Adrenal gland
Despite their small size, the adrenal glands, located at the superior pole of each kidney, are critical regulators of metabolism. The outer portion of each gland, the cortex, is responsible for the production of two broad classes of steroid hormones: the mineralocorticoids, of which aldosterone is a prime example, and glucocorticoids, represented by cortisol.
The center of the gland, the medulla, is derived from neural tissue. It is essentially postganglionic tissue that is part of the sympathetic division of the autonomic nervous system. When stimulated it secretes epinephrine, a structural cousin of the neurotransmitter norepinephrine. Both of these regions are important in adaptations necessary to respond to stress and to maintain homeostasis. When animals are under prolonged stress it is not uncommon that the adrenal glands become enlarged. However, under extreme situations the capacity to respond can be lost, resulting in exhaustion. In the 1930s the work of Sir Hans Selye was among the first to focus on the role of the adrenal to combat stress. He studied the response of adrenalectomized animals to injury or stress and pioneered what came to be called the general adaptation syndrome. His hypothesis consisted of three phases: (1) alarm reaction, (2) stage of resistance, and (3) stage of exhaustion. This can be envisioned by considering the pathways responsible for glucocorticoid secretion and biological responses to glucocorticoid release. First, stress induces neural stimulation, leading to hypothalamic secretion of CRH. This results in secretion of ACTH, which produces glucocorticoid secretion. An early effect of glucocorticoid release is mobilization of glycogen reserves to increase circulating glucose. Other tissues are progressively Catabolized to provide fatty acids or amino acids for energy production. If these actions provide the necessary nutrients to respond to the stress alarm, repairs are made and conditions return to normal. When stress continues but is manageable, a new "set point" or stage of resistance is achieved. However, with more extreme or prolonged stress, a new balance cannot be achieved so that signals for more glucocorticoid release cannot be answered by the adrenal cortex. This is the stage of exhaustion.The adrenal cortex is organized into three zones. Just below the protective capsule that surrounds the gland, is the zone glomerulosa, which typically accounts for about 20% of the cortical cells. Here the
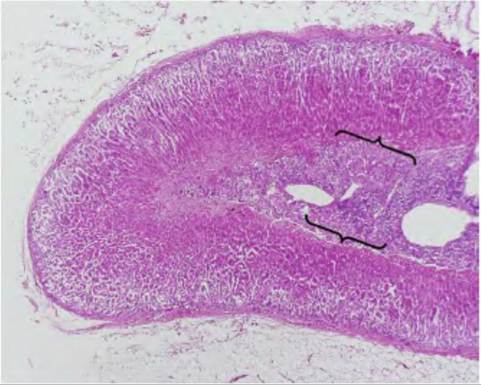
Fig. 12.32. Adrenal gland. Note the central area (medulla, brackets) and the outer cortex; H&E staining, 2? magnification.
cells are organized into more or less spherical clusters, hence the name glomerulus, Latin for "little ball." The zona fasciculata is the largest region and the cells are typically arranged into columns. Between the columns there are long fenestrated capillaries. The innermost region of the cortex, the zona reticularis, is about the size of the zone glomerulosa and the cells are arranged in an irregular network (Fig. 12.32 and Fig. 12.33).
The major hormones made in the adrenal cortex are cortisol or corticosterone (typical glucocorticoids), androgens (testosterone or testosterone-like), and aldosterone (mineralocorticoids). The zone glomerulosa is primarily responsible for the synthesis of aldosterone. Although we discussed some of the basics of steroid hormone structure earlier, some review is in order. Steroid synthesis begins with cholesterol. Much of the needed cholesterol comes from uptake from plasma lipoproteins but synthesis directly from acetate also occurs. A critical, rate-limiting step involves the conversion of cholesterol to pregnenolone. In the two inner zones of the cortex, this is stimulated by ACTH. This conversion occurs within the mitochondria and involves two hydroxylation steps along with cleavage of the side chain of the cholesterol.
