Hormones of the anterior pituitary
Our earlier discussion briefly outlined the names and hypothalamic partners for the anterior pituitary hormones. WeTl now consider major physiological actions and some of the structural details of each of these hormones.
Given the importance of these hormones in regulation of growth, development, lactation, and reproduction, it is apparent that these agents are critical to understanding and improving animal agriculture. To illustrate this idea, consider the importance of these hormones in mammary development and function and therefore the dairy industry specifically and management of mammals more generally.The endocrine system, perhaps more than any other physiological system, is central in all aspects of mammary development (mammogenesis), onset of lactation (lactogenesis), and maintenance of milk secretion (galactopoiesis). Experiments beginning in the 1920s (Stricker and Grueter, 1928) showed that milk secretion could be induced in virgin rabbits by injecting an anterior pituitary extract. In 1933, Riddle et al. purified the protein responsible for the milk secretion response observed by Stricker and Grueter, naming it prolactin (Prl). Even now, the widely touted and utilized galac- Iopoietic effect of somatotropin or GH to increase milk production in lactating dairy cows had its foundation in studies by Asimov and Krouze in the 1930s. They showed that injections of pituitary extracts consistently increased milk production in lactating cows. Scientists describing and quantifying the potent effects of pregnancy on mammary growth and changes in the mammary gland at puberty spurred others to isolate and identify the steroid hormones estrogen and progesterone. Advances in purification techniques and understanding of steroid hormone chemistry allowed further studies, leading to the production of these steroids for widespread animal testing.
Although the existence of mammogenic and lactogenic substances from the pituitary had long been known, the efforts of C.H. Li and colleagues in the 1940s to purify larger quantities of Prl and GH were essential. Soon thereafter, specific roles for these hormones in the regulation of mammogenesis in rodents were delineated in classic ablation replacement experiments (Lyons et al., 1958; Nandi, 1958). In an extensive series of studies, triply operated (adrenalec- tomized, Ovariectomized, and hypophysectomized) rats and mice were treated with various combinations of purified hormones see if normal mammary development could be restored. Injections of estrogen and GH together caused proliferation of mammary ducts. However, treatment with estrogen, progesterone, Prl, and GH were needed for lobulo-alveolar development. The maximum ductular and lobulo-alveolar development, although still less that in pregnancy, was obtained in animals also given glucocorticoids. For some strains of mice, GH and Prl were both capable of stimulating lobulo-alveolar development. Interestingly, it was not until the 1960s that it was conclusively shown that human Prl and human GH were distinct proteins.
British researchers, focused on efforts to improve and maintain milk supplies during World War II, initiated many endocrine studies on mammary development and function in dairy animals (Cowie, 1980). For example, the effects of estrogen and progesterone on mammogenesis were extensively evaluated in attempts to induce lactation in nonpregnant animals. Although difficulties with needed surgeries and expense continues to limit use of ablation replacement experiments to study mammary development and function in cattle, Cowie et al. (1966) studied hypophysectomized- Ovariectomized goats and showed that mammary development comparable to that at mid-gestation could be obtained in animals treated with a combination of estrogen, progesterone, Prl, GH, and ACTHs. Such experiments served to confirm that at least general effects attributed to these hormones on mammary development in rodents also applied to mammary development in dairy animals.
Somatotropin (GH)
By the 1920s it was discovered that crude extracts prepared from homogenates of bovine pituitary glands could stimulate the growth of rats. The active agent was dubbed somatotropin after the Greek word for growth. Soon thereafter, the ability of such extracts to promote milk secretion in pseudopregnant rabbits and milk production in lactating goats were reported. Some of the more extensive early experiments with cows by the Russians Asimov and Krouze in the 1930s involved the treatment of more than 2000 cows with crude anterior pituitary extracts. Soon after, as part of efforts to increase food production during World War II, the British scientists Folley and Young and colleagues also studied effects of GH on milk production in cows and goats. They identified GH as the primary active galactopoietic component in bovine pituitary extracts. Other studies established dose response curves and confirmed that relative responses were greater in declining lactation, that gross milk composition was unaffected, and the generally potent effect of GH on milk yield in dairy ruminants. They concluded that use of pituitary GH would be highly profitable to individual farmers but that an inadequate supply limited the impact that GH could have in stimulating the national milk supply. For example, approximately 25 pituitaries are needed to produce enough GH for a typical daily treatment. These studies were the prelude to subsequent studies and ultimately large-scale use of recombinant bovine GH (bGH) in dairy cows (Etherton and Bauman, 1998; Bauman, 1999).
