Developing Technologies: Clustered Regularly Interspaced Short Palindromic Repeats Interference
Clustered regularly interspaced short palindromic repeats interference (CRISPRi) is a modification of the original CRISPR∕Cas9 (CRISPR-associated protein 9) system, designed with a goal of gene silencing instead of knockouts.
For this approach, the Cas9 gene is modified to render its product catalytically dead (dCas9), eliminating endonuclease activity (Qi et al., 2013; Adli, 2018). Lacking nuclease activity, the complex otherwise behaves identically to the wild-type. The short guiding RNA (sgRNA) directs the dCas9 to the complementary DNA and pairs with it, placing dCas9 over the target strand, where it remains unable to cleave. The target site is then silenced through simple physical hindrance, with dCas9 remaining bound so long as conditions permit sgRNA∕DNA pairing. To date, there are two primary dCas9 CRISPRi systems developed for Mycobacterium: codon-optimized from Streptococcus pyogenes (Choudhary et al., 2015; Singh et al., 2016) and one from Streptococcus thermophilus (Rock et al., 2017). While both systems share the advantage of being anhydrotetracycline (ATc) inducible, fully reversible at any stage and minimize toxicity from protein overexpression, the more traditional S. pyogenes dCas9 shows less robust and specific gene knockdowns than the S. thermophilus dCas9.This section will focus on the Rock et al. protocol (Rock et al., 2017). When using the CRISPRi system, the desired repression strength is initially selected through a list of a protospacer adjacent motifs (PAMs) with repression levels for a gene target ranging from 2.7- to 216.7-fold. The broad library of PAMs identified allows flexible target selection across mycobacterial genomes while minimizing the difficulties and risks with gene knockouts. The demonstration of efficacy in both saprophytic M. smegmatis and pathogenic M. tuberculosis establishes its function across dissimilar mycobacteria.
As shown in Fig. 7.2, to disrupt transcription initiation through the promoter region, either the template (T) or non-template (NT) strand can be targeted. However, when silencing through the ORF or 5'-UTR, part of the sgRNA must pair with the NT strand. Here, we will assume a PAM is selected within the ORF. For the designed sgRNA to pair with the NT strand, a PAM is sought in the T strand. After the preferred seven nucleotide (nt) PAMs are identified, the complementary sgRNA sequence itself is pulled from the 20-25 nt directly upstream. The 5' end of this sequence should end in a G or A, so if the 20th nucleotide is a C or T, one additional base can be moved to the 5' direction as necessary. In this scenario, the guiding portion of the sgRNA would be identical to the template strand sequence immediately at the 5' end of but not including the PAM sequence. This 20-25 nt sequence serves as the forward primer, and the reverse complement as its reverse. 5'-GGGA-3' is added at the 5' end of the forward primer, and 5'-CAAA-3' added to the 5' end of the reverse primer to allow cloning into the BsmBI-digested vector and ligation into the dCas9 handle sequence therein. This allows the chosen pairing sequence to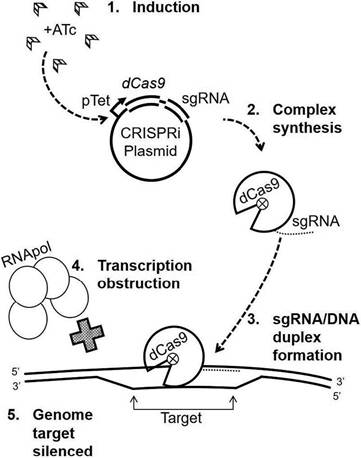
Fig. 7.2. Schematic steps in the clustered regularly interspaced short palindromic repeats interference (CRISPRi) system developed for Mycobacterium from Streptococcus thermophilus dCas9. (1) anhydrotetracycline (ATc) attaches to the tetracycline (Tet) repressor regulated optimized promoter (pTet) to induce expression of dCas9 from CRISPRi plasmid with a mycobacterial integrating vector (Table 7.1) as a backbone; (2) formation of the dCas9 complex; (3) dCas9 complex is directed to the target DNA sequence; (4) duplex formation prevents transcription initiation (RNA polymerase) or elongation (steric hindrance); (5) the gene is silenced (downregulated).
be integrated into the dCas9 protein as a full sgRNA.
Transformants are selected by kana- mycin resistance, and activation of the system occurs by addition of ATc. Overexpression of dCas9 is known to show toxicity to the bacterial host (Rock et al., 2017). As such, Rock et al. designed two distinct plasmids for mycobacterial use - pLJR962 (optimized for M. smegmatis) and pLJR965 (optimized for M. tuberculosis). Each vector responds differently to ATc pools in the bacterial host, leading to differential induction and system expression. Our Michigan State Laboratory has successfully cloned pLJR965 into MAP. However, ATc handling in the MAP system and therefore the optimum vector has not yet been determined. Inducible and tunable gene silencing in MAP without genome modification offers promise for studying gene function and pathways in this stubborn pathogen, as its slow-growing and fastidious nature have made traditional gene knockouts comparatively difficult.By adopting the CRISPRi system, researchers can explore the classical reductionist analysis on the effects of a single gene silenced on a host, to the broader systems approach studying the interplay of one silenced gene between pathways and the overall bacterial lifecycle. This approach still maintains the original genomic context and with minimal upstream and off-target effects. Additionally, the system's inducibility and tunable repression allows a much more nuanced view of biological effects beyond the on/off binary. For essential genes where knockouts were previously impossible, CRISPRi allows investigation by either inducing only in permissible conditions or growth phases, or by graded knockdown rather than outright depletion. The CRISPRi knockdown system offers several advantages as discussed here, but its simplicity and low cost combined with its potential for comparatively rapid implementation make this technique very attractive, especially for researchers working with MAP.
7.7