Estrous cycles, estrus, and ovulation
The ovaries are the source of the female gametes (ova) and estrous cycles in postpubertal animals. These cycles delineate periodic development or maturing of follicles, which results in release of eggs or ova that have the possibility of being fertilized.
The cellular events required to create eggs are called oogenesis. The period when the female is receptive to sexual activity is called estrus or more commonly heat. An estrous cycle is simply the time from the beginning of one estrus period to another. It is worth taking a moment to clarify some of this confusing terminology. Estrus is a noun. (A cow displays estrus or heat). Estrous is an adjective. (The average length of an estrous cycle in the Holstein dairy cow is 21 days). A review of British and European scientific literature shows that oestrus and oestrous are the equivalent terms for estrus and estrous.Animals that exhibit only one estrous cycle per year are monoestrous and estrus lasts for several days (dogs, wolves, foxes, and bears). As a reproductive strategy, having the extended period of estrus increases the chance of a successful mating. In contrast, polyes- trous species exhibit multiple cycles each year. However, these cycles are not necessarily uniformly distributed throughout the year. There are windows of inactivity when the animals are anestrous. These animals are seasonal polyestrous. Even among animals that are not seasonal breeders, there can be interruptions in the usual regular pattern. For example, in cattle that breed year round, there is typically an anestrous period for several weeks or months after calving. This can be especially pronounced in beef cows that are suckling their calves. Table 19.2 provides a summary of selected reproductive measures in some common farm animals.
It is important to appreciate that there can be substantial differences between breeds.
For example, onset of puberty in cattle can vary from a low of about 9 months in Holsteins to 19 months in Brahman cows. Similarly, Meishan pigs can show estrus as early as 3 months but Yorkshire gilts average about 7 months. In the case of onset of puberty, several external factors are also important. In sheep and goats, season of birth or photoperiod can act to hasten or delay onset of puberty. Presence or absence of the opposite sex during the peripubertal period impacts cattle and swine, as does the density of housing in swine. In a large grouping of gilts, normal puberty is 28 weeks, but in a smaller group (~3), puberty is often delayed until 32 weeks. Age at onset of puberty is also greatly influenced by body size and condition. Onset of puberty is a particularly important aspect of farm animal management. In dairy cattle it is an advantage to breed females as soon as practical so that the animals enter the milking herd earlier. This has to be coupled with the animals also having the necessary body size and condition to avoid calving difficulties and successfully compete with older stronger cows at the feed bunk. From a genetic view point, minimizing the age at puberty in males likely has the greatest benefit. In other words, like many aspects of animal production— growth, lactation—reproduction is also markedly impacted by management of the animals.Table 19.2. Average age or time for selected reproductive attributes.
We will use the dairy cow as our model species to describe the estrous cycle and process of ovulation. The general development of follicles is similar in most mammals. The ovary contains thousands of primary or dormant follicles. Essentially these are the ova, each of which is surrounded by a thin layer of cuboidal follicular or granulosa cells. In response to tonic secretion of FSH and LH, some of these primary follicles enlarge to become small antral follicles.
Antral follicles exhibit the appearance of a fluid-filled space between the oocyte in the center and the surrounding granulosa cells. These follicles provide the source of follicles that are eligible for activation during the next estrous cycles.Based on convention, the estrous cycle is divided into two phases, named after the dominant structures that are present on the ovary. The follicular phase is relatively short, -20% of the estrous period, and the preovulatory follicles that produce estradiol are in control. The follicular phase encompasses the time from the regression of the corpora Iutea to the time of ovulation. This is not to say that there is no follicular activity at other stages of the estrous cycle. Indeed, there are populations of follicles that sequentially develop throughout the estrous cycle. However, as in many things, timing is everything. As described in the following, it is only the dominant follicles that are part of a wave of development occurring near the time when the regression of the corpus Iuteum that has the opportunity to be qualified for ovulation.