The pregnenolone passes out of the mitochondria for further processing. Differences in steroid synthesis between zones of the adrenal cortex reflect differences in the enzymes present. The greatest differences are between the zona glomerulosa and the inner zones. This explains why aldosterone is the predominant product of the glomerulosa. Specifically, the zona glomerulosa lacks 17α-hydroxylase activity so cannot produce either 17oc-hydroXypregnenolone or 17α-hydroxypro- gesterone. This is relevant because these molecules are the immediate precursors for cortisol and adrenal 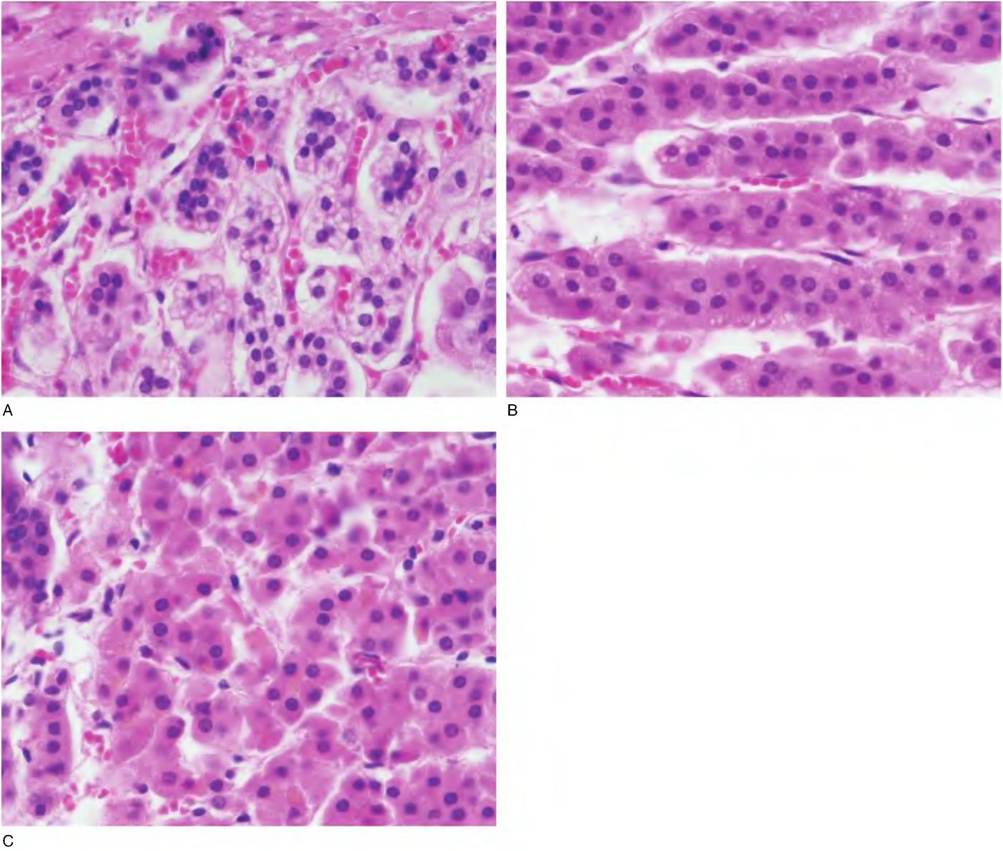
Fig. 12.33. Areas of the adrenal cortex. A = glomerulosa; B = fasciculata; C = reticularis.
androgens. Furthermore, only the zona glomerulosa can convert corticosterone to 18-hyHroxycortisosterone and aldosterone. Synthesis of aldosterone is largely controlled by the renin-angiotensin system of the kidney. This will be discussed in greater detail in our study of the urinary system. However, the essential point is that decreases in renal blood pressure lead to the production of angiotensin II, which, among other actions, stimulates the secretion of aldosterone. Aldosterone then acts to increase resorption of sodium by the distal convoluted epithelial cells of the kidney nephrons. Increased recovery of sodium from the urinary filtrate allows the companion recovery of more water (via osmosis) so that interstitial fluid volume and subsequently blood volume is increased. This returns blood pressure to normal, thus shutting off the trigger for increased aldosterone secretion in the first place. Thus, secretions and regulation of the zona glomerulosa and two inner zones of the adrenal cortex are quite distinct. Some of the pathways for synthesis of various steroids in the adrenal gland are illustrated in Figure 12.34.
