Epidemiology
15.2.1 Spatial and temporal patterns of infection in host populations and the environment
Clustering of hosts and pathogens in the environment can lead to hot spots of disease at local, regional, national and international levels.
Scotland contains regional hot spots of MAP in rabbits and this mirrors the distribution in cattle (Greig et al., 1999). MAP-infected rabbits, and the associated risk of interspecies transmission, are highly clustered in the environment (Greig et al., 1999). However, this is mostly due to the clustered distribution of the general rabbit population in the environment. In addition to spatial clustering, there is also temporal variation in infection levels. For example, within a farm with an overall MAP infection prevalence of 40%, the temporal distribution of infection in rabbits followed a cyclical pattern, with a peak in spring of 55.4% and a low in summer of 19.4% (Judge et al., 2005a).15.2.2 Inter- and intraspecies routes of transmission within wildlife communities
The faecal-oral route is generally seen as the major route of horizontal transmission in ruminant species, although vertical and pseudovertical (via the ingestion of colostrum or milk contaminated with faeces) have also been shown to occur (Sweeney, 1996). Few studies have examined intra- and interspecific routes of transmission within wildlife communities. MAP has been isolated from the testes, uterus, placenta and fetuses of rabbits, suggesting the possibility of both sexual and vertical transmission (Judge et al., 2006). MAP has also been isolated from the milk of lactating rabbits. However, as rabbits wean their young on their own cecotropes and faeces, the role of milk vs faecal-oral transmission may be difficult to separate. In a population of rabbits with 42% infected adult females, field studies suggest that vertical, i.e.
transplacental and/or pseudovertical, transmission can occur in 14% of offspring entering the population at 1 month of age, and this equates to a probability of infection via this route of 0.326 (Judge et al., 2006). In the same population, an analysis of the prevalence of infection as a function of age estimated that the probability of infection via horizontal transmission (including interspecies transmission) was as high as 3.7% per month. Given an average 18-month lifespan of a wild rabbit, this estimate suggests a high probability of acquiring MAP infection in an affected population (Judge et al., 2006).Carnivores have relatively little direct contact with contaminated pastures or farm buildings, and it is therefore more likely that they become infected with MAP through ingestion of infected prey. MAP has been isolated from the mesenteric lymph nodes and intestinal tissue of infected rabbits and rats (Greig et al., 1997, 1999; Florou et al., 2008), and carnivores often ingest entire rodents and large enough proportions of rabbit carcasses to include lymph nodes and intestinal tissue. Lagomorphs and small rodents form the majority of the diet of foxes, stoats and weasels in Scotland (Harris and Lloyd, 1991; King, 1991a, b; Leckie et al., 1998), and coyotes and raccoons are also known to feed on rodents and lagomorphs. This may explain the relatively high numbers of PCR positive samples from these species (Anderson et al., 2007). As detailed above, the prevalence of infection in carnivore species is generally higher than that in prey species. In one study, 62% of the predators (fox, stoat and weasel) sampled were infected compared with 10% of the prey species (rabbits, rats and mice). Transmission of pathogens through predation has been reported and is explained by carnivores’ higher trophic level in the food chain (Zarnke et al., 2000). At least part of the diet of the predators and opportunistic animals such as the opossum and a large part of the diet of corvids may consist of scavenging potentially infected prey carcasses (Mason and MacDonald, 1995; Anderson et al., 2007).
Therefore, scavenging may constitute an additional mode of paratuberculosis transmission.15.2.3 Potential livestock-to-wildlife routes of transmission
The association between MAP in non-ruminant wild species and paratuberculosis infections on farms suggests that interspecies transmission may occur between livestock and non-ruminant wildlife. Evidence for interspecies transmission has been provided by molecular typing of cattle and rabbit MAP isolates, which has shown that the same strains infect both species (Greig et al., 1999; Stevenson etal., 2009). Interspecies transmission has also been demonstrated experimentally in the case of rabbits infected with a cattle isolate of MAP (Mokresh and Butler, 1990) and calves infected with a rabbit isolate (Beard et al., 2001c). Given the high rates of shedding by clinically infected cattle and sheep (Cranwell, 199 7; Whittington et al.,
2000), it is likely that wild herbivores such as rabbits or hares sharing pasture with infected cattle become infected through the ingestion of contaminated grass.
Based on a sample of 20 farms across Scotland, Greig et al. (1999) found a statistically significant relationship between farms with an ongoing or recent paratuberculosis problem in cattle and the presence of similar strains of paratuberculosis in cattle and rabbits, providing further support for transmission between the species. In the case of MAP infection of granivorous or omnivorous rodents, exposure is more likely to occur in contaminated buildings than pasture (Beard et al., 2001a). Indeed, in one study, rodents from which MAP was isolated were all caught in buildings that housed cattle, whereas those captured in adjacent field margins or woodland were negative on culture (M.R. Hutchings, personal communication).