It is now known that several waves (typically three) of follicular development occur during the estrous cycle in cattle. Beginning after ovulation, groups or clusters of small or medium antral follicles become sensitive to gonadotropins. These follicles are described as recruited follicles. Among this group of recruited follicles, several are selected and begin to mature. However, typically only one of these selected follicles will win the maturity race so that it becomes the dominant follicle. Other selected follicles in this class begin to undergo regression or atresia. Even the dominant follicle is destined to undergo atresia if it is so unlucky to have been recruited in the first or second wave of follicular development during the estrous cycle. This developmental process is illustrated in Figure 19.11. Figure 19.12 illustrates changes in major reproductive hormones during a bovine estrous cycle.
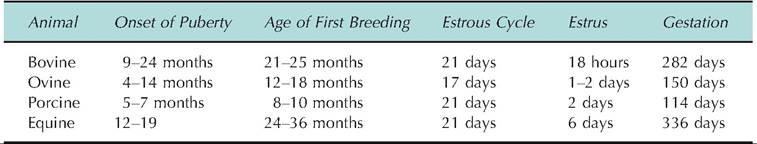
For a moment let's consider development of the ovary and specifically the relationship between the follicle and the oocytes. In the fetus, primordial germ cells migrate from the yolk sac to the immature ovary. These germ cells (now called an oogonium) become surrounded by a single layer of follicular cells. As the ovary matures, it is usual to find clusters or nests of these germ cells in the cortex of ovary (Fig. 19.9A). It is worth remembering that these cells undergo meiosis but they stop in the first prophase before the first division. In most animals, the first of the two meiotic divisions is completed, producing the first polar body, at about the time of ovulation. The essential point is that unlike spermatogenesis, where each primary germ cell produces four spermatozoa, the maturation of the oocyte creates only one mature ovum and three polar bodies. Thereafter some of these primary or primordial follicles are stimulated to growth progressing first to a secondary follicle stage. This is characterized by an increase in the number of granulosa cells surrounding the oocyte (Fig. 19.7B). A tertiary follicle or antral follicle is characterized by the coalescence of fluid that appears between the granulosa cells so that a prominent space is evident (Fig. 19.7C). With progressive development the stroma cells surrounding the follicle segregate into two distinct layers—the theca interna (closest to the layer of granulosa cells) and an outer theca externa. These layers give additional structural substance to the rapidly developing follicle. Moreover, the theca interna, if the follicle is a dominating ovulatory follicle, is destined to be a major source of steroid hormones. The granulosa cells are separated from the theca cells by the basement membrane. At this time the oocyte is typically displaced to one side of the follicle and antrum where it is surrounded by a cluster or cloud of granulosa cells called the cumulus oophorus (Fig. 19.8). Other granulosa cells remain in layers surrounding the oocyte adjacent to the theca interna.
These cells are referred to as the membrana granulosa. The granulosa cells adjacent to the ovum secrete glycoproteins that create or form a protective layer called the zona pellucida. Figure 19.9 shows examples of developing follicles.While we have used the cow as our primary model, there are dramatic differences between species. The LH surge and ovulation in most farm animals (cow, sow, ewe, and mare) take place regularly, independent of copulation (Fig. 19.10). These animals are spontaneous ovulators. In contrast, in rabbits, mink, camels, llamas, and alpacas, ovulation requires copulation. Such animals are induced ovulators. In these animals, appearance of the preovulatory LH surge depends on neural reflexes produced by vaginal simulation. These animals (often depending on season) have typical estrous cycles and associated follicular development, but the mature follicles undergo atresia if copulation does not occur.
