Fertilization and pregnancy
Assuming a successful ovulation and insemination has occurred, the reproductive story is just beginning. As spermatozoa ascend through the cervix, uterine body, and the oviduct, many are lost.
However, those that remain must undergo a process called capacita- tion. These are biochemical changes to the sperm cells induced by secretions of the female reproductive tract. It is generally accepted that exposure of spermatozoa to seminal fluid during maturation in the testes or at the time of ejaculation leads to the coating of the cell surface with a complex layer of proteins and carbohydrates. Removal of these materials via the Capacitation process in the female tract is essential for the spermatozoa to bind the oocyte. Fertilization typically occurs when the oocyte and spermatozoa meet in the ampulla region of the oviduct.Interestingly, when the spermatozoa reach this area, swimming patterns change from very regular linear movements to more a more erratic or frenzied motion. This change is induced by molecules secreted in this region of the oviduct and is thought to increase the opportunity of contact between sperm and oocyte. Fertilization depends on a complex series of steps, which are outlined in Figure 19.11.
Sperm cells have very specific proteins associated with the acrosome portion of the head of the spermatozoa, which have an affinity for the zona pellucida of the oocyte. This outer layer of the oocyte is composed of three glycoproteins called zona protein 1, 2, and 3 or ZPl, ZP2, and ZP3 for short. ZPl and ZP2 are primarily structural proteins that maintain the space and organization of the zona pellucida. ZP3, in contrast, acts an anchor or receptor for proteins found in the membrane of the sperm cell. Two binding sites are present. The primary zona binding region allows for the close adherence of the oocyte and sperm cell.
A second site induces the acrosome reaction when ZP3 from the zona pellucida binds.The acrosome is a lysosome-like, membrane-bound structure oriented around the outer portion of the
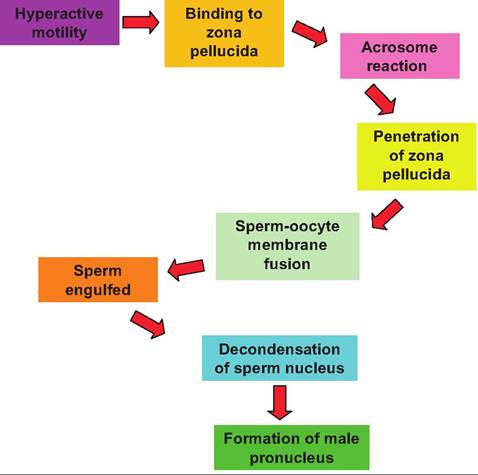
Fig. 19.11. Events following Capacitation and fertilization.
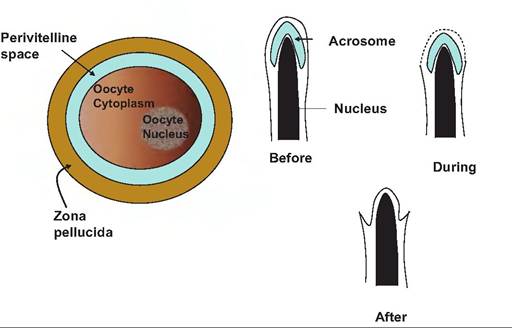
Fig. 19.12. The oocyte and fertilization. The upper panel illustrates the structures of the oocyte. Before fertilization can take place, the spermatozoon must penetrate the zona pellucida, enter the perivitelline space, and ultimately fuse with the oocyte plasma membrane and be engulfed. The lower panel illustrates events of the acrosomal reaction. During the reaction, the outer plasma membrane surrounding the sperm head fuses with the other membrane of the acrosome. This leads to release of the acrosomal contents. The end result is that the inner membrane of the acrosome encompasses the nucleus and continues in a continuous fashion with the remaining plasma membrane of the spermatozoon
spermatozoa where it partially encapsulates the condensed nucleus of the spermatozoa. It contains enzymes that are important for events associated with fertilization. The acrosome reaction is essential because it allows the spermatozoa to penetrate the zona pellucida, and it exposes the nucleus of the sperm cell that is now surrounded by the former inner membrane of the acrosome. After binding, the acrosomal reaction begins as the plasma membrane surrounding the sperm head forms fusion points with the acrosomal membrane. As small vesicles appear, this produces a morphological cellular landmark called Vesiculation that is used to identify cells that have undergone the acrosome reaction. Among the enzymes released, proacrosin has a very high affinity for the zona pellucida. This supports the close adherence of the oocyte and spermatozoa undergoing the acrosome reaction.