GH, like Prl, is a single-chain protein and the proteins share about 50% structural homology. GH is also structurally similar between species, for example, bovine GH is 192 amino acids (23,000 MW) but biological activity within species is characteristically distinct. As the name suggests, GH is closely associated with body growth. For example, soon after its characterization, it was hoped that bovine GH (bGH) might supply material to treat humans with impaired growth caused by hyposecretion of human GH (hGH).
However, it was soon discovered that bGH had no effect in primates. Some human patients were ultimately treated with hGH derived from cadavers but supplies were limited and unfortunately some samples contained agents (viruses and possibly prions) that made then harmful. More widescale therapeutic use of GH in humans had to wait until recombinant hGH became available.Somatotropin or GH, depending on the tissue, can act directly or indirectly to coordinate biochemical adaptations that chronically alter the metabolism of carbohydrates, lipids, and proteins. Although generalizations can be misleading, under most circumstances elevations in GH act to increase available nutrients by promoting mobilization of tissue stores. For example, GH tends to increase protein synthesis by promoting the uptake of amino acids while at the same time decreasing protein catabolism and promoting lipid mobilization. This acts to make fatty acids preferential fuel sources. Somatotropin directly or indirectly coordinates metabolic adaptations that promote increased milk production in the lactating dairy cow. These adaptations involve chronic alterations in carbohydrate, lipid, and protein metabolism in a number of tissues and serve to preferentially direct nutrients toward the mammary gland. This coordinated regulation to support the priorities of a physiological state is called homeorhetic regulation.
In farm animals, this has been studied rather extensively in dairy cows because of the capacity of GH treatments to enhance milk production. In some of the early short-term studies, GH markedly increased serum triglycerides. A common belief related to nutrient regulation at the time of these studies was that nutrient use largely involved a competition between organs for needed substrates. This means that an increase in milk production in response to, for example, treatment with GH would require either an increase in the metabolism of the mammary gland to better "fight" for nutrients or reduction in nutrient use by other tissues to support increased nutrient demands of the mammary cells for milk synthesis and secretion.
Mechanisms for GH's actions were generally thought to depend on acute effects to "push" metabolism of peripheral tissue to reduce competition and thus favor the lactating mammary gland. On the surface many acute effects of GH seem to support this model. These GH-Stimulated responses include glycotropic (reduced response to insulin in glucose tolerance testing), diabetogenic (hyperglycemia and glycouremia), and lipolytic (increases in blood nonesterified fatty acids) activity. Given these effects, a number of researchers were concerned that GH induction of these responses would promote metabolic problems in lactating cows. There was concern that acute mobilization of adipose tissue, particularly in postpartum animals already in a negative energy balance, might suffer increased metabolic disorders—ketosis, fatty livers. However, these "extra" nutrients were preferentially directed toward the mammary gland for use in increased milk production.About this time, the concept of homeorhesis came to be applied to the high milk production of genetically superior dairy cows. The overriding idea was that homeorhesis does not act to defeat homeostasis but rather that chronic changes in physiological controls allow coordination of physiological processes to support major physiological events (the definition of homeorhesis). Examples include physiological support for fetal development and reproductive performance, or in this case a sustained high level of milk production while at the same time preserving homeostasis. The preponderance of evidence suggests that GH enhances milk production largely by partitioning nutrients to support milk production by both direct and indirect actions, but GH does not alter digestibility of nutrients. In dairy cows given exogenous GH, metabolism is altered in an organ-specific fashion to establish nutrient flux toward the mammary gland. Increased appetite and dietary intake interact to moderate some of the demand for nutrients some days after the start of treatments (Box 12.1).
Box 12.1 Hormones in my milk—Yes please!
As a lactation biologist (RMA) I am intrigued by the popular press and general discussion about hormones and milk. As physiology students you must be coming to realization that milk (like other secretions) is a biological fluid. So there should be no surprise that milk like other secretions and fluids is likely to contain a variety of hormones, growth factors and other potentially biologically active agents. As described in the lactation chapter, the relative amount of these materials is much higher in the colostrum than "mature" milk but nonetheless small amounts of a variety of hormones and growth factors are present in milk. It is also relevant that there is no difference in the nutritional value or presence of these agents in milk from organic versus conventional dairies.
