Fibrinogen
Fibrinogen is a positive acute phase protein. Most fibrinogen is utilized at the site of inflammation. The fibrin that results from cleavage of fibrinogen serves to wall off bacterial agents and form a scaffold for the healing process to begin.
Fibrinogen increases erythrocyte aggregation, leading to an increased sedimentation rate, which can also be used as an indicator of inflammation (Kesmarky et al., 2008).Fibronectins
Fibronectins are important positive acute phase reactants. Tissue fibronectins bind fibrocytes and epithelial cells to the extracellular matrix, maintain cell shape, and promote fibroblast growth. Insoluble fibronectins bridge fibrin to cells and promote platelet adhesion (Stoffels et al., 2013). Soluble fibronectins opsonize bacteria to decrease bacterial adherence to tissues and stimulate phagocytosis by inflammatory leukocytes (Butler et al., 1987).
Transferrin and hepsidin
Transferrin and hepsidin are acute phase proteins involved in iron transport. Iron is an essential nutrient for most pathogenic bacteria, so protective mechanisms have developed to sequester iron away from bacteria (Johnson & Wessling-Resnick, 2012). During inflammation, transferrin is downregulated and neutrophils release lactoferrin, which removes iron from iron–transferrin complexes in tissues and produces oxidizing agents that can kill bacteria. Macrophages then take up iron–lactoferrin complexes and sequester iron within ferritin. Hepcidin is a positive acute phase protein that binds to the iron uptake protein of the intestine to prevent iron uptake from the gastrointestinal (GI) tract.
Complement
Complement components are small plasma glycoproteins, mostly synthesized in the liver, that become activated by a series of enzymatic cleavages (Figure 2.4). There are three pathways to complement formation: (1) the mannose-binding lectin pathway, (2) the alternative pathway, and (3) the classical pathway.
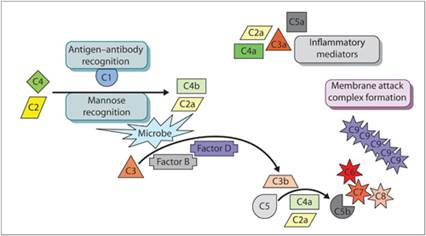
Figure 2.4 Complement cascade. Inactive complement proteins are cleaved by activated complement proteins to form inflammatory mediators and the membrane attack complex.
The mannose-binding lectin pathway is activated by unique sugars on the surfaces of bacterial and fungal pathogens that are not present on vertebrate cells. Mannose-binding lectins recognize foreign sugars and activate complement components C2 and C4. This leads to the activation of C3 and C5 and the formation of a membrane attack complex (MAC), made up of a complex of C5b, C6, C7, C8, and C9 molecules, which induces cell lysis.
The alternative pathway is microorganism dependent. This pathway protects the host during the early phases of microbial invasion when sufficient antibodies have not yet been produced. The alternative pathway is nonspecific and is activated when C3b binds to a pathogen, preventing plasma proteins from inactivating C3b. Factor B in the plasma binds to C3b. Factor D then cleaves factor B. The cleaved segment of factor B combined with C3b activates additional C3 molecules.
The classical pathway is immune complex dependent. Antibody–antigen (Ab–Ag) complexes bound to an infectious organism activate C1. Activation of C1 leads to the sequential activation of other C proteins in the cascade.
Complement fragments C3a, C4a, and C5a are anaphylatoxins; they mediate histamine release from mast cells, leading to increased vascular permeability. Fragment C5a also is a chemoattractant for neutrophils and increases neutrophil respiratory burst. Complement component C3b opsonizes bacteria to make them more prone to phagocytosis by neutrophils and macrophages. Other C3 fragments can mobilize cells from the bone marrow. Neutrophils and macrophages have receptors for C fragments and are resistant to MAC lysis (Sacks, 2010). Lymphocyte functions are also modified by complement components (Clark & Tenner, 2014).
Cell-derived inflammatory mediators
Vasoactive amines
Vasoactive amines (histamine and serotonin) are stored in mast cells, platelets, and basophils and are secreted extremely early following tissue injury. They initiate the first phase of vascular permeability in the inflammatory response. Vasoactive amines cause dilation of pre-capillary sphincters of arterioles and induce venule and capillary endothelium to round up and develop intercellular gaps that allow fluid to escape (Figure 2.5).