Once ovulation occurs, the granulosa cells lining the now empty follicular cavity begin to divide, fill the space, and begin a process called luteinization in response to high levels of LH. The result is a structure
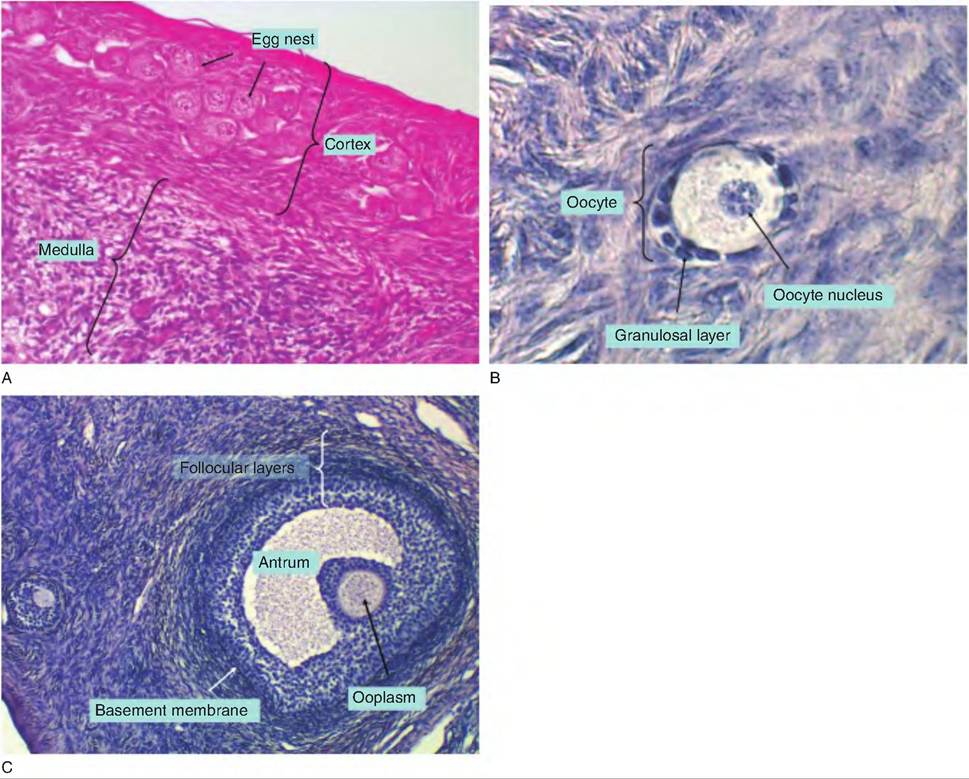
Fig. 19.7. Examples of follicular development in the bovine. Panel A shows a low-power view of the ovary. In the outer cortex, larger clusters of cells (egg nest with arrows) are primordial oocytes. The center is called the medulla. Panel B illustrates the appearance of a secondary follicle. The oocyte with its evident nucleus and large area of cytoplasm is surrounded by multiple layers of granulosa cells (layer). Panel C illustrates a tertiary follicle. The antral space or antrum and surrounding layers of follicular cells are evident. In this particular case, the nucleus of the oocyte is not apparent, but the ooplasm can be seen. The basement membrane demarcates the boundary between the follicular and theca cells.
called the corpus Iuteum (yellow body or CL).
The CL is a powerful endocrine tissue that produces large amounts of progesterone. Secretion of progesterone prepares the uterus to receive the ovulated ovum or ova. If fertilization and implantation of the ova is successful, this produces maternal recognition of pregnancy, and the usual regression of the CL does not take place. Progesterone concentrations in the blood are maintained throughout pregnancy. Some species are dependent on the CL for all of the progesterone needs, but in others the CL can actually be removed and pregnancy maintained because of progesterone produced by the placenta. Progesterone, especially in the later stages of pregnancy, is critical for mammary development needed to support lactation after the birth of the young.If pregnancy is not established, the CL must regress for the animal to continue estrous cycles. In most domestic species, the signal to induce CL regression is prostaglandin F2Of (PGF2oc). Secretion of PGF2oc begins to increase after ovulation with timing that corresponds with estrous cycle length in the species.
When PGF2OC secretion is sufficient, this stimulates a series of biochemical and cellular changes in the CL so that it regresses or undergoes Iuteolysis. This then allows for another estrous cycle to begin.
