Thyroid gland
Located just below the larynx, the thyroid has left and right lobes medial to the midline of the ventral surface of the trachea. A sliver of tissue called the isthmus connects the lobes.
The thyroid gland is composed of clusters of follicles whose internal surfaces are lined by a layer of simple cuboidal epithelial cells. T3 and T4 are synthesized by the follicular cells and stored as part of a larger protein (thyroglobulin) within the Iumenal spaces of the follicles. This stored material is called colloid. Figure 12.23 shows the histological structure of a bovine thyroid gland. The follicular cells predominate but in the spaces between follicles parafollicular or C cells occur. The C cells synthesize CT, which is involved in calcium metabolism (Fig. 12.26).Biosynthesis of triiodothyronine and thyroxine
The critical elements required for thyroid hormone synthesis are the amino acid tyrosine and iodine. As iodine is absorbed into the follicular cells, it becomes attached to the ring structure of tyrosine amino acids that are part of the thyroglobulin sequence of amino acids. Each tyrosine is capable of becoming iodinated with two iodide atoms. When this occurs, the result is diiodotyrosine, but attachment of a single atom produces monoiodo tyrosine. The subsequent coupling of two diiodotyrosines produces T4. The coupling of one diiodo tyrosine and one monoiodotyrosine yields T3. When the intestinal cells absorb iodine it is converted to iodide for use in the thyroid. Thyroglobulin is a very large glycoprotein (5496 amino acids) that has about 40 tyrosine residues. The follicular cells have an active transport system that efficiently sequesters iodide so that intracellular concentrations can be 200- fold greater that in circulation. Structures of thyroid hormones are shown in Figure 12.27.
Pathways for the synthesis, storage, and release of thyroid hormone are illustrated in Figure 12.28. The process can be divided into six steps: (1) uptake of I, (2) oxidation of iodide and iodination of tyrosine residues in the thyroglobulin, (3) linking of the DIT and MIT to create T3 and T4, (4) proteolysis of thyroglobulin to produce free T3 and T4, (5) deiodination of iodotyrosines in the follicular cells for reuse, and (6) 5'-deoiodination of T4 to generate T3.
Biological effects of thyroid hormones
Thyroid hormones are the major regulators of basal metabolism. They increase oxygen consumption and therefore heat production because they stimulate oxidative phosphorylation. This is called a Calorigenic effect, a response especially useful when animals are exposed to cold stressful situations. Of course increasing energy demands require nutrient fuel. Small amounts of thyroid hormones promote glycogen storage but glycogenolysis is stimulated as concentrations rise. Other effects include increased absorption of glucose and promotion of glucose uptake by cells. Thyroid hormones also impact lipid metabolism, espe-
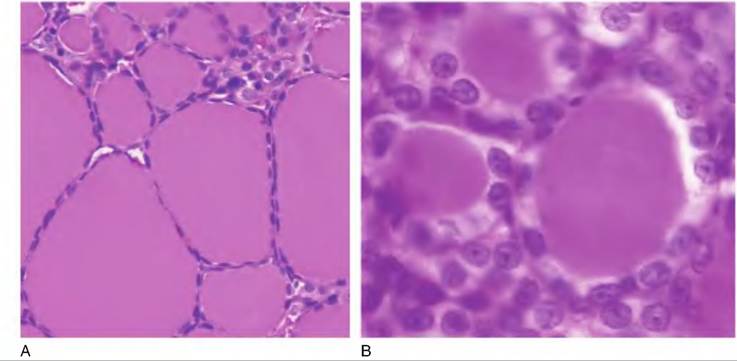
Fig. 12.26. Thyroid histology. The left panel is a low-power (20?) view of a sheep thyroid. The follicles are filled with colloid and the epithelial cells are compressed. The left image (40?) is the thyroid from a cat. Here the cells are more cuboidal and are actively reabsorbing colloid to make T3 and T4 available.
