with the tissue, now located at the end of the bullet, is sectioned. The ribbon of sections is then floated onto the copper grid. As can be appreciated from the scale offered by the coin, these ribbons must be maneuvered via a dissecting microscope attached to the microtome. The middle row shows a slide with a ribbon of unstained tissue compared with deparaffinized, stained tissue. The plate to the right of the middle row is an exposed, developed film plate (negative) from the electron microscope. The lower portion of the figure shows a part of a glossy print made from this negative (the tissue is from the pituitary gland).
Certainly, biochemical differentiation of the secretory cells is required for onset of milk secretion. However, the cells must also acquire the structural machinery needed to synthesize, package, and secrete milk constituents. When alveolar cells first appear during mid-gestation, they exhibit few of the organelles needed for copious milk biosynthesis or secretion. The cells are characterized by a sparse cytoplasm with few polyribosomes, some clusters of free ribosomes, limited rough endoplasmic reticulum (ER), rudimentary Golgi usually in close apposition to the nucleus, some isolated mitochondria, and widely dispersed vesicles. Individual cells often contain large lipid droplets (especially during later stages of gestation) that along with irregularly shaped nuclei account for much of the cellular area. Electron microscopic studies solidified the dramatic structural changes in the alveolar secretory cells at the onset of lactation. These differences are illustrated in Figure 2.7 and Figure 2.8. Also, it is important to appreciate that similar changes in cell differentiation occur in many
Chapter 2
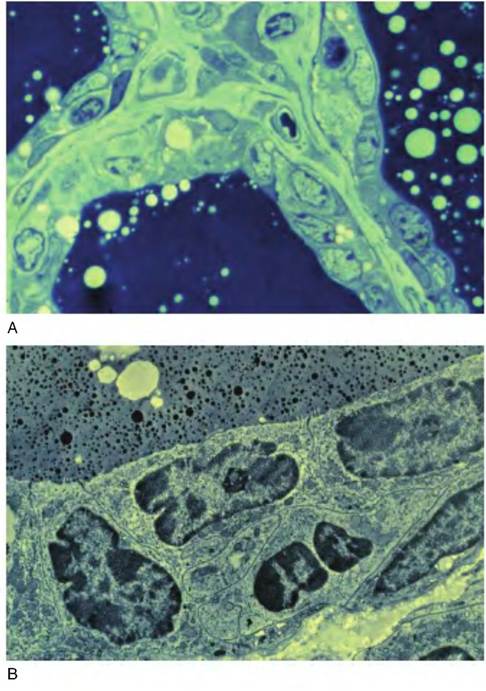
Fig. 2.7. Cellular differentiation examples. Shown are companion light (upper) and transmission electron microscopic (lower) images of bovine mammary tissue of a nonlactating cow in late gestation. Note the relatively large proportion of cell area occupied by the nuclei of the cells, relative lack of cellular organelles, absence of cellular polarity, and minimal evidence of secretion. These cells are minimally active, so there is a correspondingly minimal complement of cellular organelles.
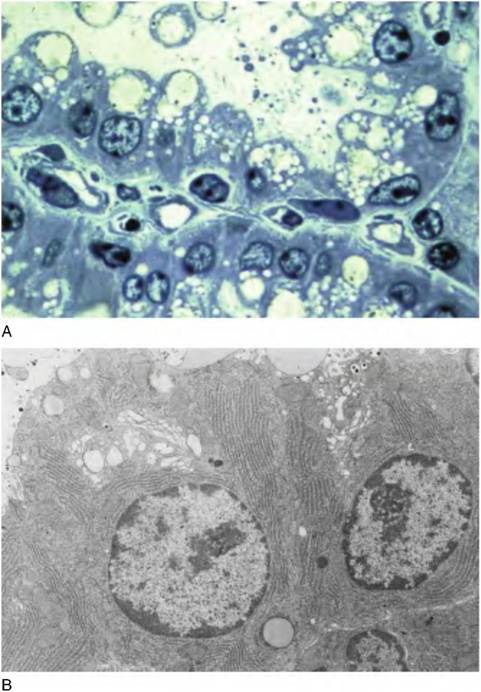
Fig. 2.8. Examples of light and transmission electron microscopic images of lactating bovine mammary tissue. The cells typically have rounded, basally displaced nuclei, scattered fat droplets, and evidence of secretions (panel A). Note the Iacy appearance of the apical ends of these well-differentiated, polarized cells. Conformation that the Iacy appearance indicates the presence of abundant secretory vesicles is evident in the EM view of portions of two secretory cells in panel B. Panel A is unpublished, and panel B is adapted from Nickerson and Akers (1984).
epithelial cells, that is, various glands, intestines, and pancreas; the mammary gland makes a convenient and dramatic example.