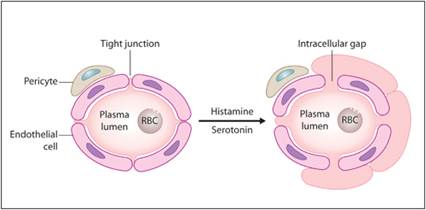
Figure 2.5 Effect of vasoactive amines on capillaries. Histamine and serotonin induce endothelial cells to round up and develop intercellular gaps that allow fluid to escape.
Histamine and serotonin
Histamine induces vasodilation and causes endothelial cells to express adhesion molecules for neutrophils and macrophages. The effects of histamine are short-lived (15–20 minutes) but immensely important for the initiation of inflammation. Serotonin has many of the same physiologic effects as histamine and acts very early during the inflammatory response to mediate smooth muscle contraction and pain. These vasoactive substances are rapidly inactivated (Rutkowski et al., 2012).
Eicosanoids
Eicosanoids are derivatives of arachidonic acid (AA), which is one of the fatty acids released from cell membranes during cell membrane metabolism, injury, or death. Membrane phospholipids are broken down by phospholipases to form free AA and lysophosphatidylcholine (Figure 2.6). AA-derived eicosanoids include prostaglandins, thromboxanes, leukotrienes, lipoxins, and hydroxyeicosatetraenoic acids. The type of eicosanoid produced is dependent on which enzymes mediate the AA breakdown. The enzymes present in a cell are dependent on the cell type.
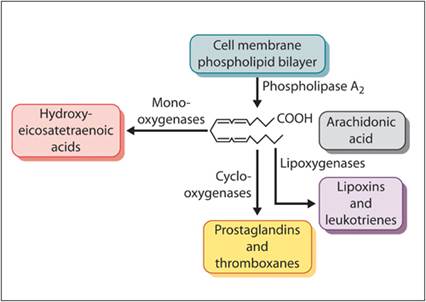
Figure 2.6 Formation of eicosanoids.
Membrane phospholipids are digested by phospholipase A2 to form arachidonic acid and lysophosphatidylcholine. Mono-oxygenases in the cell process arachidonic acid into hydroxyleicosatetraenoic acids, whereas cyclo-oxygenases produce prostaglandins and thromboxanes and lipoxygenases stimulate the formation of lipoxins and leukotrienes.
Prostaglandins and thromboxanes
Prostaglandins are produced by the action of cyclo-oxygenase (COX). Prostaglandin Es (PGEs) are key mediators of inflammation and are produced by macrophages, platelets, and several other cell types. Most PGEs affect vasodilation, increase blood flow, and mediate pain. They also induce the release of neutrophils into the circulation from the bone marrow and peripheral storage pools. Thromboxanes are also produced by COX, mainly in platelets. Thromboxane A2 acts as a vasoconstrictor and facilitates platelet aggregation.
Leukotrienes
Leukotrienes (LTs) induce leukocytes to move out of blood vessels towards an inflammatory stimulus and are extremely potent mediators of vascular responses. Some forms of LTs are more than 1,000? more potent than histamine (Dahlen et al., 1981). Lipoxygenase initiates the LT cascade and is activated when cytosolic Ca2+ levels are elevated. Endotoxin from Gram-negative bacteria can be a potent stimulator of LT production (Rossi et al., 2005). Neutrophils produce a large amount of the precursor form LTA4 while other cells help to convert LTA4 to more active forms of LTs. Myeloid cells (neutrophils, eosinophils, and basophils) can produce both LTBs as well as LTC4, LTD4, and LTE4. Nonmyeloid cells (platelets, endothelial cells, Kupffer cells, and hepatocytes) can only synthesize LTC4, LTD4, and LTE4.
Leukotriene B4 is chemotactic for neutrophils and makes them more adherent to endothelium (Sadik & Luster, 2012). The respiratory burst and release of enzymes by neutrophils are enhanced by LTB4. Also, immunoglobulin production by B lymphocytes is indirectly stimulated by LTB4 (Terawaki et al., 2005). Leukotriene B4, LTC4, LTD4, and LTE4 synergize to increase capillary permeability. In addition, LTC4, LTD4, and LTE4 induce airway smooth muscle contraction in asthma and anaphylaxis in certain species.