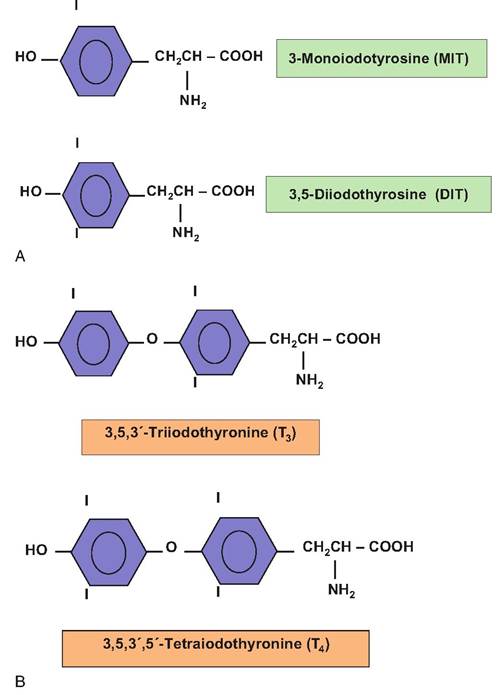
Fig. 12.27. Structures of thyroid hormones.
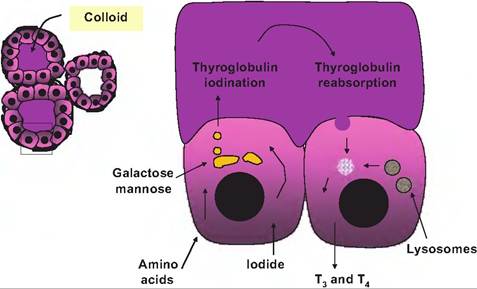
Fig. 12.28. Iodination of thyroid hormone. A cluster of follicles is shown on the left and on the right an enlargement of two cells.
In the cell on the left, amino acids allow synthesis of the thyroglobulin followed by posttranslational modification and packaging in the Golgi. Once thyroglobulin is secreted, iodination is completed. Secretion of TSH promotes the resorption of the iodinated thyroglobulin, fusion with lysosomes, and ultimately releases of T3 and T4.cially lipolysis. Fitting their ability to increase metabolism, the thyroid hormones also enhance the effects of sympathetic nervous system stimulation. This is linked to thyroid hormone stimulation of synthesis of β-adrenergic receptors in tissues that are targets for epinephrine and norepinephrine. This mechanism also likely explains the enhanced force of cardiac contractions when thyroid hormones are elevated.
Other effects of thyroid hormones are evident during growth and development. For example, classic experiments showed that T4 causes differentiation of tadpoles into frogs. Thyroidectomy or treatment with antithyroid drugs caused the animals to grow into very big tadpoles but metamorphosis was blocked. The situation is less drastic in mammals but thyroid hormones are nonetheless essential for normal development of the nervous system.
Given the myriad of effects attributed to thyroid hormones, it should not be a surprise that deviations from a normal or euthyroid state impact physiology generally and homeostasis in particular. Hypothyroidism, or a deficiency, slows metabolic processes. In young animals development and growth is impaired and in primates serious permanent failure of neural development produces mental retardation, leading to cretinism. Hypothyroidism in mature animals produces lethargy. For example in humans, deposition of glycosaminoglycans in the skin causes puffiness and clinical symptoms called myxedema. Causes of hypothyroidism vary but can be classified as (1) primary (thyroid failure), (2) secondary (pituitary TSH problem), (3) tertiary (hypothalamic defect), or (4) quaternary (peripheral resistance to action of the hormones).
This last case is frequently related to autoimmune disease in which generation of antibodies against thyroid cells, thyroid hormone receptors, or thyroglobulin impairs function. Among domestic species, lactating dairy cows are typically hypothyroid so that peripheral deiodination of T4 to produce the more potent T3 is especially important to maintain function of many target tissues. Hypothyroidism is also common in dogs. The cause is not well understood but detection of antibodies against thyroglobulin in many of these animals suggests autoimmune disease plays a role. Hyperthyroidism is relatively common in older cats and is usually associated with appearance of benign thyroid tumors.Fortunately, because of the common structure of thyroid hormones across species, RIA procedures originally developed for human testing are appropriate for animal testing. A common functional test is to measure concentrations of plasma T3 for T4 in response to injections of TSH or TRH. In correspondence with variations in metabolic rate with season, the data in Figure 12.29 illustrate secretion of T3 and T4 following TRH injections in cows during winter compared with
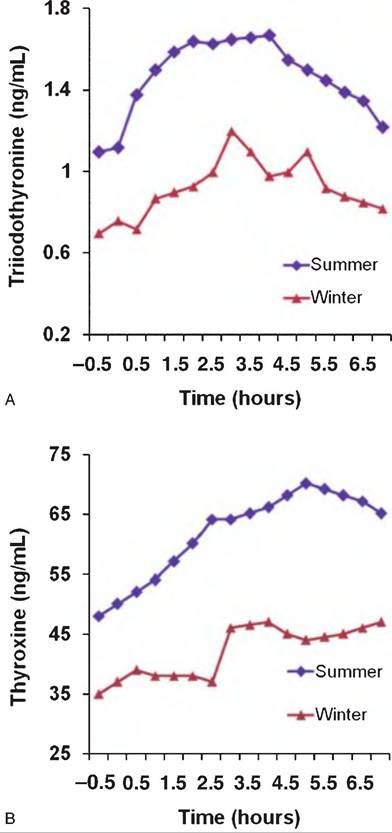
Fig. 12.29. Thyroid hormone responses to TRH. Lactating cows were injected with TRH (25μg∕1OOkg body wt.; arrow) and blood was collected to monitor changes in T3 and T4. Secretion of both hormones was reduced in winter. Data adapted from Perera et al. (1985).
