Cryptosporidium cuniculus Infection: Cryptosporidiosis
The very large Cryptosporidium group, although morphologically similar among members, has been a taxonomic challenge based on host range, biologic behavior, and morphology. Genetic sequencing has added more complexity to this issue.
There is evidence of cophylogeny within specific hosts, but also considerable cross-species infectivity. Many Cryptosporidium spp., including C. cuniculus, have been traditionally named after the host species in which they were originally observed. Recent studies have shown that C. cuniculus has unique genetic sequences, and is now considered a validated species. It is most closely related to, but distinct from, C. hominis. Infections of rabbits are usually subclinical, and the organism is typically an incidental finding. When examined microscopically, the villi of the terminal small intestine may be shortened and blunted. Round to ovoid bodies are present within the brush border of enterocytes at the tips of villi. Changes on enterocytes are minimal and consist of elongation or shortening of microvilli adjacent to attachment sites. Recently, several outbreaks of cryptosporidiosis in humans have been associated with C. cuniculi, underscoring the zoonotic significance of this organism.Eimeria spp. Infections
Intestinal coccidiosis
Enteric coccidia are important causes of clinical (or subclinical) disease in commercial rabbitries, resulting in weight loss and mortality. There are 10 recognized species of intestinal Eimeria in the rabbit. Based upon experimental inoculation of pathogen-free laboratory rabbits, Eimeria spp. can be divided into 5 overlapping groups according to pathogenicity: nonpathogenic (£. coecicola), slightly pathogenic (£. perforans, E. exigua, and E. vejdovskyi), mildly pathogenic or pathogenic (£. media, £. magna, £. piriformis, and £. irresidua), and highly pathogenic (£.
intestinalis and £. flavescens). Each of these species targets specific segments of the intestinal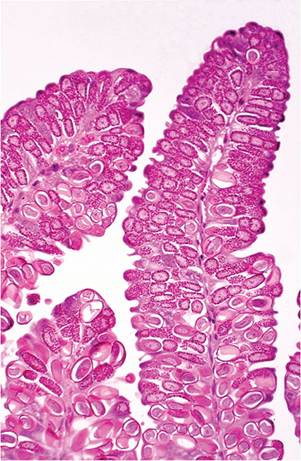
FIG. 6.65. Small intestine from a juvenile rabbit with acute coccidiosis. £nterocytes contain large numbers of micro- and macrogametocytes and oocysts.
tract (see Pakandl, 2009). Rabbits are usually coinfected with multiple Eimeria species. For example, all 10 species of intestinal Eimeria were found in young rabbits maintained at family farm rabbitries in Europe. As rabbits aged, E. flavescens and E. piriformis were predominant, and other species were rare. Disease severity depends upon infective dose, parasite species, immune status, and age of the rabbits. Enteritis is often accompanied by overgrowth of E. coli and presence of rotavirus, emphasizing the multifactorial nature of rabbit enteritis.
Epizootiology and Pathogenesis
Following passage in the feces, oocysts require 1 or more days to sporulate at room temperature before they become infective. Upon ingestion, sporulated oocysts (sporocysts) release sporozoites, which invade enterocytes and multiply by schizogony. In rabbits inoculated with sporocysts directly into the duodenum, excystation and the invasion of enterocytes occur as early as 10 minutes postinoculation. Sporozoites appear in the mucosa of the ileum within 6 hours, suggesting systemic rather than intraluminal migration. Depending on the species of Eimeria,1 or more asexual cycles occur, followed by gametogony and oocyst passage in the feces. The prepatent period is from 5 to 12 days, depending on the species. There may be a phenomenal number of progeny from a single ingested oocyst. One oocyst of E. magna may produce over 25,000,000 oocysts in a susceptible host.
Rabbits most frequently develop clinical disease during the postweaning period. Rabbits younger than 20 days of age are highly resistant to infection, and can only be infected with very high doses of oocysts.