ACTH rapidly stimulates secretion of glucocorticoids from the inner zones of the adrenal cortex, along with increases in DNA, RNA, and protein synthesis.
With chronic stimulation there is hyperplasia and hypertrophy of the cortical cells. Binding of ACTH to cortical cell receptors activates adenylyl cyclase, leading to increased concentrations of cAMP. This activates intracellular phosphoprotein kinases. This ultimately stimulates the rate-limiting conversion of cholesterol to pregnenolone to allow for steroidogenesis. ACTH secretion is controlled by secretion of CRH from the hypothalamus so that changes in ACTH are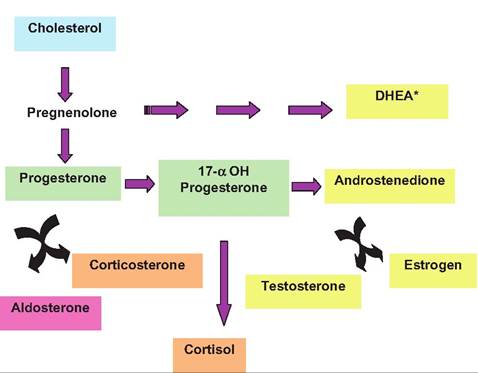
Fig. 12.34. Adrenal steroid biosynthesis. Cholesterol is essential but conversion to pregnenolone allows subsequent production of sex steroids (yellow), glucocorticoids (tan), and miIteraldocorticoids (pink). *DHEA is dehydroepiandrosterone sulfate.
closely paralleled by changes in cortisol or corticosterone secretion (depending on the species). However, neuroendocrine control of adrenal cortical activity is complex. Three levels of regulation include: (1) circadian rhythm, (2) stress response, and (3) feedback inhibition. A circadian rhythm is superimposed on the typical episodic secretion of glucocorticoids that occurs throughout the day. This reflects CNS activity that controls both the number and magnitude of CRH and ACTH secretory episodes during the day. In primates, glucocorticoid secretion is usually low in the late evening and with the onset of sleep, but as dawn or wakefulness begins, CNS activity and corresponding CRH and ACTH secretory events increase in number and magnitude so that glucocorticoid concentrations increase. This pattern of low activity during sleep and increased activity during early day is relatively fixed, but substantial relative variability occurs. This inherent rhythm is impacted by physical and mental stresses and various disease conditions. Stress responses are usually tied to corresponding increases in release of epinephrine and thyroid hormones. This family of reactions allows for metabolic effects to maintain homeostasis.
For example, increased glycogenolysis and gluconeogenesis provide fuel to synthesize ATP in stressful fight-or-flight situations. In conjunction with secretion of epinephrine from the adrenal medulla, a major effect in adipose tissue is increased lipolysis and release of glycerol and fatty acids for energy production.Glucocorticoids impact multiple tissues. In excess they inhibit collagen synthesis by fibroblasts. This can produce thinning of the skin, easy bruising, and slow healing of wounds. Impaired bone formation can occur from reduced cell proliferation and extracellular matrix protein synthesis required for deposition of in inorganic components of bone. This can clearly impact mineral metabolism as well as growth and development. One of the effects of glucocorticoids, inhibition of neutrophils, is used therapeutically to lessen inflammation. These potent effects demonstrate the dramatic problems that accompany situations where there is either too much or two little of the adrenal steroids are produced.