It is likely that some of these agents have impacts on the neonate but gut closure and activation of digestive function is also likely to limit effects in older offspring or adults. However, it is also true that a number of peptides which are biologically activity can be produced by the digestion of a number of milk proteins, especially the caseins. A key question is whether or not these substances actually have effects within the gut or induce signaling or other responses that impact the body of the suckling young or consumer. Just because peptides with biological activity are generated does not mean they elicit responses normally (after consumption of milk). Nagpal et al. (2011) review aspects of this topic from the point of view of functional foods.
The somatomedin hypothesis
The primary function of GH is promotion of linear growth, but many of its growth-promoting effects are indirect because of its ability to stimulate the liver to produce IGF-I. This is called the somatomedin hypothesis. Initially, IGF-I and IGF-II were called somatomedins, to account for their relationship with GH action. However, once structures of the molecules were delineated it became clear that the molecules were similar to insulin, hence the new names. Regardless, it is now certain that many actions originally attributed to GH are mediated by IGF-I. But, it should not be forgotten that many tissues also express GH receptors. The liver is the primary source for IGF-I in circulation but IGF-I is also produced locally in many tissues. It may be that locally produced IGF-I is as important as circulating IGF-I. The importance of nonliver sources became clear from knockout mouse studies. In these experiments, genetic engineering techniques were used to block or knock out normal liver IGF-I synthesis. Despite this, these animals exhibited essentially normal growth and development. This suggests that for many situations, local production of IGF-I can replace and/or supplement circulating IGF-I supplied by the liver. Other complications include the discovery of a family of IGF-I binding proteins (IGFBPs). These molecules also appear in circulation and are produced locally in many tissues. Depending on conditions, these proteins can either inhibit or enhance biological effects associated with IGF-I. Some of these relationships are illustrated in Figure 12.17. The black arrows illustrate the pathways associated with the "classic" somatomedin hypothesis and the red dashed arrows, more recent findings related to local production of IGF-I and IGFBPs.
Figure 12.18 demonstrates synthesis of IGF-I by stroma cells in the bovine mammary gland. In this
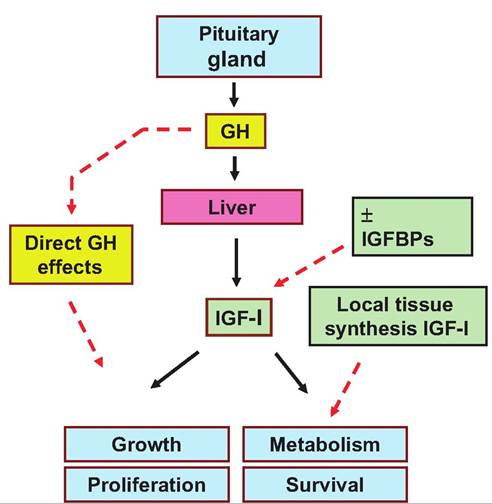
Fig. 12.17. The somatomedin hypothesis. Relationships between secretion of pituitary GH and liver IGF-I are illustrated by the solid black arrows. Dashed red arrows indicate direct effects of GH, significance of local tissue production of IGF-I, and the role of IGF-l-binding proteins control of biological actions of IGF-I.
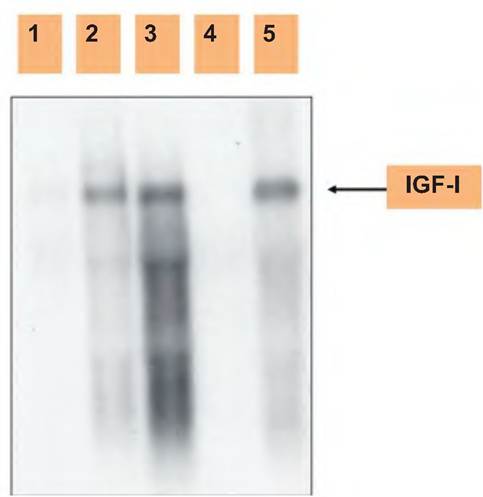
Fig. 12.18. Expression of IGF-I. Shown is a northern analysis of IGF-I mRNA expression of mammary epithelial cells or stromal tissue prepared from heifer mammary. 1, 4: Epithelial cell; 2, 3: stroma; 5: liver control. Notice there is a signal for IGF-I in samples of mammary stroma and liver RNA but not in mammary epithelial cell RNA. Data adapted from Berry et al. (2003).
instance, stromal tissue and isolated mammary epithelial cells were tested for the presence of mRNA for IGF-I. These results suggest that IGF-I mRNA detected in samples of mammary parenchymal tissue is actually synthesized by stromal tissue cells that surround the developing mammary ducts (Akers et al., 2000, 2005).