When activated to acrosin, its hydrolytic action degrades the zona pellucida in the very local region where the sperm cell is attached. This is important because the continued presence of the zona pellucida is necessary for subsequent development. The continued beating of the tail of the spermatozoa supplies the mechanical force needed for the activated spermatozoa to penetrate through the zona pellucida to the perivitelline space, which surrounds the plasma membrane of the oocyte. When the spermatozoon gets to the perivitelline space it binds to microvilli on the surface of the oocyte. The plasma membrane of the oocyte then fuses with the membrane surrounding the spermatozoon and engulfment occurs. After this event, dense vesicles—the cortical granules, previously produced inside the oocyte—migrate to the cell surface and release their contents by exocytosis. The proteases, mucopolysaccharides, plasminogen activator, acid phosphatases, and peroxidases contained in the granules alter the biochemical properties of the zona pellucida so that additional sperm cannot enter. This is called zona block. In some species these substances also reduce the capacity of the oocyte plasma membrane to fuse with additional spermatozoa, the vitelline block. This provides a second mechanism to prevent polyspermy (fertilization by more than one spermatozoon), which results in embryonic death. After the sperm nucleus is freed into the cytoplasm of the oocyte, it becomes the male pronucleus. However, this requires a major decondensation of the sperm nucleus. During the maturation of the spermatozoa in the testis the nucleus becomes very highly compacted and ordered due to the creation of many disulfide crosslinks. In this state the chromosomes are essentially inert. Fortunately, the oocyte cytoplasm is rich in glutathione, which allows for the loosening of the chromosomes so that interaction can take place. The last step in fertilization is the fusion of male and female pronuclei. This is called syngamy. After this point the zygote enters the first stages of embryo genesis. Structures of the oocyte and spermatozoa are illustrated in Figure 19.12 (see also follicular development).Implantation and placentation
After a successful fertilization, the embryo then develops into a blastocyst and hatches from the surrounding
zona pellucida. It then develops a functional trophoblast and secretes signaling proteins that allow maintenance of the corpus luteum. After fusion of the male and female pronuclei, the single-celled egg is called a zygote. It can also be referred to as an embryo (defined as an organism in the early stages of development). Embryos typically have not acquired features that allow for recognition of a particular species. By contrast a fetus—a potential offspring still within the uterus—can generally be recognized as a member of the species. A conceptus, defined as the product of conception, is a commonly used term but is poorly descriptive. It is really a catch-all term since it includes early and late embryos, the embryo + extraembyronic membranes during preimplantation, as well as the fetus and placenta.
Soon after syngamy, the zygote initiates as series of mitotic divisions or cleavages. The first cleavage produces a two-celled embryo. Each of the cells at this time is called a blastomere. Subsequent divisions produce 4, 8, and 16 identical daughter cells. At these early stages the blastomeres are totipotent. In other words, each of the individual cells is capable of giving rise to a fully formed offspring. For example, identical twins can be produced in experimental situations by separating individual blastomeres, placing then into surrogate zona pellucida, and allowing development in the uterus of a recipient female. It was long thought that adult or differentiated cells were not capable of this process. However, the production of Dolly the sheep from adult mammary gland fibroblast cells showed that it is possible with just the correct conditions to dedifferentiate cells so that they can become totipotent once again.
Once beyond the 16-cell stage it becomes impossible to accurately count the growing ball of cells so the structure is called a morula. With further divisions a blastocyst develops. It is characterized by the presence of inner cell mass (ICM), a fluid- filled cavity called a blastocoele, and a layer of cells around the periphery called the trophoblast. Figure 19.13 illustrates some of these stages of development.During the morula stage the cells begin to segregate into two distinct populations, inner and outer cells. Cells in the ICM develop gap junctions which allows for coordinated communication between the cells. The outer cells, by contrast, become linked by tight junctions. This allows for the creation of a unique environment with respect to ions inside the developing blastocoele. Specifically, increasing concentrations of Na+ produce osmotic drive for water to enter the structure and expand the volume of the blastocoele. The ICM develops into the embryo while the trophoblast gives rise to the chorion, which eventually becomes the fetal component of the placenta. With continuing expansion of the blastocyst via fluid and cell proliferation and pressure increases, the trophoblast cells begin to secrete enzymes and an area of the zona pellucida degrades and ruptures. The blastocyst escapes or hatches and becomes a free-floating embryo within the lumen of the uterus. This development cycle is illustrated in Figure 19.14.