This innate resistance is not fully understood, but has been attributed to factors in the milk, with age-related susceptibility evolving with changes in the intestinal environment attributed to consumption of plant feed. The most damaging stage in the life cycle is the sexual cycle, where there may be extensive destruction of enterocytes and cells in the lamina propria in affected sections of the gut. Because oocysts require sporulation at room temperature before they are infective, reingestion of the “night feces” (cecotrophy) does not play a role in the dissemination of the disease. Exposure to relatively small numbers of oocysts should result in a subclinical infection, with appropriate immune response. However, immunity to 1 species of Eimeria is unlikely to provide good protection against other species. In many commercial operations, the feed is routinely medicated with anticoccidials to control the disease. However, this should not be considered an acceptable substitute for rigid sanitation practices. In well-managed operations where anticocci- dials are not used, control is dependent on rigorous sanitation practices.Pathology
At necropsy, the perineal region and belly are frequently smeared with watery dark green to brown feces. The animal may be thin and dehydrated. Fecal losses of potassium result in hypokalemia. The cecum and colon contain dark green to brown watery, foul-smelling material. The mucosa of affected areas of the gut is congested and edematous, occasionally with hemorrhagic areas. Depending upon region of intestine involved, during acute coccidiosis, there is destruction of enterocytes, villus attenuation in affected areas of small intestine, denudation of cecal mucosa, and marked leukocytic infiltration of the lamina propria. With E. intestinalis, lesions are most severe at day 7-10 postinoculation with repair of the mucosa by day 12. Gametocytes and oocysts are usually evident in the intestinal mucosa in affected areas (Fig. 6.65).
The higher pathogenic nature of E. intestinalis and E. flavescens has been attributed to infection of crypt epithelium by these species, resulting in failure of enterocyte regeneration and bacterial invasion.Diagnosis
Fecal flotations, mucosal scrapings, and microscopic examination for oocysts are standard diagnostic procedures. An approximate oocyst count and oocyst speciation are recommended, particularly in view of the recognized variation in pathogenicity among species of Eimeria. A PCR assay has been developed for definitive identification of each of the 11 Eimeria species that infect domestic rabbits, including E. stiedae (below). In acute cases of coccidiosis, oocysts may not yet be present in the feces but will be evident in sections of the appropriate areas of small and large intestine. There is frequently a significant rise in the bacterial count, especially coli- forms. Quantitative aerobic cultures are useful indices of disease, since aerobic coliform growth is minimal in the normal rabbit intestine. Differential diagnoses include E. coli and Salmonella enteritis, Lawsonia enteritis, Tyzzer's disease, clostridial enteropathies, viral enteritides, and mucoid enteropathy. Multiple factors often cocontribute to rabbit enteritis.
Hepatic Coccidiosis
Eimeria stiedae infections occur in both domestic and wild rabbits and represent an important cause of poor weight gains, disease, and mortality in commercial rabbitries.
Epizootiology and Pathogenesis
Following the ingestion of sporulated oocysts (sporocysts), sporozoites invade the duodenal mucosa and migrate to the lamina propria prior to systemic migration. Sporozoites have been demonstrated in the regional mesenteric lymph nodes within 12 hours postexposure and in the liver by 48 hours. Organisms have been reported to migrate to the liver in mononuclear cells via lymphatics. However, viable sporozoites have also been demonstrated in the peripheral blood and bone marrow in E.
stiedae-inoculated rabbits, and the hematogenous route has been proposed as a means ofmigration to the liver. In the liver, sporozoites invade the epithelial cells of the bile ducts and schizogony begins. Following the gametogeny, oocysts are formed, released into the bile ducts, and passed to the intestine. The prepatent period is approximately 15-18 days. Oocysts may be shed in the feces for up to 7 or more weeks. Oocysts are normally resistant to environmental change; thus, contaminated premises and fomites may be a source of infective sporulated oocysts for several months. Eimeria stiedae infections may be manifest either as clinical or subclinical disease and typically accompany intestinal Eimeria infections.
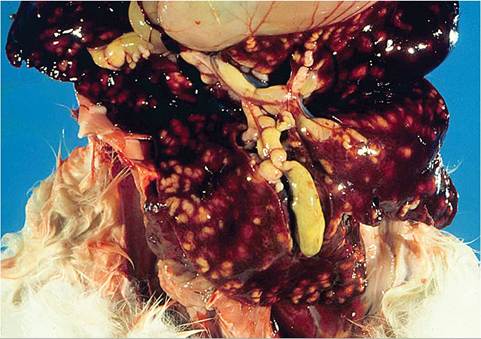
FIG. 6.66. Liver from a juvenile rabbit with florid hepatic coccidiosis. In addition to the raised linear hepatic lesions, representing involved bile ducts, the gall bladder and common bile duct are dilated and contain flocculent material.