Mitochondria
Called the powerhouses of the cell, mitochondria provide most of the ATP necessary for energyrequiring reactions. Two membranes enclose the generally elongated, thin, hot dog-shaped mitochondria. The outer membrane smoothly encapsulates the organelle, but the inner membrane is thrown into multiple folds that form partitions called cristae. Surfaces of the cristae are studded with embedded enzymes that interact with the internal gel-like matrix of the mitochondria. Although details are discussed in subsequent chapters, as energy-yielding nutrients are metabolized, intermediate products from the digestive process, for example, glucose, amino acids, fatty acids, are converted into compounds that enter the mitochondria. These compounds are Catabolized to carbon dioxide and water by the action of the mitochondrial enzymes, and a portion of the bond energy is captured and used to attach phosphate groups to ADP to generate ATP. This is called aerobic respiration because it requires oxygen. Essentially, the need for oxygen is explained by the fact that it is required for production of adequate amounts of ATP
Mitochondria are very complex organelles. They have their own DNA (derived incidentally from the mother) and RNA. As energy demand increases, mitochondria increase the density of cristae or undergo fission to create new mitochondria. Active cells such as those in muscle, pancreas, or the lactating mammary gland may have hundreds of mitochondria, but inac-
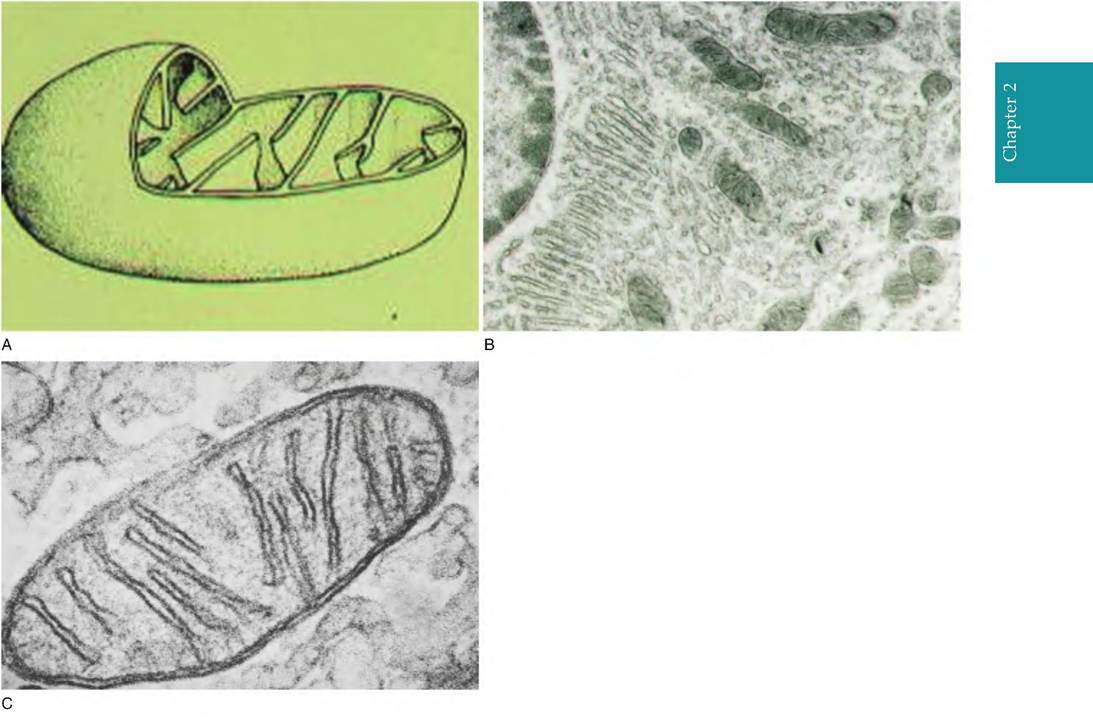
Fig. 2.9. Panel A shows a diagrammatic representation of prototypical mitochondria. The structure is clearly bounded by a double membrane with the inner membrane thrown into distinct folds or cristae. Panel B shows a group of mitochondria (arrows) in the basal region of a bovine kidney cell, and panel C illustrates a high-resolution image of mitochondria from the bovine ovary.
tive cells (nonlactating mammary gland) or quiescent lymphocytes, for example, have only a few. In living cells, the mitochondria can also change shape. Regardless of its particular morphology, the emergence of mitochondria was a major evolutionary event. Figure 2.9 illustrates typical mitochondria. It is widely believed that mitochondria arose from bacteria that invaded the ancestors of plant and animal cells.