After hatching of the blastocyst, cell division occurs very rapidly. For example, in the cow on day 13 the blastocyst is about 3mm in diameter. Over the next few days the blastocyst increases in length to 250 mm and appears as a string or thread. By day 18 of gestation, the blastocyst occupies areas of both uterine horns. Much of this growth involves appearance of extraembryonic membranes essential for the embryo to attach to the uterus of the dam for subsequent development. The outer trophoblast combined with the newly developed endoderm gives rise to the chorion and amnion.
The yolk sac develops from the endoderm (see Fig. 19.15).Implantation is the attachment of the free-floating blastocyst to the uterine epithelium and corresponding growth or penetration of the epithelium by embryonic tissues. Just how dramatic and extensive this interaction is varies among species. In most farm animals, the degree of penetration is much less than in rodents and humans, for example. Implantation in most farm animals is considered noninvasive and primarily associated with cell to cell junctions between embryonic and uterine tissues. After fertilization, attachment occurs in the cow at about 35 days, the sow about 11 days, and the mare about 55 days.
Placentation refers to the development of the extra- embryonic membranes or the placenta. The placenta and its layers allow for exchanges between the maternal and fetal circulations so that nutrients can be supplied and wastes removed. The chorion is the outer most membrane and therefore is in contact with the maternal uterine wall. The next layer toward the fetus is the allantois, which forms a continuous layer that