summer. After a delay, TRH produced modest increases in circulating T3 and T4 in lactating cows. This reflects the time delay required for TRH to first stimulate the secretion of TSH by the anterior pituitary gland and for TSH to induce the process of colloid reabsorption by the thyroid follicular cells. There is also an evident seasonal variation in the response to TRH in these cows.
Mean concentrations of T3 and T4 before administration of TRH were lower in cows sampled during winter and response to TRH was reduced. These reductions likely reflect greater utilization of thyroid hormones to enhance thermogenesis during the winter. Concentrations of T4 were about 50-fold greater than concentrations of T3. This is similar to other domestic animals and suggests that substantial T3 for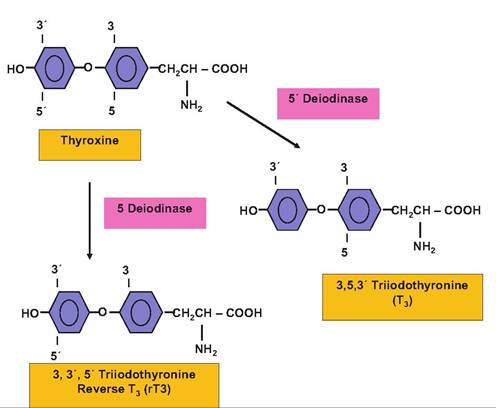
Fig. 12.30. Structure and nomenclature for conversion of thyroxine.
mation occurs outside of the thyroid gland by the deiodination of T4. Tissues with high levels of deiodination enzymes include the liver and kidneys. The mammary gland also expresses a deiodinase that increases with the onset of lactation and in response to other hormones known to stimulate milk production. This provides an enhanced local tissue concentration of mammary T3 available to stimulate metabolic activity to support high levels of milk production despite the fact that lactating cows are typically in a hypothyroid state.
Thyroid hormones do not circulate freely in the serum but are bound to plasma proteins. The most predominant is T4-binding globulin, but its concentration is relatively low so that despite its high- binding affinity, substantial amounts of T3 and T4 also circulate bound to albumin. A third protein, T4-binding pre-albumin, is especially good at sequestering T4. Although only free thyroid hormones are able to pass into target cells to bind with nuclear T3 receptors, the hormones have the potential for long lasting effects, in part because of their very long half-life. Whereas most hormones are rapidly degraded or taken from circulation (half-lives of minutes), T3 and T4 have long half-lives of about 1 and 7 days, respectively.
Since T3 is the essential biologically active thyroid hormone, deiodinase reactions to convert T4 are physiologically important. It is reasonable to view the relatively larger quantities of circulating T4 as a local tissue source for T3 in target cells. Two primary deiodination pathways act to convert T4 to either biologically potent T3 (5'deio- dinase enzyme) or to reverse T3 (5-deiodinase enzyme), which is biologically inert. Thus, the increased 5- deiodinase can be viewed as a degradation pathway or increased 5'deiodinase as an activation pathway. These conversions are illustrated in Figure 12.30.Given the importance of thyroid hormones in regulation of metabolism, it was only natural for animal scientists to consider if administration of thyroid hormones might be used to improve metabolic rate to support enhanced growth or development. For example, involvement of the thyroid gland in maintenance of lactation has been known since reports in the early 1900s showed that milk yield was reduced in Ihyroidectomized goats. By the 1930s, it was shown that thyroidectomy of dairy cows reduced milk yield and, conversely, that treatment with T4 increased milk yield by approximately 20%. Because T4 is also efficacious when fed, these reports stimulated much interest in the practical utilization of the hormone to increase milk production in cattle. This was made economically feasible by the relatively low cost of manufacture of T4 and other thyroactive iodinated proteins. However, results of multiple studies showed that while feeding T4 (or iodinated proteins) increased milk production by 10-40%, the galactopoietic effect was of variable duration and milk production returned to normal or below normal levels despite continued treatment.