Ribosomes
These small, dark-staining organelles are composed of proteins and a class of RNA called ribosomal RNA. Each of the ribosomes has two subunits identified based on size as 18 and 28s RNA. Ribosomes can appear singly as free structures in the cytoplasm or sometimes arranged along coiled loops of mRNA called polyribosomes. Alternatively, especially in cells that are synthesizing abundant amounts of protein for secretion, ribosomes are often attached to membranes to create rough endoplasmic reticulum (RER). As subsequently discussed, the ribosomes are the sites of protein synthesis. Because of the relationship between ER and the Golgi apparatus (Fig. 2.10), ribosomes of the RER allow for newly manufactured proteins to be packaged in secretory vesicles for secretion from the cell. Free ribosomes in the cytoplasm function to synthesize proteins destined to act within the cell.
Endoplasmic reticulum and Golgi apparatus
The endoplasmic reticulum (ER) is an interconnected network within the cytoplasm of the cell. It is a system of interconnecting membrane tubes or sheets that enclose fluid-filled spaces that appear in two variations: smooth or rough ER. It is also continuous with the nuclear membrane. Protein synthesis depends on three forms of RNA. These are (1) transfer RNA (tRNA), (2) ribosomal RNA (rRNA), and (3) messenger RNA (mRNA). When the mature mRNA reaches the cytoplasm, it binds to a small ribosomal subunit by base pairing to rRNA. The tRNA transfers amino
Chapter 2
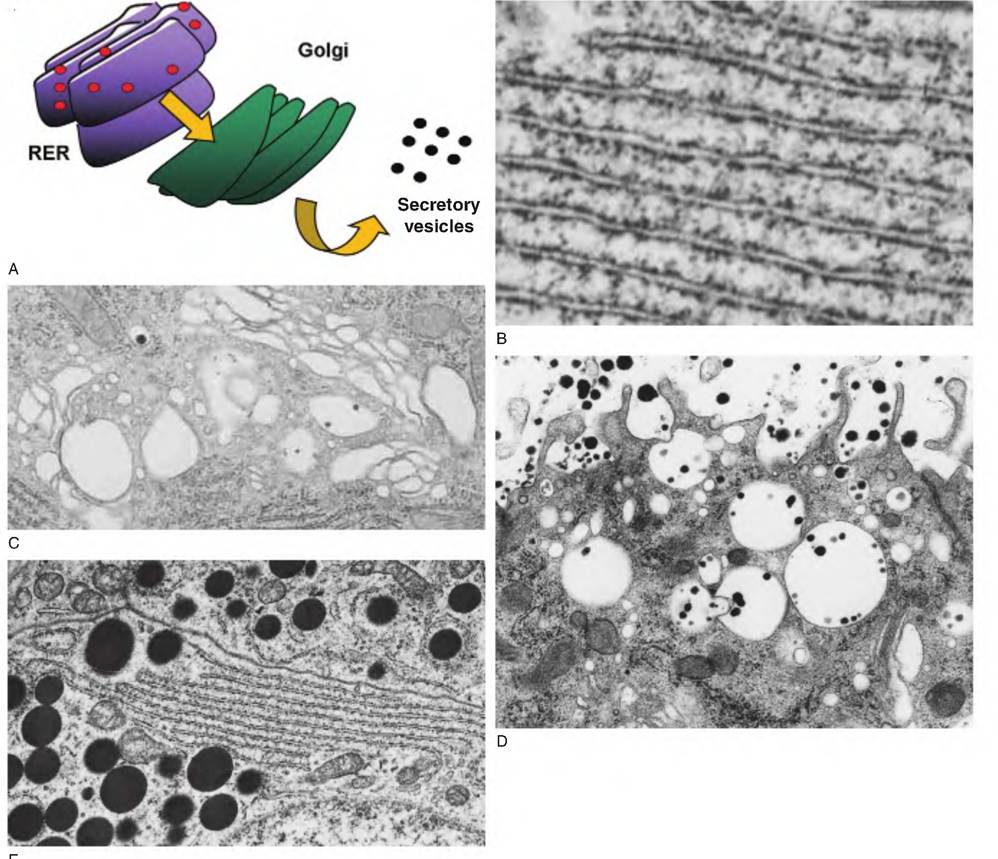
Fig. 2.10. Relationships between RER, Golgi, and secretory vesicles. Panel A shows the arrangement between ribosomes (red dots) on RER and movement of newly manufactured proteins into the cisternal space of the RER and then to the Golgi for packaging and appearance of secretory vesicles. Panel B shows a transmission electron microscopic view of RER, and panel C shows an array of Golgi membranes. Panel D shows secretory vesicles from epithelial cells in a mammary gland of a lactating animal. The dark, black granules are the casein micelles. Since lactose is also produced in the Golgi and packaged for secretion along with specific milk proteins, the vesicles appear swollen. This is because lactose cannot pass across the vesicle membrane so water is drawn osmotically into the vesicle. For other protein-synthesizing and secreting cells, the secretory vesicles more often appear densely compacted, with the vesicle membrane directly adjacent to the product.