The physiological relevance of GH becomes apparent in situations with either hyper- or hyposecretion. Hypersecretion of GH prior to puberty produces gigantism because of rapid, prolonged proliferation of growth plate chondrocytes. If excess secretion is initiated after closure of the epiphyseal plates of the long bones, the result is acromegaly. This is characterized by enlargement of extremities and facial bones. Failure to produce sufficient GH in young animals leads to production of stunted development and a type of dwarfism. Given its importance to growth, it should be no surprise that there is a great deal of interest in understanding effects of GH related to growth and development in farm animals. For example, do strains or lines of naturally rapidly growing animals produce more GH? Could administration of exogenous GH or perhaps immunization against somatostatin be used to promote GH secretion and more rapid growth? Aside from indirect effects of GH on chondrocytes and protein accretion, changes in GH secretion are certainly important in both short-term and more chronic changes in metabolism.
GH secretion
Figure 12.19 illustrates average concentrations of GH and insulin in the blood of cows during the lactation cycle. In early lactation, the rapid increase in milk production causes the animals to exhibit a negative energy balance. In other words, feed intake does not keep up with demands. After about 90 days of lactation, peak milk yields are past, feed intake continues to rise, and the animals begin to go into a positive energy balance. In the later stages of lactation, body condition increases and the animals begin to deposit excess energy in the form of subcutaneous fat. Changes in average concentrations of blood insulin and GH mirror these physiological changes. Insulin is typically elevated to drive glucose into storage but GH is elevated to mobilize nutrients. In early lactation, insulin concentrations are suppressed while GH is elevated. Toward the end of lactation this situation is reversed.
Secretion of GH is not constant. Instead it is secreted in bursts or pulses so that with frequent blood sampling blood concentrations periodically spike. In
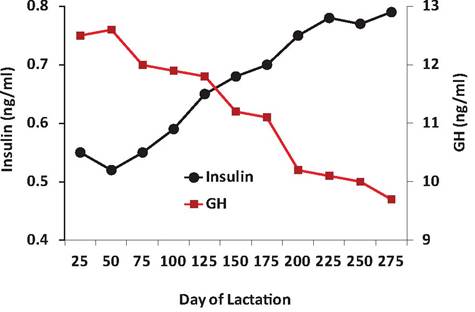
Fig. 12.19. I nsιιlin and GH during lactation. Basal insulin and GH in serum of cows during lactation. Adapted from Herbein, J. H., R. J. Aiello, L. I. Eckler, R. E. Pearson and R. M. Akers. 1985. Glucagon, insulin, growth hormone, and glucose concentrations in blood plasma of lactating dairy cows. JDS 68: 320-325.
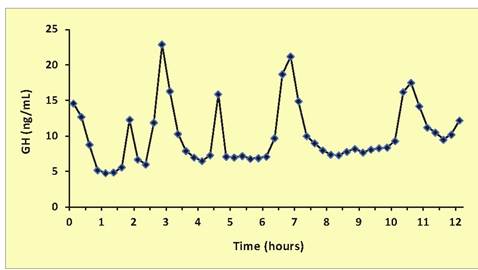
Fig. 12.20. Pattern of GH secretion. In this Holstein heifer calf, note there are several apparent secretory episodes during this 12-hour period of time. Samples were collected at 15-minute intervals for assay. Average GH concentration was 9.4ng∕mL.
younger animals and in males, these events are more frequent and the amplitude of these secretory spikes greater. This results in higher average or basal GH concentrations as well. As with many hormones, there are also intrinsic overall patterns of secretion that follow circadian and ultradian rhythms (Lefcourt et al., 1995). Figure 12.18 illustrates changes in serum GH over the course of 12 hours in a Holstein heifer calf (Fig. 12.20).
Prolactin
As its name suggests, Prl has undoubtedly been the most widely studied hormone related to lactation and mammary growth. However, it is very clear that Prl exhibits a very wide variety of physiological actions. For example, increased secretion of Prl is associated with onset of nest building and brooding in birds. Prl is also important in kidney and immune system function. In the past 20 years, it has been established that native Prl is really part of a family of structurally related protein isoforms or variants. The Prl gene is transcribed not only in the Iactotrophs of the anterior pituitary gland but also by cells in the placenta, hypothalamus, mammary gland, and in lymphocytes. Moreover, either as it is secreted or after interaction with target cells, some Prl is enzymatically modified to become cleaved, phosphorylated, or glycosylated. Thus, the pleiotropic actions of Prl may ultimately be attributed to the presence of these different isoforms of the hormone. As has been suggested (Das and Von- derhaar, 1997), the signaling pathway for Prl stimulation of cell differentiation compared with that for Prl signaling of cell proliferation needs to be deciphered to truly appreciate the role of Prl in one of its specific target tissues, the mammary gland.
