Endocrine pancreas
The two primary endocrine products of the pancreas are insulin and glucagon. Both hormones are simple proteins that synthesized in small clusters or islands of cells nested within the exocrine tissue of the pancreas.
The German pathologist Paul Langerhans discovered these clusters of cells, now called the islets of Langerhans, in 1869. They represent only a small fraction of the mass of the pancreas (1-2%). Each individual islet (~0.4mm in diameter) contains only a few thousand cells but they are nonetheless critical for homeostasis. Four cell types can be distinguished within the islets. Glucagon-producing a cells are about 20% of the total. Insulin-secreting β cells are the most abundant, constituting 60% to 80% of the total. Somatostatinproducing δ cells are infrequently observed, 3-10% of the total, but pancreatic polypeptide secreting PP- or F-cells are rare with ~1% abundance.Insulin is a small protein, with a molecular weight of about 6000 daltons. It is composed of two chains held together by disulfide bonds but it is synthesized as a precursor molecule in which a fragment called the C peptide is removed to produce the biologically active molecule. The amino acid sequence of insulin is highly conserved. For example, it is still common for diabetic patients to be treated with porcine insulin. In fact, the first case of treatment of a human diabetic with insulin was in 1922.
Insulin synthesis begins with translation of its mRNA as a single chain precursor called preproinsu- lin. As it passes into the cisternal space of the RER, removal of its signal peptide produces proinsulin. Proinsulin has three domains: an amino-terminal B chain, the A chain carboxy-terminal region, and a connecting peptide in the middle called the C peptide. Once it passes into the cisternal space of the RER, endopeptidases excise the C peptide to produce mature insulin.
Insulin and free C peptide are packaged in the Golgi into secretory granules, which accumulate in the cytoplasm. When the β cell is stimulated, insulin and C peptide are secreted from the cell by exocytosis. However, C peptide has no known biological activity.Secretion of both insulin and glucagon is closely tied to circulating concentrations of glucose. For optimal health and maintenance of homeostasis, blood glucose concentrations must be maintained within fairly narrow boundaries. In the case of the β cells, extracellular glucose is transported into the cells by facilitated diffusion involving specific glucose transporter proteins. Since its uptake depends on diffusion, the greater the extracellular glucose concentration, the greater the corresponding intracellular concentration. As concentrations reach a threshold value, this produces a change in membrane depolarization and subsequently an influx of extracellular calcium. The precise mechanism(s) responsible for depolarization are not well defined but may depend on metabolism of glucose that is taken up into the cell and alterations in the ATP:ADP ratio within the cytoplasm. Regardless, the increase in free calcium is believed to be a primary stimulator of exocytosis of insulin-containing vesicles.
It is clear that increased glucose concentrations markedly increase secretion of insulin in normal animals. This is particularly evident in monogastric animals after feeding of a high-carbohydrate meal. Figure 12.35 illustrates the increase in insulin in cows that were given a bolus injection of glucose into the
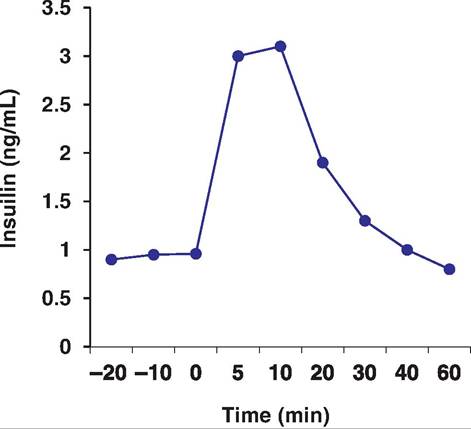
Fig. 12.35. I nsιιlin after glucose. Cows were infused with glucose (0.1 g/kg body wt.) at time zero. Data adapted from Denbow et al. (1986).
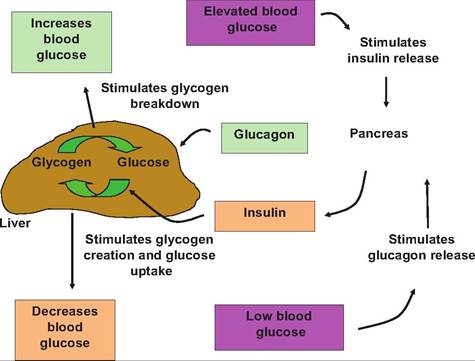
Fig. 12.36. Regulation of blood glucose. When glucose is elevated, the pancreas releases insulin.
This promotes the uptake ofglucose and glycogen formation in the liver, thus reducing blood glucose-promoting hypoglycemia. Glucagon is released when blood glucose is low. This stimulates glycogen breakdown, thereby returning glucose concentration to normal, a hyperglycemic response.jugular vein. This experiment was done to compare pancreatic responsiveness among cows in various seasons. There is, plainly, an abrupt increase in serum insulin following a glucose challenge even in ruminants.
Although secretion of insulin acts to lower blood glucose by promoting the uptake of glucose into most cells and particularly increased glycogenesis in liver tissue, control of blood glucose concentration does not simply depend on changing secretion of insulin. When blood glucose concentration drops, the islets secrete glucagon. Glucagon stimulates the breakdown of glycogen and promotes gluconeogenesis. As a generalization, increases in blood insulin concentrations act to drive energy substrates into storage in tissues by increasing accumulation of glycogen, which can be mobilized when needed. This association is illustrated in Figure 12.36.