Routine control over glucocorticoid secretion depends on negative feedback at multiple levels (see Fig. 10.13). In particular, increased concentrations of glucocorticoids inhibit secretion of CRH from the hypothalamus as well as ACTH from the pituitary. In fact, two distinct negative feedback responses have been described. A fast feedback, glucocorticoid-induced inhibition of ACTH secretion, is directly related to the rate of increase in glucocorticoid secretion. This phase occurs within minutes and acts to reduce both basal and ACTH- Stimulated increases in adrenal glucocorticoids. A delayed feedback inhibition subsequently suppresses both CRH and ACTH secretion. At the extreme, prolonged administration of glucocorticoids suppresses CRH and ACTH secretion and the adrenal cortex atrophies. Addison's disease or adrenocortical insufficiency occurs when there is a failure of the adrenal cortex to produce sufficient amounts of mineralocorticoids or glucocorticoids.
The consequences of this are clearly dramatic. Symptoms can include muscle weakness, fatigue, anorexia, hypotension, hyponatremia, and hypoglycemia. With companion failure of aldosterone production, additional symptoms include dehydration hyperkalemia and acidosis. Causes include destruction or dysfunction of the adrenal cortex. For example, tuberculosis can destroy the adrenal cortex, as can autoimmune disorders. Secondary causes reflect failures at the level of the hypothalamus (impaired secretion of CRH) or pituitary (impaired secretion of ACTH). Before adrenal steroids became available for therapy, primary adrenocortical insufficiency was fatal.The opposite problem, chronic secretion of excess glucocorticoids, leads to an array of problems called Cushing's syndrome in humans. The most common cause is excessive glucocorticoid therapy. This again emphasizes the importance of careful use of these potent steroids. Classic Cushing's syndrome is often caused by pituitary or adrenal abnormalities that lead to excess concentrations of ACTH, that is, tumor products. Cushing's disease has multiple effects on tissue metabolism and organ function. This is expected given the widespread distribution of glucocorticoid receptors. Obesity can occur with alterations in distribution of adipose tissue; that is, moon face and appearance of a "buffalo-hump" is a symptom that can develop over time. Hypertension, glucose intolerance, and gonad dysfunction are also manifestations of the disease.
As concerns with animal health and welfare and perceived problems attributed to stress have emerged in recent years, tools to quantitatively measure stress have been sought. Certainly there are behavior attributes and production-related measures (absence of chronic disease, rate of gain, milk production, etc.) that can be linked with disruption of homeostasis tied to stress, but many of these are poorly defined. It is also true that some level of stress is necessary, even desirable, for normal physiological responses and health. As indicated earlier, during short-term stress, secretion of glucocorticoids and epinephrine allow mobilization of nutrients necessary for homeostasis. The problems arise with severe chronic stress and consequences of prolonged secretion of glucocorticoids, that is, immunosuppression and atrophy of tissues. Regardless, measuring changes in circulating concentrations of glucocorticoids, responses to an ACTH challenge, and/or secretion of epinephrine provide a generally accepted quantifiable stress index for animals. Paradoxically, it is possible that the process (handling, needle sticks, and restraint) of taking frequent blood samples (often necessary because of the episodic nature of glucocorticoid secretion) can be stressful in itself. This has led to development of remote blood sampling devices or sampling of other body fluids (saliva or urine), but these samples may also require confinement or handling that can confound results. Mostl and Palme (2002) described the assay of metabolites of cortisol in feces as a tool to noninvasively monitor secretion of glucocorticoids as a possible stress index. The concentrations of these cortisol metabolites in feces reflect a kind of "average" glucocorticoid production over a period of hours that is likely species specific. For example, in sheep and cattle changes in fecal concentrations of 11,17-dioxoandrostane are correlated with changes in blood levels of cortisol following treatment with a bolus challenge of ACTH but with a 10- to 12-hour delay (Palme et al., 1999).