At least three distinct forms of the Prl receptor are also known to exist. The three forms of the receptor exhibit differences in their cytoplasmic domains. The long form is 90kDa and differs from the short form (40kDa) because of differential splicing of mRNA transcribed from the Prl receptor gene. An intermediate version of the receptor is a deletion mutant of the long form that lacks 198 amino acids in the cytoplasmic domain. The intermediate form of the receptor is more sensitive to Prl and a major form of the receptor found in the liver also belongs to a superfamily of structurally related proteins that include GH and placental lactogen. This realization has facilitated the study of intracellular signaling pathways since the structural similarities suggest parallel similarities in mechanisms of action.
As a specific example, expression of the Prl receptor in the bovine or ovine mammary gland increases dramatically near the time of parturition in concert with lactogenesis and this level of expression is generally maintained during lactation. But there is no evidence for expression of different forms of the receptor to suggest that possible mammogenic versus lactogenic effects on mammary growth or function in ruminants is associated with a particular Prl receptor subclass (Smith et al., 1993).
Some of the best evidence for the importance of increased periparturient secretion of Prl in stage II lactogenesis in cows has come from experiments in which the administration of a dopamine agonist has been used to inhibit Prl secretion and correspondingly impair lactation. In ruminants where postpartum milking continues, administration of dopamine agonist oc-bromoergocryptine (CB154) reduced basal prolactin concentration about 80% and prevented the usual periparturient rise as well as milking-induced Prl rise during the first week postpartum. Differences in Prl secretion in control and CB154-treated cows are shown in Figure 12.21. Milk production was reduced 45% during the first 10 days postpartum. Lost milk production was associated with reduced synthesis of Oc-Iactalbumin, lactose, and fatty acids, as well as impaired structural differentiation of the mammary secretory cells. Selected effects are summarized in Table 12.9. Cows treated with exogenous Prl in addition to the agonist (to replace the periparturient surge in prolactin) showed no loss of milk production or effects on milk component biosynthesis or alveolar cell differentiation. An effect of Prl suppression and replacement during the periparturient period on milk production in multiparous cows is shown in Figure 12.21. Clearly, Prl is important in mammary cell differentiation and lactogenesis (Akers et al., 1981) (Fig∙ 12.22).
Prolactin secretion
Prl secretion, like that of GH, is not only episodic, but it is also is acutely increased by a variety of stimuli. Average concentrations are higher with warm temperatures and longer photoperiods. Manipulation of photoperiod has been used to increase livestock productivity for many years. Examples include manipulation of photoperiod to increase egg production in chickens or to manipulate breeding activity in seasonal breeders such as horses and sheep. Early studies noting the positive effect of photoperiod on Prl secretion prompted a landmark investigation by Peters et al. (1978) on the effects of increased photoperiod on milk production in cows. They showed that exposure of lactating cows to a long day photoperiod between September and April in Michigan significantly increased milk yields (2.0kg∕day) compared with cows exposed to the ambient photoperiod. This effect has been confirmed by numerous other research groups in North America and Europe, in latitudes
Image not available in this digital edition
Image not available in this digital edition
Fig. 12.21. Serum Prl in cows at calving. Changes in serum Prl in cows given ergrocryptine (CB-154) around the time of calving are shown. Note the usual surge in Prl secretion at calving is blocked and that average Prl concentrations before and after calving are reduced but secretion of Prl is not completely inhibited. Adapted from Akers et al. (1981).
Fig. 12.22. Milk production after suppression of Prl. Milk production per milking in control, CB154-treated, and CB154+Prl- treated cows is shown. Note the marked decrease in milk yield of cows given CB154 and that that milk yield is restored to normal for cows also given Prl. Data adapted from Akers et al. (1981).