15.2.4 Potential wildlife-to-livestock routes of transmission
There are two main ways sheep and cattle can potentially ingest faecal matter from infected wildlife: grazing contaminated pasture and consuming contaminated farm-stored feed.
Grazing livestock tend to avoid pastures contaminated with their own and other species’ faeces through: (i) the selection of non-contaminated sward patches; (ii) reduced bite rate when forced to graze contaminated sward patches; and (iii) reduced grazing depth when grazing contaminated swards (Hutchings et al., 1998; Smith et al., 2009b). However, the only known species for which livestock do not use faeces-avoidance behaviour to limit exposure to parasites and pathogens is the rabbit. Sheep and cattle do not avoid rabbit faeces and graze contaminated pastures as they do non-contaminated pastures (Daniels et al., 2001; Judge et al., 2005b; Smith et al., 2009a, b), so the number of rabbit faecal pellets ingested by calves is directly proportional to the level of contamination of the pasture. Given a density of 10-59 rabbits/ ha for Scotland, the number of rabbit faecal pellets present at any one time on a grazing pasture is between 30,657 and 290,752 pellets/ha (Daniels et al., 2003a). Cattle ingest 1.3% of the faecal pellets, or between 389 and 3693 pellets/ ha grazed. If one-third of such pellets harbour live MAP organisms, one calf could ingest 1401329 infected faecal pellets/ha grazed. Sheep ingest an average of 0.5% of the faecal pellets, giving the potential to ingest between 146 and 1384 faecal pellets/ha grazed, of which 53-498 may be infected. Consequently, of the known non-ruminant wildlife host range in the UK, rabbits represent the greatest MAP risk to sympatric livestock due to the combination of a relatively high prevalence of infection, high population density of rabbits with access to livestock pasture and the lack of avoidance of rabbit faeces by grazing livestock. This risk is compounded by the ability of rabbit isolates of MAP to infect calves, which has been demonstrated experimentally (Beard et al., 2001c).Livestock are also susceptible to infection through feed. Wildlife excreta, particularly from rodents and birds, can contaminate feed during on-farm storage, after which faeces are likely to be ingested when the feed is given to livestock.
Levels of wildlife faecal contamination during on-farm winter feed storage have been quantified in feed stores on four farms in the east of Scotland by Daniels et al. (2003b). They reported a mean of 79.9 rodent faeces (95% confidence interval (CI): 3 7.5-165.9) and 24.9 bird faeces (95%CI: 14.3-41.7) deposited per m2 of stored feed per month. It was estimated that individual cattle and sheep could encounter 1626 and 814 rabbit faeces, respectively, over the winter. Similarly, infected rodents were found to be associated with livestock premises in Wisconsin (Corn et al., 2005; Anderson et al., 2007), and rodents were trapped in sheds housing goats and sheep in Greece (Florou et al., 2008). It is hypothesized that they became infected through scavenging livestock feed from the floor of the sheds where it has become contaminated with livestock faeces (Florou et al., 2008). Once infected it is likely that the rodents would contribute to contaminating the livestock feed and potentially contribute to the cycle of infection in the host community.Livestock that are fed supplementary feedstuffs are often unable to avoid contaminated feed. When presented with contaminated feed such as concentrates, cattle and sheep have some limited ability to actively avoid rat faecal pellets, but this is not the case for mouse faeces (Daniels and Hutchings, 2001). Moreover, the discriminatory power of livestock is limited, and group feeding that increases competition between animals further increases the rates of ingestion of rodent faeces (Daniels and Hutchings,
2001). Given such low levels of avoidance, even with extremely low probabilities of MAP transmission, infection may be maintained in livestock populations (Daniels et al., 2003b).
The importance of carnivores as paratuberculosis hosts at the wildlife-livestock interface is more likely to be indirect, since livestock are unlikely to come into contact with large amounts of their faeces. Such species could still play a role in disease dynamics as infected carnivores and corvids could theoretically transport MAP over far greater distances than most prey species.