Certainly successful breeding depends on the ability to detect candidate animals in estrus. In situations where cattle are bred naturally, herd bulls handle these demands very well. However, in dairy operations, it is not usual to maintain bulls for breeding. Dairy bulls are exceptionally aggressive and dangerous.
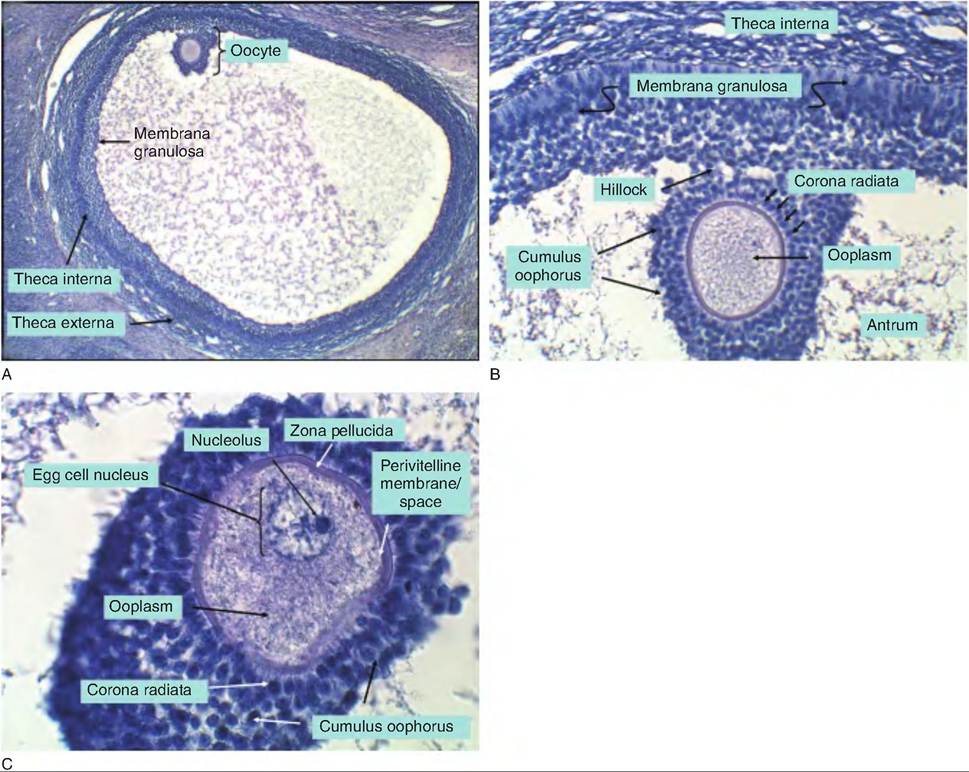
Fig. 19.8. Details of tertiary follicle development. In panel A, the antrum occupies most of the area. The ovum (without evident nucleus) appears to the upper left. The granulosa membrana (the edge of the layer of follicular cells) borders the antrum. To the outside, theca interna and theca externa are evident as distinct layers. In panel B, regions of the cumulus oophorus and corona radiata (layer of granulosa cells that immediately surround the ovum) and hillock (group of granulosa that anchor the oocyte within the antral space) are illustrated. In panel C, some of the detail of the ooplasm can be seen as well as the oocyte nucleus and the zona pellucida and perivitelline space.
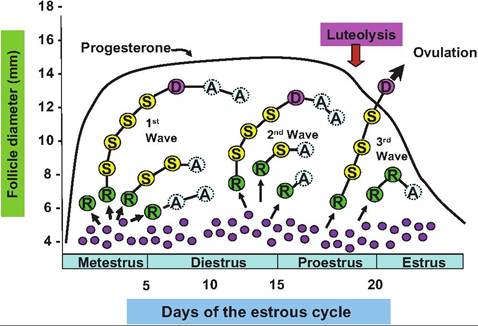
Fig. 19.9. Several follicular waves occur during the estrous cycle. The small filled circles represent gonadotropin-sensitive follicles. During each wave, some follicles are recruited (R), some of these are selected (S), and some become dominant (D). Most eventually regress or become atretic (A). Only follicles recruited during the third wave or after Iuteolysis of the corpus Iuteum produced in the previous cycle will become eligible for ovulation.