Table 12.9. Mammary biochemistry after prolactin suppression.
| Treatment | ||||
| Prepartum | Postpartum | CB154 | CB154 + Prl | |
| Lactose synthesis (μg∕h∕100mg) | 39 ± 63 | 552 ± 70 | 327 ± 63 | 628 ± 81 |
| CX-LactaIbumin (μg∕mg∕protein) | 1.7 ±0.04 | 5.4 ± 0.4 | 2.8 ± 0.4 | 6.8 ± 0.5 |
| RNA (g) | 23.6 ± 2.1 | 87.2 ± 16.8 | 56.0 ± 8 | 91.5 ± 12.8 |
| DNA (g) | 27.9 ± 2.9 | 46.0 ± 3.8 | 40.1 ± 4 | 42.2 ± 3.8 |
ranging from 39 to 62oN. In cattle, increasing the duration of daily photoperiod from 8 hours light:16 dark to 16 hours light:8 dark increases Prl concentrations in the blood several fold. It is, however, important that animals experience at least a minimal dark cycle, since animals on a continuous lighting regimen appear to revert to a short-day photoperiodic response pattern. However, recent work suggests that increases in milk production are likely mediated by changes in the IGF-I axis rather than Prl (Dahl et al., 2000).
In various species (cows, goats, sheep, human, rat), stimuli associated with milking or sucking promote the secretion of Prl. However, the secretion of Prl or other hormones in response to milking or suckling does not directly depend on the removal of secretions, since teat stimulation alone can also induces Prl secretion in nonlactating animals. Little is known about the development of this neuroendocrine reflex, but it is affected by stage of development. For example, in an experiment with 3-, 6-, and 10-monfh old heifers, three of six 3-month-old heifers showed a moderate increase in blood Prl with mimic hand milking but only two of six for the 6-month-old heifers and none of six for the heifers at 10 months of age. In contrast, for heifers tested during gestation, all of the heifers responded and Prl secretion increased monotonically as gestation advanced (Fig. 12.23). The response continues into lactation with Prl routinely secreted with each milking, but in cows, the magnitude of the response declines with advancing lactation (Akers and Lefcourt, 1982).
Image not available in this digital edition
Follicle-stimulating hormone and luteinizing Hormone
It is fitting that follicle-stimulating hormone (FSH) and luteinizing hormone (LH) be discussed together. In both males and females, together these gonadotropins regulate reproductive physiology. But first, some of the basics: both hormones are have an a and β chain. In fact, TSH shares this structural similarity. For each of these hormones, the biological action of the hormones depends on the β chain. FSH has a major effect on the growth and development of ovarian follicles. This reflects the capacity of the hormone to induce estrogen secretion by the theca cells surrounding developing oocytes. As the waves of follicles develop during the estrus cycle of the cow, for example, only one of a cohort of follicles becomes dominant. This follicle proceeds to enlarge and develop so that it becomes the follicle destined for ovulation. Closely tied with FSH control of estrogen secretion and changing sensitivity of the hypothalamus, there is an acute increase in secretion of LH, which promotes ovulation of the dominant follicle. This is the ovulatory surge in LH. This not only causes rupture and release of the
Fig. 12.23. Prl secretion after teat stimulation. Panel A shows changes in serum Prl concentrations before, during, and after teat stimulation of pregnant Holstein heifers sampled at 100, 150, 200, or 250 days of gestation. Note the monotonic increase in response with advancing gestation. Panel B shows Prl secretion in heifers in response to machine milking 30 days postpartum. Data adapted from Akers and Lefcourt (1982).
egg, but it also promotes remaining follicular cells to differentiate into luteal cells and create a corpus Iuteum (yellow body). This luteinization process explains how the hormone gets its name. More importantly, as these cells are luteinized they begin to secrete progesterone, which prepares the uterus to accept a fertilized oocyte. In the absence of fertilization, the corpus Iuteum of CL degenerates. Figure 12.24 illustrates relative changes in secretion of estrogen and progesterone during the estrus cycle and relationship to changing secretion of LH.
A preovulatory surge in the LH begins about 24 hours prior to ovulation in most domestic species (cow, goat, sheep, and pig); this triggers critical changes in the follicle to impact its endocrine activity
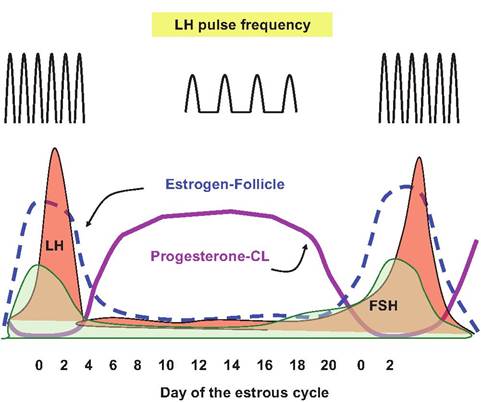
Fig. 12.24. Hormones during estrus cycle. Relative changes in concentrations of estrogen, progesterone, FSH, LH, and LH pulse frequency during the estrus cycle in the cow are illustrated.