Panel E shows secretory vesicles and Golgi area from bovine anterior pituitary cells. Notice the close apposition of the membrane surrounding the secretory granules.
acids to the ribosome. There are approximately 20 different types of tRNA, each capable of binding a specific amino acid. The linkage process is controlled by a synthetase enzyme whose action depends on the cleavage of ATP to form the peptide bonds between amino acids of the growing peptide chain. Once its amino acid is loaded, the tRNA migrates to the ribosome, where it moves the amino acid into position, based on the codons of the mRNA strand. The amino acid is bound to one end of the tRNA (the tail), but the other end of the molecule (the head) has a three- nucleotide base sequence (anticodon), which is complementary to the codon of the mRNA. For a given strand of mRNA, multiple ribosomes can become attached, and as the ribosomes move along the molecule, many chains of new protein can be made simultaneously. In fact, it is not uncommon to find polyribosomes in the cytoplasm. These are represented in transmission electron microscopic views of active cells by chains or coils of ribosomes seemingly organized in the cytoplasm.
However, proteins destined for secretion from the cell are synthesized by ribosomes attached to the endoplasmic reticulum. The mRNA for these proteins codes an initial short peptide sequence (signal peptide), which directs the growing peptide chains into the cisternal space of the endoplasmic reticulum. Because this space is continuous with the Golgi apparatus, proteins destined for secretion are vectored into Golgi for packaging into secretory vesicles and secretion from the cell by exocytosis. After synthesis in the RER, modifications to secretory proteins may also occur in the Golgi apparatus. These posttranslational modifications can markedly affect the structure of the protein. Common modifications include the addition of sugar or phosphate groups. Other components can also be added to developing secretory vesicles in the Golgi. For example, in the mammary gland, the milk sugar lactose is synthesized within the Golgi apparatus by the action of galactosyl-transferase and Oc-Iactalbumin.
Lysosomes and peroxisomes
Peroxisomes are intracellular vesicles containing a mixture of enzymes, namely, oxidases and catalases. Oxidases depend on the presence of oxygen to detoxify various noxious substances, for example, alcohols and aldehydes. They also convert toxic-free radicals into hydrogen peroxide for neutralization by catalase. Free radicals are very reactive substances known to alter the structure and function of a variety of regulatory molecules. Thus, the peroxisomes are essential to limit the free radical accumulation. Peroxisomes are abundant in liver and kidney cells, two organs recognized for their capacity to detoxify harmful substances.
Lysosomes also contain hydrolytic enzymes that are capable of digesting many cellular proteins. Known as suicide bags or sacs, inappropriate release of the contents of these organelles could destroy the cell. In fact, rupture of activated lysosomes is believed to be involved in some aspects of programmed cell death or apoptosis. Lysosomes are present in all cell types, but they are especially plentiful in neutrophils, macrophages, and other leucocytes. The acid hydrolases within the lysosomes function best in an acidic environment. Consequently, the lysosomal membrane contains hydrogen transport proteins that sequester hydrogen ions from the cytoplasm to maintain a low pH. Many cells are capable of capturing materials from near the cell surface by endocytosis. Vesicles produced in this manner can then fuse with lysosomes. Captured molecules can be digested by the acid hydrolyses and released into the cytoplasm for use by the cell or for excretion. This digestion process is especially important in macrophages and neutrophils since these cells actively engulf potentially harmful bacteria and other toxins. Destruction of these agents by the lysosomes is protective, and in the case of processed foreign proteins, fragments of the digested proteins are presented to other cells of the immune system to allow development of specific immunity. Lysosomes are also critical in the recycling of worn-out or nonfunctional organelles as well as a variety of metabolic actions, for example, release of thyroid hormones from storage.