The galactopoietic effect of T4 supplementation depends on a general increase in body metabolism. Thus, it is not effective when cows are in early lactation (and negative energy balance) and already mobilizing body reserves to meet the energy demands of lactation. A general increase in body metabolism at this time would be counterproductive to meeting the nutrient demands of lactation. It was concluded that T4 treatment should not be initiated before midlactation and that the energy density of the diet should be increased during treatment because feed intake does not increase in proportion to increased energy utilization. Furthermore, upon withdrawal of treatment, a hypothyroid condition ensues that exacerbates the usual decline in milk yield in late lactation. Despite the initial interest in thyroid hormone supplementation to increase milk yield, the temporary nature of the milk yield response and frequent undershoot below normal production afterward led to the conclusion that its adaptation would be of minimal value.
Although T4 is the predominant thyroid hormone in circulation, it essentially serves as a prohormone because it has little if any biological activity. The most metabolically active thyroid hormone, T3, is produced by enzymatic 5'deiodination of T4 within the thyroid and peripheral tissues. Changes in the extra thyroidal activity of T4-5'-deiodinase (5'D) alter localized T3 availability. Activity of the enzyme also varies with physiological state. For example, with onset of lactation in rodents and ruminants, there is an increase in 5'D in the mammary gland and a decrease in the liver. These changes are believed to maintain a euthyroid state in the lactating mammary gland despite the fact that the body is hypothyroid as a whole. The transfer of iodine, iodinated nonhormonal compounds, and thyroid hormones through the mammary gland into the milk further exacerbates a systemic hypothyroid condition. Maintenance of a euthyroid state in the lactating mammary gland in the midst of a functional hypothyroid condition is consistent with increasing the metabolic priority of the mammary gland and providing T3 to heighten the effect of other galactopoietic hormones. For example, this occurs in response to treatment of cows with exogenous bST (Capuco et al., 1989).
The relationship between GH and thyroid hormones is not limited to GH-induced alterations in 5'D during lactation and galactopoiesis. There is a close relationship between thyroid hormones, thyroid hormone metabolism, and GH and IGF-I synthesis. Mechanistically, T3 can alter hepatic GH receptor binding and thus enhance GH stimulation of IGF-I synthesis. Alternatively, T3 can also increase IGF-I synthesis in the absence of GH. It is worth noting that in those situations when GH does not stimulate IGF synthesis (e.g., during food restriction, fetal development, sex-linked dwarfism, and hypothyroidism) there is evidence for T3 deficiency. In addition, T3 serves as a regulator of GH synthesis by the pituitary. Conversely, GH can alter synthesis of 5'D and therefore peripheral production of T3 (Capuco et al., 1989).
Calcitonin
It would be difficult to overstate the physiological relevance of the thyroid hormones, but before we leave the thyroid gland behind, two other hormones are also synthesized in the thyroid or the parathyroid glands. CT is a 32-amino acid peptide whose major function is to inhibit the enzymatic action of osteoclasts in compact bone to reduce resorption of the inorganic matrix surrounding the cells. Parafollicular or C cells of the thyroid synthesize and secrete CT in response to changes in serum Ca concentration. As concentrations rise—hypercalcemia—the cells increase CT secretion, but with hypocalcemia, hormone secretion is inhibited so that degradation of bone matrix is enhanced and Ca concentrations can return to normal. Although therapeutic use of CT is limited in animals, in humans it is used in patients with Paget's disease (associated with abnormal bone resorption) and in some cases of osteoporosis.