and culminate in ovulation. Poorly characterized factors in the granulosa cells prevent the premature conversion of the cells into luteal tissue until after ovulation. Increased LH allows for the resumption of meiosis by the oocyte and removes the block on luteinization of the granulosa cells. This effectively converts the cells from predominately estrogen production to progesterone secretion as illustrated in Figure 12.24. The process actually begins in the period before ovulation but the LH surge hastens the decline in estrogen secretion and concomitant rise in progesterone secretion. Indeed, the progressive increase in secretion of estrogen by the rapidly developing follicles just before ovulation acts to signal the hypothalamus and ultimately the anterior pituitary that conditions are appropriate for ovulation.
Secretion of FSH and LH is controlled by release of GnRH from the hypothalamus. Secretion of GnRH occurs in bursts or spikes so that the corresponding secretion of FSH and LH also occurs episodically. However, at least two clusters of nerve cells or nuclei in the hypothalamus (ventromedial and arcuate nucleus) produce GnRH in a tonic release center. This secretion pattern produces short-lived pulses of GnRH sometimes compared with the drip, drip of a leaking faucet and corresponding brief bursts of FSH and LH secretion.
As illustrated by the pulse frequency for LH illustrated at the top of Figure 12.24, gonadotropin secretion is increased during the follicular phase of the estrus cycle but decreased during the luteal phase. Acute control to allow for increased pulse frequency but decreased pulse amplitude allows for final growth and development of the dominant follicle. The rapid increase in estrogen creates a positive feedback loop at the level of the hypothalamus. Increased estrogen from the rapidly developing follicle increases GnRH pulses. This produces more FSH secretion, thus more estrogen, and further GnRH secretion. This cycle is broken by the maturation of the follicle and the LH surge. As estrogen concentrations decline, the positive feedback is lost, and inhibitory progesterone concentrations increase. This likely explains the rapid rise and equally rapid decline in LH. However, since GnRH generally stimulates the secretion of both LH and FSH, what explains the abrupt LH surge without a corresponding peak in FSH secretion?
This is explained by the presence of a second cluster of hypothalamic nuclei (preoptic nucleus, anterior hypothalamic area, and Suprachiasmatic nucleus) called the surge or preovulatory center. The cells in this area to respond to a threshold level of estrogen. As the estrogen concentration increases rapidly just before estrus, this induces a flood of GnRH secretion (like turning on a facet compared with the drip of the tonic center) and a corresponding marked increase in LH called the ovulatory peak. Although GnRH usually stimulates secretion of both FSH and LH, it is thought that secretion of inhibin by the follicle at this time suppresses FSH secretion.
In males LH, plays a major role in regulation of testosterone secretion by the Leydig cells located in the interstitial tissue surrounding the seminiferous tubules of the testes. Within the tubules, FSH stimulates development and function of the Sertoli cells that nourish and regulate developing spermatozoa (Box 12.2).
Thyroid-stimulating hormone
TSH is a glycoprotein that shares a similar structure to FSH and LH, having an a and a β chain. The a subunits are identical and the β chains confer biological specificity. During synthesis the chains are produced separately then joined as glycosylation occurs in the Golgi. Mature TSH ultimately binds to receptors in the thyroid to promote thyroid uptake of iodine, iodination to produce T3 and thyroxine (T4) by the follicular cells of the thyroid gland. These events are triggered by adenylyl cyclase and subsequent synthesis of cyclic AMP. With more prolonged stimulation the thyroid gland is enlarged and follicular spaces are filled with stored thyroglobulin. Further details will be described in our discussion of the thyroid hormones.