Problems with regulation of blood glucose concentrations are relatively common in humans and animals. Naturally occurring diabetes is probably the most common endocrine disorder diagnosed in dogs, with estimates for the frequency of diabetes ranging from a high of 1 in 100 to 1 in 500. Diabetes mellitus is a complex disease that is manifested by inappropriate hyperglycemia. Symptoms and problems result from of a failure of insulin secretion, failure of biological action of insulin, or both. It is useful but somewhat arbitrary to classify diabetes as either insulin -dependent (type I) or noninsulin dependent (type II). In type I diabetes, insulin is very low or absent, plasma glucagon typically elevated, and the β cells fail to respond to stimuli known to stimulate insulin secretion in healthy animals. Typical symptoms include polyuria and polydipsia, as well as weakness and fatigue and often polyphagia with weight loss.
The increased urination is a consequence of osmotic diuresis related to hyperglycemia. Thirst and polydipsia occur as a consequence of hyperosmotic state of body fluids. Since glucose is poorly utilized in the absence of insulin, ketosis and subsequently ketoacidosis occur as fatty acids and are Catabolized to supply energy demands. Wasting of muscle mass can also happen as muscle proteins are degraded to supply amino acids for glucose and ketone bodies. Ironically, it is as if the body is starving, despite the presence of large quantities of blood glucose. Without insulin, the capacity of cells to capture blood glucose is impaired, yet energy demands remain. To add insult to injury, so to speak, the β cells of the islets also require insulin to sense that glucose concentrations are adequate. This means that a cells are stimulated to secrete glucagon; this only worsens the problems by promoting glycogenolysis and gluconeogenesis. With acute insulin deficiency, the increased energy demands, and failure to utilize glucose, ketones (acetoacetate, β-hydroxybutyrate, and acetone) rapidly increase. Along with reduced renal blood flow (reduced extracellular fluid volume), reduced renal excretion of hydrogen ions also can occur. This can lead to dramatic ketoacidosis. Continuing increases in blood osmolarity (>330mOsm∕L) and progressive acidosis produce coma and ultimately cardiovascular collapse and death.Clearly, the primary treatment option is to monitor blood glucose to minimize drastic swings with careful attention to diet and replacement of missing insulin with exogenous treatments. Fortunately, recent advancements in molecular biology and biotechnology have made multiple options available. Many patients still are treated with porcine or bovine insulin but recombinant human insulin is also available. There are also multiple forms of insulin available for therapy, that is, short acting (so called regular insulin), as well as intermediate and long-acting formulations.
This has increased the opportunity to better control blood glucose, particularly with respect to meals.With fewer acute episodes but chronic hyperglycemia, diabetes can lead to a variety of pathological changes. Vascular diseases include both microvas- cular and macro vascular problems. In capillaries and smaller precapillary arterioles there is frequently a thickening of the basement membrane. These can be especially problematic in the retina, leading to retinopathy and progressively failed vision, and in the kidney, progressive disease produces pathological problems in the peritubular capillary beds surrounding the nephrons. In larger vessels, diabetes is believed to accelerate atherosclerosis and all of the associated problems, that is, it increases incidence of myocardial infarction, stroke, and peripheral gangrene, which seems to be largely unique to diabetic patients.
Diabetes is also associated with increased incidence of cataracts. Increased blood glucose is believed to contribute to formation of cataracts by promoting glycosylation of lens proteins and by increasing concentrations of sorbitol within the lens tissue. Sorbitol is a by-product of glucose metabolism. As it accumulates this promotes osmotic changes that ultimately promote swelling and fibrosis. Changes in the cornea and lens are frequently rapid and are a consistent ocular manifestation of diabetes. A decrease in corneal sensitivity is often observed as the disease progresses. Corneal nerves are important in producing protective responses (tear production, eyelid closure) so that diabetic-related impairment of these responses can have important consequences. Table 12.10 shows the results of Cochet-Bonnet aesthesiometer measurement to compare the sensitivity of the corneal touch threshold (CTT) in diabetic compared with normal dogs.
Table 12.10. Corneal touch threshold in diabetic and normal dogs.
| Region of Cornea | Diabetic | Normal |
| Central | 2.9 | 1.8 |
| Nasal | 4.0 | 2.2 |
| Dorsal | bgcolor=white>5.12.8 | |
| Temporal | 5.1 | 2.8 |
| Ventral | 6.5 | 5.1 |
Data adapted from Good et al.
(2003).This test utilizes an instrument that has 6.0 cm length of monofilament nylon that is pushed against the cornea until the there is a slight deflection. Reducing the length of the filament in 0.5 cm segments until a blink reflex is recorded continues the process. This length is recorded and converted to a force value (g∕ mm2), in other words, the force required to elicit the blink response (from 0.4 to 15.9g∕mm2) depending on the length from 6.0 to 0.5 cm. Testing was done to compare various regions of the cornea, that is, central (C), nasal (N), Dorsal (D), Temporal (T), and Ventral (V), and averaged for both eyes. These data show a consistent pattern of reduced sensitivity (i.e., greater force required to elicit the blink response) in diabetic compared with clinically normal dogs.
In type II diabetes, the problem is failure of insulin to be biologically active. However, this is typically a milder form of the disease in that it is heterogeneous with respect to symptoms and the degree of biological failure. This also influences treatment options. Sim- plistically, type I diabetes treatment requires replacement of missing insulin. The situation is more complex with type II because treatment options depend on the relative contribution of still-functional β cells and the degree of insulin insensitivity. For example, type II is more common in obese animals. This in part reflects the fact that adipose tissue has abundant insulin receptors and is a key tissue in control of energy balance and homeostatic alterations in carbohydrate metabolism. Therefore, a common treatment scheme is to promote weight loss and to control episodes of hypo- or hyperglycemia with diet.