Parathyroid hormone
Parathyroid hormone or PTH works with CT to provide even greater control over calcium metabolism and ultimately improved homeostasis. The parathyroid glands usually occur in four pairs located near the poles of the two lobes of the thyroid gland. They are typically small clusters of tissue, -50 mg each. Cells that are actively synthesizing and secreting PTH are called chief cells. PTH is initially produced as a precursor (preproPTH) of 115 amino acids that has 25 amino acids cleaved in transit from the RER to the Golgi. Once in the Golgi, removal of 6 additional amino acids yields biologically active PTH that is secreted in secretory vesicles by exocytosis.
The primary effect of PTH is to increase calcium and decrease phosphate concentrations in extracellular fluids. These effects are mediated by actions in multiple tissues. Since compact bone is the most abundant mineral reserve in the body, it is a major target tissue. An early effect of PTH is stimulating the passage of calcium out of osteocytes and osteoblasts. Soon after, PTH activates osteoclast activity, to increase enzymatic resorption of hydroxyapatite. Since the hydroxyapatite is essentially calcium, phosphate, and water, this action increases serum calcium and phosphate. Secondarily, PTH targets cells of distal convoluted tubules of kidney nephrons to increase calcium absorption from the filtrate and simultaneously targets cells of the proximal convoluted tubules to decrease phosphate reabsorption. These actions improve calcium status without a corresponding increase in serum phosphate. In addition, PTH enhances the kidney tissue production of ofl -hydroxylase, an enzyme that is needed for conversion of 25 hydroxy vitamin D3 into its more biologically active 1,25 dihydroxy analog. This is important because l,25-(OH)2-vitamin D acts like a hormone to stimulate the synthesis of calcium binding—transport proteins in the intestinal epithelial cells. Over the course of several hours or days this increases absorption of calcium from the gut and therefore serum calcium. Given its myriad of actions, it is reasonable to assume that PTH plays a more prominent role in calcium homeostasis than CT.
In domestic animals the most common disturbance in calcium metabolism typically occurs at the time of parturition and is most frequent in dairy cows (milk fever) and dogs. Affected animals exhibit severe hypocalcemia and often severe neuromuscular dysfunction. The animals frequently become recumbent (cows typically are immobilized) and in dogs there are often involuntary muscle spasms referred to as tetany or eclampsia. The problem arises from the sudden demand for calcium needed for milk production so that serum concentrations are no longer maintained within usual limits. Treatment typically involves infusion of glucose and calcium. After treatment animals generally recover mechanisms to maintain calcium concentrations.
The etiology of milk fever is complex but it does not seem to involve a failure of PTH secretion. However, it may be that responsiveness of PTH target tissues (receptor expression) or vitamin D activation is impaired. In dairy cows, a common management recommendation is to feed diets that are low in calcium in the period before calving. The idea is to induce mobilization mechanisms prior to the dramatic increase that occurs with the onset of lactation so that the increased demand can be met. Figure 12.31 illustrates changes in serum calcium around the time of parturition in horses and cows. Both exhibit a decrease but it is relatively more dramatic in the cow and especially those that exhibited milk fever symptoms. Also, the lowest serum calcium level in the mares does not occur until 2 days postpartum.
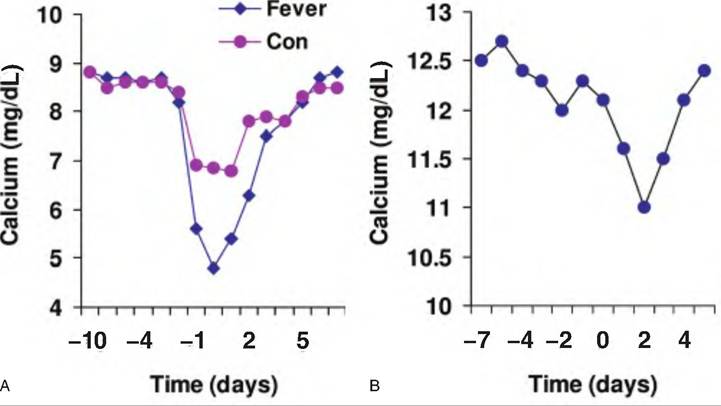
Fig. 12.31. Serum calcium. Panel A shows changes in serum calcium in Jersey cows around the time of calving. Animals that exhibited milk fever (diamonds) showed lower minimum values compared with herd mates (con) that did not exhibit milk fever. Data adapted from Goff et al. (1 995). Panel B illustrates periparturient serum calcium for mares. The decrease in calcium of small magnitude and minimum values occur 2 days after foaling. Data adapted from Martin et al. (1996).