Secretion of TSH is regulated by both stimulatory and inhibitory influences. The most direct control is from hypothalamic TRH that promotes secretion. However, somatostatin also acts as a potent inhibitor
Box 12.2 Practical endocrinology
Is endocrinology directly important to animal producers? The short answer is yes very much so. One easy example is the use of oxytocin with animals that have issues with milk letdown (cows, sheep, and goats). Another is the very wide-spread use of lutalyse® (dinoprost tromethamine) which is an analog of the prostaglandin F2α that is used to cause regression the corpus luteum. The product is used extensively in estrus synchronization protocols to allow for timed breeding of beef and dairy cows as well as treatment for endometritis. Other products used in a variety of reproduction issues include: (Cystorelin®, Factrel®, or Fertagyl®) which are preparations of GnRH. The history and current protocols for estrus control and synchronization are outlined in recent reviews (Wiltbank et al., 2011; Lamb et al., 2010). Pregnant mare serum gonadotropin (PMSG) or human chorionic gonadotropin (hCG) have FSH and/or LH activity and are used in superovulation protocols that are used in the production and harvest of embryos from genetically superior cows.
Finally there are a variety of anabolic steroids that are widely used as growth stimulants primarily in beef cattle (Capper and Hayes, 2012). Depending on the product these agents have estrogen, testosterone, progesterone, zeranol, or tenbolone activity (Stephany, 2010). The products are designed for specific animals and/or physiological states. Others have described use of β-Adrenergic agonists to modify tissue composition in meat animals (Smith, 1998; Bohrer et al., 2013).
of secretion. Under usual circumstances, negative feedback from T4 at the level of the pituitary and T3 at the level of the hypothalamus provides acute control. However, since TSH and the thyroid hormones are so critical in metabolism it should not be surprising that other Uietabolically active hormones have an impact. For example, estrogen enhances the effect of TRH on secretion of TSH but GH and glucocorticoids dampen the effectiveness of TRH. At higher areas in the brain the activities of bioaminergic and peptidergic neurons also impact secretion of TRH and therefore TSH. For example, dopaminergic agonists (bromocriptine) inhibit TSH secretion but antagonists increase TSH release. Well-known effects of cold and stress to increase metabolic rate begin with alterations in neural activity that ultimately impact hypothalamic secretion of TRH. This cascade of events leads to secretion of TSH and then T3, which is ultimately responsiblefor increased metabolism.
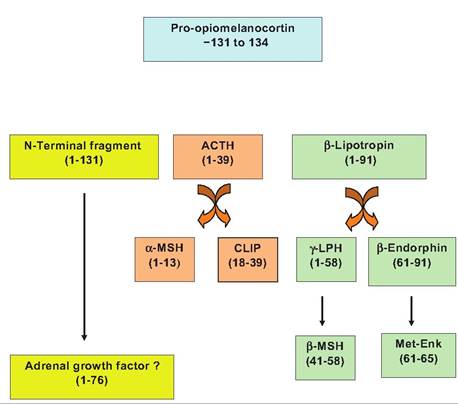
Fig. 12.25. Processingofpro-Opiomelanocortin. Abbreviations include corticotropin-like intermediate lobe peptide (CLIP), melanocyte-stimulating hormone (MSH), lipotropin (LPH), and methynine enkaplin (Met-Enk).
Aclrenocortropic hormone
Adrenocortropic hormone (ACTH) is a 39-amino-acid peptide processed from a larger precursor molecule called pro-opiomelanocortin (POMC) as illustrated in Figure 12.25. In the Corticotrophs of the anterior pituitary, the mRNA from the POMC gene directs the synthesis and processing of the transcript to yield at least eight biologically active fragments. ACTH primarily stimulates the secretion of glucocorticoids from the cortex of the adrenal gland. Two of the fragments are derived from ACTH. The first 13 amino acids of ACTH are β-MSH and the ACTH 18-39 corticotrophin-like intermediate lobe peptide (CLIP). The larger precursor POMC appears in the intermediate lobe of the pituitary in many species and is associated with secretion of MSH. Biological effects of adrenal steroids will be considered in a subsequent section.
While ACTH is the focus, a family of diverse peptides is derived from POMC. Secretion of ACTH is greatly impacted by neural factors and hormones. These agents modify the secretion of CRH, which, like GnRH, is secreted in an episodic pattern. In many species there is also a diurnal rhythm in the ACTH secretion pattern and therefore a diurnal secretion of adrenal steroids. Negative feedback loops involving cortisol are important in control of ACTH. One mechanism is sensitive to the rate of change in circulating glucocorticoids (fast feedback) and another responds to the absolute concentration of cortisol in a thresholdlike response (slow feedback). These effects are mediated at both the pituitary and hypothalamus. Furthermore, a host of stresses (pain, hypoxia, cold exposure, etc.) can override the usual rhythmic secretion of ACTH. This is classically thought of as the part of the fight- or-flight reactions and the need for nutrient mobilization and is associated with more prolonged secretion of glucocorticoids.