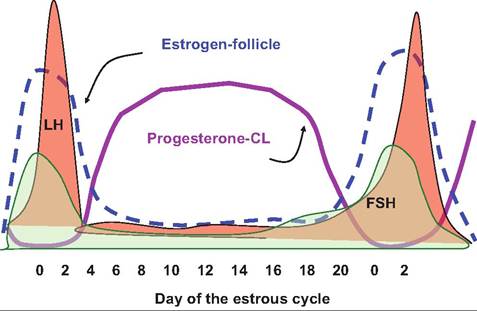
Fig. 19.10. Relative changes in secretion of major reproductive hormones during a bovine estrous cycle. During the follicular phase (proestrus+estrus) from about day 1 7 to 1, progesterone declines rapidly due to Iuteolysis, and there is a correspondingly rapid increase in estrogen coming from recruited and selected follicles. This promotes increases in FSH with further stimulation of estrogen secretion and finally the dramatic LH surge that leads to ovulation.
Thus, the physical needs for adequate handling and housing facilities can be substantial. Perhaps more importantly, genetic progress in dairy has depended on the ability to breed many cows to genetically superior bulls. This is only possible by use of Al. Without bulls to detect females in heat, this means that producers had to learn other methods of heat detection. Fortunately, behavioral cues in cattle are dramatic and animals are housed so that interactions between females are possible, that is, appropriate pastures or lots with good footing and sufficient time for observation. Specifically, as the cow enters estrus she gradually begins to display activity that signals approaching sexual receptivity. These include increased physical activity (locomotion), bellowing, nervousness, and attempts to mount other females. As the period progresses, the female's willingness to accept the male increases. During this time the cow will display mating posture called lordosis. Such animals stand still so that that herd mates will periodically engage in mounting behavior. The animal ready for breeding is said to be in standing heat. Observation of standing heat is the major cue for managers to breed these animals. To be most efficient—after breeding, spermatozoa deposited in the body of the uterus after AI have a limited viability period—it is important to have the inseminate deposited very near the time of ovulation. This means that most operations would need to observe candidate animals at least twice daily, likely 30-45 minutes per session. Since labor is an important part of farm costs, it is not surprising that multiple techniques to better automate heat detection have been devised. As examples, patches placed on the rump of candidate animals that contain packets of dye that rupture when the animal is mounted, mounted pressure sensors that send radio signals to a base station on the farm, or monitoring of activity via daily reading of pedometers are all methods used to decrease the costs and increase the efficiency of heat detection. One of the more interesting experimental approaches was to train dogs to identify cows in heat.
In recent years, control of estrous cycles to more efficiently manage the breeding of cattle has become a reality through the use of estrus synchronization schemes. This has become possible because of the availability of commercial formulations of PGF2OC, GNRH, and progesterone. Indeed there are now effective schemes to allow the timed breeding of cows and heifers without the need for heat detection. As an example, Peeler et al., 2004 reported results from a trial in which heifers were synchronized using intravaginal progesterone inserts also called controlled internal drug release devices (CIRD), coupled with GNRH treatment and timed Al. Briefly, heifers were assigned to the protocol irrespective of stage of the estrous cycle. At this time they received a CIDR containing 1.38 g of progesterone and a 1-mg injection of estradiol cypionate (ECP). After 7 days, the CIDR was removed, and the heifers were given a 25-mg injection of PGF2OC. On day 9, the heifers were given an injection containing 100-μg GNRH. The heifers were bred 48, 56, or 72 hours after the CIDR was removed. The first service pregnancy rates averaged 57.6%. There was no estrus detection in the study but more importantly the success rate was similar to that that obtained with laborious heat detection systems. Some of the advances in reproductive technologies have recently been reviewed (Sakaguchi, 2011; Wiltbank et al., 2011; Perry, 2012).