For example, in the east of Scotland the home range size of rabbits is 6.3 ha (Hulbert et al., 1996; Daniels et al., 2003c) but is 4000ha for foxes (Harris and Lloyd, 1991).15.2.5 Persistence of infection in nonruminant wildlife
For a wildlife species to be considered a reservoir host, the pathogen must be capable of being permanently maintained within that host population. A longitudinal study of an island population investigated if MAP infection levels would be maintained in a rabbit population following the implementation of paratuberculosis control in sympatric cattle (Fox et al., 2018). It was found that applying MAP control measures in cattle led to a reduction in MAP seroprevalence in the cattle, however the same trend in prevalence was not echoed in the rabbit population (Fig. 15.1). This is the first study to show that efforts to control paratuberculosis in cattle do not reduce MAP prevalence in sympatric rabbits (Fox et al., 2018). This finding adds to mounting evidence indicating that if MAP became established in rabbit populations, they would represent a persistent and widespread
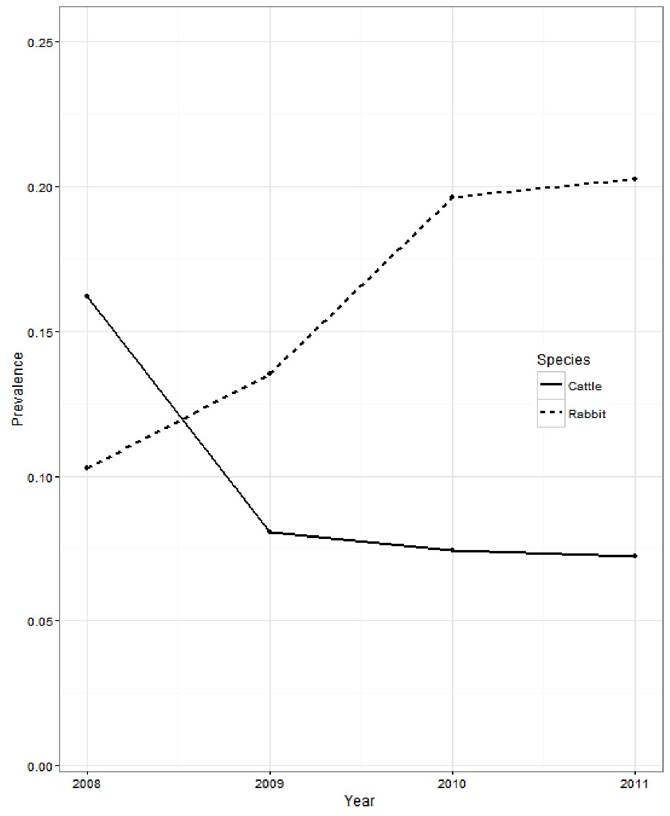
Fig. 15.1. Prevalence of paratuberculosis enzyme-linked immunosorbent assay (ELISA)-positive cattle and Mycobacterium avium subsp. paratuberculosis (MAP) culture positive rabbits from 2008 to 2011, following the implementation of a paratuberculosis control programme on all cattle farms from 2008. Composite means across 15 cattle study farms.
source of infection, potentially impeding control strategies in livestock. This supports the findings of Shaughnessy et al. (2013), who demonstrated that farms that responded well to MAP control and had low levels of MAP in sympatric rabbits could achieve effective paratuberculosis control in their herds, while farms with high MAP levels in rabbits were unable to control paratuberculosis in cattle despite implementing the same control measures. This suggests that the risks from rabbits translates to additional difficulties for the livestock industry to achieve control.
The findings of these field studies support the predictions of a mathematical model exploring the maintenance of MAP within rabbit populations. Due to the difficulties in experimentally demonstrating the long-term persistence of an infection in a host species, especially a slow-growing pathogen such as MAP, mathematical modelling is often used to synthesize known information on the host demography and pathogen transmission to predict if the infection is stable within a host population. Judge et al. (2007) developed a mathematical model and applied it to combinations of vertical (including pseudovertical) and horizontal rates of transmission, which covered a broad range of plausible values. This exploration of parameter space accounted for uncertainties in the value of R0 in wild populations. The long-run averages of the equilibrium prevalences were used to characterize disease persistence for each combination of horizontal and vertical/pseudovertical transmission rates.
This approach predicted that paratuberculosis would persist in rabbit populations at all values of the horizontal and vertical transmission parameters in the range estimated from the field data presented above, and in many cases at all values within 95% confidence intervals around this range. Indeed, the prevalence of MAP infection in the study site from which the transmission rates were estimated was 39.7%, falling within the prevalence ranges produced by the model. The persistence of MAP infection in rabbit populations suggests that they may act as a reservoir of infection (i.e. self-sustaining) for sympatric livestock. This may have significant implications for paratuberculosis control. For example, destocking is often considered as a management measure for disease control in livestock populations and was included in the National Ovine Johne's Disease Control and Evaluation Program in Australia (Sergeant, 2001; Whittington et al., 2003). However, this type of strategy fails to account for the presence of a wildlife reservoir and may therefore be unsuccessful if wildlife is infected. Furthermore, the possibility of the disease spreading from infected livestock farms to previously uninfected livestock areas through the dispersal of infected wildlife needs to be considered in control strategies. For the UK and other countries with high rabbit populations, it would seem unrealistic to eradicate rabbits at the scale necessary to prevent spillover infection in current hot spots of disease (Judge et al., 2005b). Consequently, management of disease should probably be restricted to rabbit population management (e.g. keeping population densities low), managing livestock-wildlife interactions (e.g. keeping animals, especially the most susceptible animals, away from areas with high rabbit densities) and good livestock management practices in terms of husbandry and movement between herds.
15.3