creates a fluid-filled sac—the allantoic cavity—around the fetus. The amnion is the closest membrane to the fetus. It also forms a fluid-filled cavity in direct contact with the fetus. The amnion is fused to the inner layer of the allantois. When parturition occurs, the allantoic sac is expelled, followed by the amniotic sac. Branches from the umbilical arteries and veins pass through the connective tissue between the allantois and chorion. The physical attachment of the conceptus to the uterus offers major reproductive advantages. Modification of the intrauterine environment as the fetus develops ensures that the developing fetus will be provided with nutrients and physiological support to maximize the chance for a successful birth. This was a major evolutionary step for eutherian mammals compared with lower animals that laid their eggs.
Connections between the chorion and the uterus are varied between species. By convention, the histological
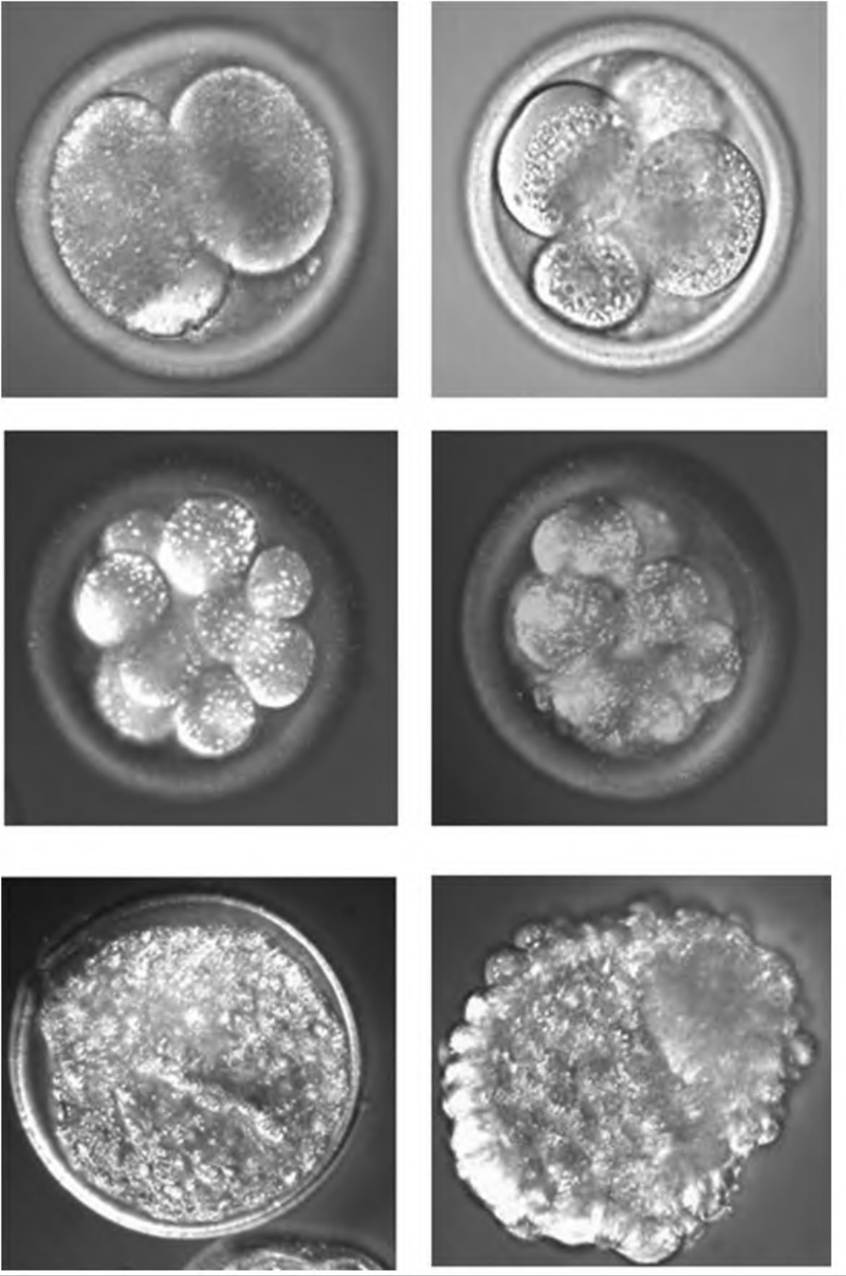
Fig. 19.13. Stages of bovine oocyte development. The upper panels (left to right) are 2 and 4 cells, the middle panels are 8 cells and morula, and the lower panels are expanded blastocyst and hatched blastocyst, respectively.
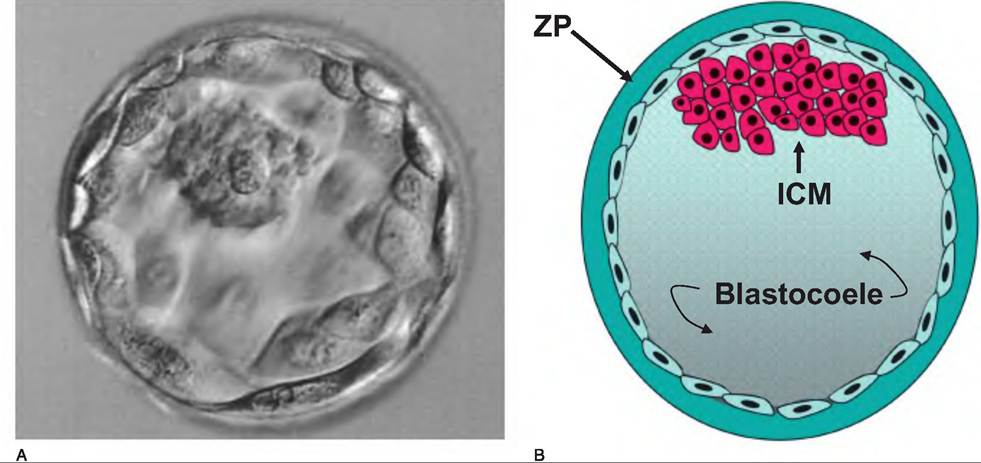
Fig. 19.14. Photograph and representation of a preimplantation embryo just before hatching. The inner cell mass (ICM) develops into the embryo (panel A), and the trophoblast cells (lower arrows) that surround the expanding space or blastocoele give rise to the chorion (panel B). After expansion, hatching occurs, so that the blastocyst escapes to become a free-floating embryo within the uterus prior to implantation.