Weanling rabbits are most often affected. In the past, a significant number of livers collected from fryer rabbits in abattoirs have been condemned because of hepatic coccidiosis. A dose-related effect has been observed in experimentally infected animals. In young rabbits inoculated orally with varying numbers of sporocysts (100-100,000 per animal), mortality rates in animals that received either 10,000 or 100,000 sporocysts were 40 or 80%, respectively. No fatalities occurred at lower dosages. Significant variations in liver enzymes and blood chemistry have been observed during the course of the disease. Four stages have been proposed: (i) the initial stage of metabolic dysfunction that coincides with liver damage during schizogony; (ii) the cholestatic stage, with elevated transaminases and serum bilirubin; (iii) the stage of metabolic dysfunction, characterized by hypoglycemia and hypoproteinemia; and (iv) the period of immunodepression in heavily infected animals resulting in an inability to curtail the production of oocysts in the biliary system.
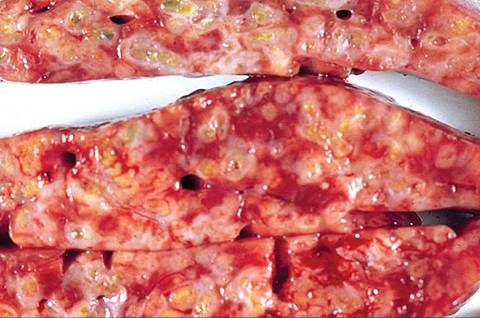
FIG. 6.67. Cut surface of the liver from a rabbit with hepatic coccidiosis. Bile ducts are dilated with mural thickening and lumina are filled with inspissated material.
Pathology
Affected animals are frequently thin, potbellied, and lack body fat reserves. There may be dark brown to green soiling in the perineal region. Ascites is a variable finding. Depending on the degree of liver involvement, there may be hepatomegaly, and in severe cases, icterus. In the liver, there are variable numbers of raised, linear bosselated, yellow to pearl gray circumscribed lesions 0.5-2 cm in diameter scattered throughout the hepatic parenchyma. The gall bladder is thickened and contains viscid green bile and debris (Fig. 6.66). On cut surface, lesions contain fluid green to inspissated, dark green to tan material (Fig. 6.67). Microscopically, there may be marked dilation of bile ducts, extensive periportal fibrosis, and mixed inflammatory cell infiltration in the periportal regions. In affected bile ducts, there is hyperplasia of epithelium, with papillary projections lined by reactive epithelial cells overlying collagenous tissue stroma. Infiltrating periductal inflammatory cells include lymphocytes, macrophages, and a sprinkling of polymorphonuclear leukocytes. Large numbers of gametocytes and oocysts may be present in parasitized ducts (Fig. 6.68). In lesions of some duration, organisms may be sparse to absent in bile ducts, with prominent periportal fibrosis.
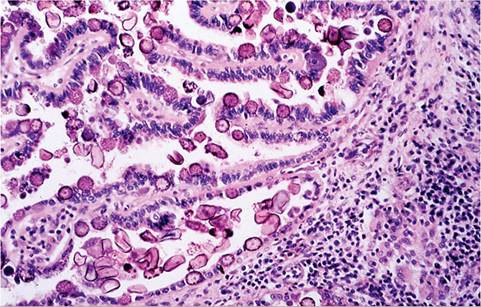
FIG. 6.68. Hepatic bile duct of a rabbit with chronic hepatic coccidiosis, featuring proliferative cholangitis, periportal fibrosis, and inflammation. Bile ductular epithelium contains large numbers of microgametocytes, macrogametocytes, and oocysts.
Diagnosis
The diagnosis can be confirmed at necropsy by wet mount preparations. Oocysts are usually readily observed in aspirates from the gall bladder or in impression smears of sectioned lesions. The characteristic proliferative biliary changes and organisms seen histologically are pathognomonic of the disease.