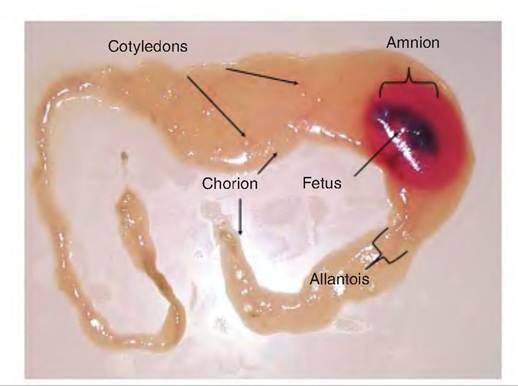
Fig. 19.15. Developing bovine fetus and associated membranes.
relationship between the fetal membranes and uterine epithelium is named beginning with the maternal side. For example, placenta of most farm animals is classified as epitheliochorial. This means that the chorion of the fetus is in direct contact with the uterine surface. This is the least intimate arrangement of the types of placentas. In hemochorial placentation, fetal vessels and chorion are invaginated so that there is direct contact between a single layer of chorion epithelial cells and maternal blood. In hemoendothelial placentation, maternal blood can directly bathe the outer surface of chorionic capillaries. The number of layers separating fetal and maternal circulations is important in part because this affects transfer of immunoglobulins in utero. Since most farm animals have epitheliochorial connection, calves, for example, are born without protective antibodies from the dam. Consequently, absorption of antibodies from the colostrum is very important. Primates, rabbits, rodents, and other species with the hemochorial or hemoendothelial arrangements are born with some antibodies that transfer in utero from the mother.
Placentation patterns also vary grossly. In the horse and pig, extensions from the chorion (chorionic villi) project into crypts scattered over the entire surface of the endometrium. This is called a diffuse placenta. Ruminants have scattered attachments, which depend on a cotyledonary type of connection. In this situation, exchange takes place at distinct structures called plac- entomes. The placentomes are formed by the combined invagination of specific regions of the chorionic membrane, the cotyledons, into the button-like projections from the surface of the endometrium, the caruncles. Caruncles project from the surface of the uterus about 2cm and vary from 1 to IOcm in diameter. The caruncles also increase in size as pregnancy advances and are larger in the gravid horn than in the nongravid horn of the uterus. Zonary placentas, found in dogs and cats, for example, are characterized by a major region of exchange that forms an area near the center of the developing conceptus essentially in a band. In primates and rodents, the discoid placenta type occurs. It is characterized by the presence of one of two distinct discs on a region of the chorion. The discs house the chorionic villi, which interconnect with the endometrium.
Exchanges that occur across the placenta involve typical mechanisms, that is, diffusion, facilitated diffusion, and active transport. Glucose is the major energy source for the fetus. Since lipids do not cross readily cross the placenta, triglycerides are hydrolyzed and the fatty acids reesterified. With the exception of antibodies—depending on the placentation type— large proteins do not pass the placenta. Unfortunately, many toxic substances are readily absorbed. Appearance of fetal alcohol syndrome in humans is evidence of the ability of ethanol to pass the placenta and adversely affect fetal development. Substances that cause abnormal fetal development or birth defects are teratogenic.
The placenta is also a major endocrine organ during pregnancy. For example, mares produce a gonadotropin—equine chorionic gonadotropins (ECG), sometimes called pregnant mare serum gonadotropin (PMSG). The hormone acts like LH and serves to stimulate the maintenance of the primary CL on the ovary. Concentrations are especially high during the period between 40 and 100 days of gestation. In fact, this "extra" stimulus can promote additional luteinization of ovarian follicles to ensure that an adequate level of progesterone is maintained. When used in other species, PMSG has a very strong FSH-Iike action, so that it is useful to induce follicular development, that is, in superovulation and embryo transfer (ET) schemes.
In addition to the ovaries, in many species the placenta is a major source of estrogen and progesterone. For example, in the sow or rabbit, Iutectomy (surgical removal of the CL) terminates the pregnancy at any time during gestation. But in other species, the placenta can take over the production of progesterone. The time when the placenta can take over and maintain the pregnancy in the cow, ewe, mare, and human is 7 months, 50 days, 70 days, and 50 days, respectively.
The placenta also produces a polypeptide in many species that has properties similar to Prl and GH; it is now most commonly referred to as placental lactogen. Whether the GH- or Prl-Iike activity predominates varies markedly, as does the amount of placental lactogen that appears in the material circulation. In sheep and goats, concentrations of several hundred nanograms per milliliters are common near the time of parturition but concentrations are only 1-2ng∕mL in cattle.